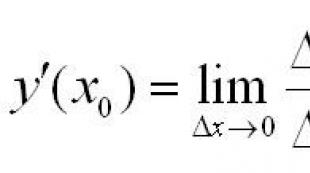Millised organellid täidavad algloomadel hingamisfunktsiooni. Lihtsamad üherakulised organismid. Millised organellid moodustavad raku?
ALLKUNINGRIIGI PROTOZOOTID,
VÕI ÜKSRAKK (ALGLOOS)
Üherakuliste organismide alamriiki kuuluvad loomad, kelle keha koosneb ühest rakust. Morfoloogiliselt on nad sarnased paljurakuliste loomade rakkudega, kuid füsioloogiliselt erinevad selle poolest, et lisaks raku tavapärastele funktsioonidele (ainevahetus, valgusüntees jne) täidavad nad terve organismi funktsioone (toitumine, liikumine, paljunemine, kaitse ebasoodsate keskkonnatingimuste eest). Üksikud funktsioonid mitmerakulistes organismides täidavad neid spetsiaalsed elundid, koed või rakud ning ainuraksetes organismides täidavad keha funktsioone konstruktsioonielemendidüks rakk - organellid. Rakkude jagunemine mitmerakulistel loomadel põhjustab keha kasvu ja algloomadel paljunemist.
Seega on algloomad üherakulise organisatsioonitasandi organismid. Algloomade organismi terviklikkust säilitavad ühe raku funktsioonid ja mitmerakulistes organismides - rakkude, kudede ja elundite koostoime tõttu.
Algloomade elutsükkel koosneb üherakulise organisatsiooniga arengufaasidest ja paljurakulistes organismides vahelduvad üherakulised arengufaasid paljurakulistega.
Praegu on teada enam kui 39 tuhat algloomaliiki, kuid igal aastal avastatakse kümneid ja sadu uusi liike, mis näitab selle loomarühma ebapiisavat tundmist.
Algloomad avastas esmakordselt hollandi teadlane A. van Leeuwenhoek, mikroskoobi esimene leiutaja (1675). Tema mikroskoobid olid suure suurendusega luubid, mis võimaldasid suurenduse 100 ja isegi 200 korda. Esimesed mikroskoobid avastasid eriti palju algloomi ürtide infusioonides (infusum - tähendab "tinktuura"), nii et alguses nimetati neid loomi "tinktuurateks" või ripslasteks. Nüüd on seda nime säilitanud ainult üks algloomade rühm. C. Linnaeuse (1759) esimeses loomade süsteemis määrati algloomad ühte perekonda – Chaos – usside klassist. Ainult
19. sajandil Kölliker ja Siebold määratlesid need iseseisva tüübina (1845). 1977. aastal toimunud rahvusvahelisel algloomade kongressil võeti vastu uus algloomade süsteem, mis kajastab teaduse uusimaid saavutusi. 1980. aastal avaldatud uute põhimõtete kohaselt (Levine et al.) rühmitatakse algloomad alamkuningriiki Üksrakulised ja jagatakse seitsmeks hõimkonnaks.
Algloomade kehakuju on äärmiselt mitmekesine. Nende hulgas on muutuva kehakujuga liike, nagu amööbid. Algloomadel on erinevat tüüpi sümmeetriat. Levinud on radiaalse sümmeetriaga vormid: radiolariaanid, päikesekalad. Need on peamiselt ujuvad planktoni algloomad. Mõnel on täheldatud kahepoolset sümmeetriat
flagellaadid, foraminifera, radiolarians. Translatsiooni-rotatsiooni sümmeetria on iseloomulik spiraalselt keerdunud kestaga foraminiferadele. Mõnel liigil täheldatakse metamerismi - struktuuride korratavust piki pikitelge. Algloomade ehk morfoadaptiivsete tüüpide eluvormid on mitmekesised. Kõige tavalisemad vormid on: amööboid, mis juhivad roomavat elustiili erinevatel aluspindadel vees või sees vedel keskkond peremehe kehas; merekarp- istuvad põhjaelustikud; aktiivselt ujuvad lipikud ja ripslased planktonis hõljuv radiaalne või kiirgav, vormid; istuv - jälitanud, kitsa või lameda korpusega aluspinna puuraugud - vahereklaamid, samuti ümmargused, statsionaarsed, puhkamas vormid (tsüstid, eosed).
Algloomaraku struktuuri iseloomustavad kõik peamised tunnused rakuline struktuur eukarüootid. Algloomade ehituse ultrastruktuuri on bioloogid uurinud elektronmikroskoopia tehnoloogia abil. Kaasaegse elektronmikroskoobi lahutusvõimed võimaldavad saada 200-300 tuhandekordset suurendust.
Algloomrakk on tüüpiline eukarüootsetele organismidele ja koosneb tsütoplasmast ja ühest või mitmest tuumast. Tsütoplasma on väliselt piiratud kolmekihilise membraaniga. Membraani kogupaksus on umbes 7,5 nanomikronit (1 nm = 10 - 6 mm). Algloomade tsütoplasmas on välimine, läbipaistvam ja tihedam kiht - ektoplasma ja sisemine, granuleeritud kiht - endoplasm. Endoplasmasse on koondunud kõik raku peamised organellid: tuum, mitokondrid, ribosoomid, lüsosoomid, endoplasmaatiline retikulum, Golgi aparaat jne Lisaks on algloomadel spetsiaalsed organellid: toetavad, kokkutõmbuvad fibrillid, seede- ja kontraktiilsed vakuoolid jne. Tuum on kaetud kahekihilise membraaniga, millel on kohati. Tuuma sees on karüoplasma, milles on jaotunud kromatiin ja nukleoolid. Kromatiin on despiraliseeritud kromosoomid, mis koosnevad DNA-st ja valkudest, näiteks histoonidest. Nukleoolid on sarnased ribosoomidega ja koosnevad RNA-st ja valkudest. Algloomade tuumad on erineva koostise, kuju ja suuruse poolest.
Algloomadel saab eristada spetsiaalseid organellide funktsionaalseid komplekse, mis vastavad mitmerakuliste organismide elundite ja kudede süsteemidele.
Struktuursed ja toetavad organellid. Mõnel üherakulisel liigil ei ole sise- ega tugistruktuure. Selliste algloomade rakku piirab ainult pehme tsütoplasmaatiline membraan. Sellistel liikidel puudub püsiv kehakuju (amööb). Teistel liikidel on tihe elastne kest - pelliikul, mis on moodustunud ektoplasma perifeerse kihi tihendamise ja erinevate ainete olemasolu tõttu.
toetavad fibrillid. Sel juhul on algloomadel kindel kehakuju (ripsloomad, euglena) ja samal ajal säilitavad nad painduvuse ning võivad liikumisel painduda ja osaliselt kokku tõmbuda. Teised üherakulised organismid eritavad väljapoole soomuste kesta, mis takistab kehakuju (diatomite) muutumist. Kehakuju võivad lisaks toetada ka muud tugistruktuurid – fibrillid, mis moodustavad osadel ripsmetel näiteks ajukoore.
Tugikonstruktsioonide hulka kuulub ka skelett. Algloomade skelett võib olla välimine (kest) või sisemine (skeletikapslid, nõelad). Kest eritub raku ektoplasma poolt ja samal ajal moodustub rakuväline moodustis, millel on kaitsefunktsioon. Sisemine skelett moodustub raku endoplasmas. Skeleti kapslite ja nõelte moodustumine toimub biokristallimise teel. Skeleti moodustised koosnevad orgaanilistest ja mineraalsetest ainetest. Kõige sagedamini sisaldavad algloomade skeletid kaltsiumkarbonaati (CaCO 3) või ränioksiidi (SiO 2), harvemini strontsiumsulfaati (SrSO 4).
Motoorsed organellid. Algloomade kõige primitiivsemaks liikumisviisiks võib pidada amööboidset liikumist valejalgade ehk pseudopodia abil. Sel juhul moodustuvad raku spetsiaalsed eendid, millesse tsütoplasma voolab. Sellised liikumisorganellid on iseloomulikud muutuva kehakujuga üherakulistele organismidele.
Rohkem keeruline liikumine iseloomulik algloomadele, mille liikumisorganellideks on lipud või ripsmed. Lipu ja ripsmete struktuur on sarnane (joon. 16). Iga flagellum on väliselt kaetud kolmekihilise tsütoplasmaatilise membraaniga. Lipu sees on fibrillid: kaks keskmist ja üheksa topeltperifeerset. Lipu kinnitatakse tsütoplasma külge, kasutades basaalkeha – kinetosoomi. Tavaliselt tekitavad flagellad pöörlevat liikumist ja ripsmed sõudmisliikumist. Lipukesed on iseloomulikud ripsloomadele ja ripsmed on iseloomulikud ripsmetele.
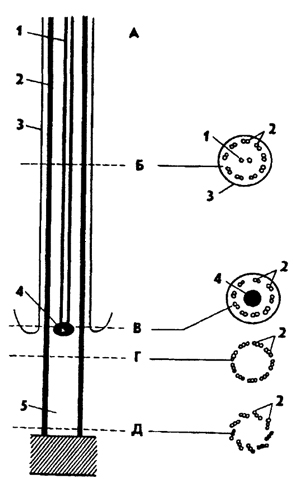
Riis. 16. Lipu ehituse skeem (Noirot-Timote järgi): A - lipu pikisuunaline läbilõige, B, C, D, E - lipu ristlõiked erinevatel tasanditel; 1 - tsentraalsed fibrillid, 2 - perifeersed fibrillid. 3 - välimine membraan flagellum, 4 - aksiaalne graanul, 5 - kinetosoom
Mõned algloomad on võimelised keha kiiresti kokku tõmbuma spetsiaalsete kontraktiilsete fibrillide – müoneemide – tõttu. Näiteks istuvad ripsloomad - suvoiki - on võimelised oma pikka varre järsult lühendama ja spiraaliks voltima. Radiolaariaanid on võimelised kas venitama rakukeha radiaalsetel selgrool või kokkutõmbuma kontraktiilsete kiudude abil. See tagab neile veesambas tasuta ujumise reguleerimise. Ebasoodsates tingimustes paljud algloomad tsüsteeruvad, s.t. eritavad enda ümber tihedat membraani ja muutuvad tsüstiks.
Toitumise tüübid ja troofilised organellid. Algloomad on oma toitumisviiside poolest erinevad. Nende hulgas on autotroofe, mis on võimelised fotosünteesiks. Need on üherakulised lipulised vetikad. Nende tsütoplasmas on klorofülli terad ehk kromatofoorid.
Enamik algloomadest on heterotroofid, kes toituvad nagu loomad valmis orgaanilistest ainetest. Mõnel neist on holosoiline toitumisviis, tahkete toidutükkide allaneelamine. Teised toituvad saprofüütiliselt, absorbeerides lahustunud orgaanilist ainet. Toiduosakesed neelavad alla amööbid ja ripslased. Nende tsütoplasmas moodustuvad seedevakuoolid, kus seeditakse toitu. Sellist tahke toidu allaneelamist raku poolt nimetatakse fagotsütoos. Toitumise saprofüütilise meetodi korral ei moodustu seedetrakti vakuoole. Siiski on teada, et paljud algloomad võivad vedelikku neelata läbi membraani ajutise invaginatsiooni – spetsiaalse lehtri. Seda vedeliku imendumist nimetatakse pinotsütoos.
Mõned liigid on segatüüpi toitumisega (miksotroofid). Nad on võimelised fotosünteesiks, nagu taimed, ja toituma valmis orgaanilisest ainest, nagu loomad. Nende tsütoplasmas on klorofülli terad, kuid võivad tekkida ka seedevakuoolid. Selliste segatüüpi toitumisega algloomade hulka kuuluvad näiteks euglena, mis toitub valguses nagu taimed ja pimedas nagu loomad.
Tuumaaparaat koosneb ühest või mitmest tuumast. Tuumad reguleerivad algloomade rakkude ainevahetusprotsesse ja tagavad paljunemise. Algloomade tuumad on erineva kuju, arvu, ploidsuse ja funktsioonide poolest. Mõnel mitmetuumalisel algloomal eristatakse kahte tüüpi tuumasid: generatiivseid ja vegetatiivseid. Seda nähtust nimetatakse tuuma dualismiks. Vegetatiivsed tuumad reguleerivad rakus kõiki eluprotsesse ja generatiivsed osalevad seksuaalprotsessis. Tuumadualism on iseloomulik ripsloomadele ja mõnedele foraminifeeridele. Algloomade tuumad võivad mõnel elutsükli etapil olla haploidsed või diploidsed või polüploidsed. Enamik algloomi on mononukleaarsed (monoenergeetilised). Liike, millel on palju tuumasid, nimetatakse polüenergeetilisteks.
Algloomade mittesugulisel paljunemisel jagunevad tuumad mitoosi teel. Algloomade tuumad, mille seksuaalprotsess on teada, läbivad meioosi ehk redutseerumisjagunemist. Erinevalt mitmerakulistest organismidest on ainuraksete organismide meioos mitmekesine. Primitiivsel juhul tekib meioos ühe raku jagunemise ajal, teistel, nagu kõrgematel loomadel, kahe järjestikuse jagunemise tulemusena. Mõnel juhul toimub redutseeriv jagunemine pärast sügoodi moodustumist (sügootne redutseerimine), teistel, nagu mitmerakulistes organismides, sugurakkude moodustumise ajal (gameetiline redutseerimine).
Paljunemise tüübid algloomad on mitmekesised. Neid iseloomustab aseksuaalne ja seksuaalne paljunemine. Mittesuguline paljunemine toimub raku jagamisel kaheks või mitmeks rakuks (agamogaamia) tuuma mitootilise jagunemise käigus. Algloomade seksuaalset paljunemist iseloomustab sugurakkude - sugurakkude (gamogaamia) moodustumine koos nende järgneva ühinemisega (kopulatsiooniga), mis viib sügootide moodustumiseni, millest areneb uus tütarorganism. Mõnedel algloomadel (ripsloomadel) ei toimu seksuaalprotsess – konjugatsioon sugurakkude liitmisel, vaid erinevatest rakkudest pärinevate generatiivsete tuumade liitmisel. Kopulatsiooni käigus võivad ühinevad sugurakud olla sama suuruse ja kujuga (isogaamia) või erinevad (heterogaamia). Sugurakkude vaheliste teravate erinevuste korral, kui üks sugurakkudest on suur, liikumatu, ilma flagelladeta (oogameet) ja teine väikese suurusega, vibudega, nimetatakse sellist kopulatsiooni oogaamiaks. Sel juhul võrdsustatakse makrogameet (oogameet) mitmerakuliste organismide munarakuga ja mikrogameet spermaga.
Algloomade elutsükkel tähistab liigi arengu tsükliliselt korduvat segmenti kahe samanimelise faasi vahel (näiteks sügootist sügootini). Algloomade elutsüklit saab iseloomustada ainult mittesugulise paljunemisega (jagunemisest jagunemiseni) või ainult sugulise paljunemisega (sügoodist sügootini) või
vahelduv seksuaalne ja mittesuguline paljunemine (metagenees). Edaspidi käsitletakse üksikasjalikumalt erinevaid algloomade elutsükleid.
Klassifikatsioon. Kaasaegsete kontseptsioonide kohaselt jagatakse algloomad algloomad seitsmeks tüübiks:
Algloomade tüüpideks jaotamine põhineb nende tuumaaparaadi ehituse põhimõtetel, liikumisorganellidel, mitmetel mikrostruktuuridel, paljunemistüüpidel ja elutsüklitel.
Ripsloomad liiguvad liikumisorganellide – ripsmete või nende derivaatide abil; on tuuma dualism ja polüenergia. Seksuaalprotsess viiakse läbi konjugatsiooni teel.
Labürindid elavad vee-meretaimedel ja on tsütoplasmaatiliste ahelate labürint, mida mööda spindlikujulised rakud liiguvad. Nad paljunevad zoospooride abil koos flagelladega.
Algloomade tüüpide võrdlusomadused on toodud tabelis 1.
Organell on püsiv moodustis rakus, mis täidab kindlaid funktsioone. Neid nimetatakse ka organellideks. Organell on see, mis võimaldab rakul elada. Nii nagu loomad ja inimesed koosnevad elunditest, nii koosneb iga rakk organellidest. Need on mitmekesised ja täidavad kõiki raku eluea tagavaid funktsioone: ainevahetust, säilitamist ja jagunemist.
Mis tüüpi organellid on olemas?
Organell on keeruline struktuur. Mõnel neist võib olla isegi oma DNA ja RNA. Kõik rakud sisaldavad mitokondreid, ribosoome, lüsosoome, rakukeskust, Golgi aparaati (kompleks) ja endoplasmaatilist retikulumit (võrkkest). Taimedel on ka spetsiifilised rakulised organellid: vakuoolid ja plastiidid. Mõned liigitavad organellideks ka mikrotuubulid ja mikrokiud.
Organell on ribosoom, vakuool, rakukeskus ja paljud teised. Vaatame lähemalt organellide ehitust ja funktsioone.
Mitokondrid
Need organellid varustavad rakku energiaga – vastutavad nende eest. Neid leidub taimedes, loomades ja seentes. Nendel rakulistel organellidel on kaks membraani: välimine ja sisemine membraan, mille vahel on membraanidevaheline ruum. Seda, mis on kestade sees, nimetatakse maatriksiks. See sisaldab mitmesuguseid ensüüme - kiirendamiseks vajalikke aineid keemilised reaktsioonid. Sisemembraanil on voldid, mida nimetatakse cristaeks. Nende peal toimub rakuhingamise protsess. Lisaks sisaldab mitokondriaalne maatriks mitokondriaalset DNA-d (mDNA) ja mRNA-d, samuti ribosoome, mis on peaaegu sarnased
Ribosoom
See organell vastutab translatsiooniprotsessi eest, mille käigus valk sünteesitakse üksikutest aminohapetest. Ribosoomi organelli struktuur on lihtsam kui mitokondritel – sellel puuduvad membraanid. See organell koosneb kahest osast (allüksustest) - väikesest ja suurest. Kui ribosoom on inaktiivne, eraldatakse need ja kui see hakkab valku sünteesima, siis nad ühinevad. Mitu ribosoomi võivad kokku tulla ka siis, kui nende poolt sünteesitav polüpeptiidahel on väga pikk. Seda struktuuri nimetatakse "polüribosoomiks".

Lüsosoomid
Seda tüüpi organellide funktsioonid piirduvad raku seedimisega. Lüsosoomidel on üks membraan, mille sees on keemilisi reaktsioone katalüüsivad ensüümid. Mõnikord need organellid mitte ainult ei lagune, vaid seedivad ka terveid organelle. See võib juhtuda raku pikaajalise nälgimise ajal ja võimaldab tal mõnda aega elada. Kuigi kui toitained ikkagi voolama ei hakka, siis rakk sureb.

ja funktsioonid
See organell koosneb kahest osast - tsentrioolidest. Need on silindrikujulised moodustised, mis koosnevad mikrotuubulitest. Rakukeskus on väga oluline organell. See osaleb spindli moodustumise protsessis. Lisaks on see mikrotuubulite organiseerimise keskus.
Golgi aparaat
See on kettakujuliste membraanikottide kompleks, mida nimetatakse tsisternaks. Selle organelli funktsioonide hulka kuulub teatud ainete sorteerimine, säilitamine ja muundamine. Siin sünteesitakse peamiselt süsivesikuid, mis on osa glükokalüksist.

Endoplasmaatilise retikulumi struktuur ja funktsioonid
See on ühe membraaniga ümbritsetud torude ja taskute võrgustik. Endoplasmaatilist retikulumit on kahte tüüpi: sile ja kare. Viimase pinnal asuvad ribosoomid. Sile ja kare võrk täidab erinevaid funktsioone. Esimene vastutab hormoonide sünteesi, süsivesikute säilitamise ja muundamise eest. Lisaks moodustuvad selles vakuoolide alged, taimerakkudele iseloomulikud organellid. Kare endoplasmaatiline retikulum sisaldab oma pinnal ribosoome, mis toodavad aminohapete polüpeptiidahelat. Siis satub ta sisse endoplasmaatiline retikulum, ja siin moodustub valgu teatud sekundaarne, tertsiaarne ja kvaternaarne struktuur (ahel on õigel viisil keerdunud).

Vakuoolid
Need on organellid. Neil on üks membraan. Neisse koguneb rakumahl. Vakuool on vajalik turgori säilitamiseks. Ta osaleb ka osmoosi protsessis. Lisaks leidub neid peamiselt veekogudes elavates üherakulistes organismides ja need toimivad pumpadena, mis pumpavad rakust välja liigse vedeliku.
Plastiidid: sordid, struktuur ja funktsioonid
Need on ka organellid. Neid on kolme tüüpi: leukoplastid, kromoplastid ja kloroplastid. Esimesed on mõeldud varutoitainete, peamiselt tärklise säilitamiseks. Kromoplastid sisaldavad erinevaid pigmente. Tänu neile on taimede kroonlehed mitmevärvilised. Organism vajab seda eelkõige tolmeldavate putukate ligimeelitamiseks.
Kloroplastid on kõige olulisemad plastiidid. Suurim kogus neid leidub taimede lehtedes ja vartes. Nad vastutavad fotosünteesi eest – keemiliste reaktsioonide ahela eest, mille käigus orgaaniline aine keha saab orgaanilist. Nendel organellidel on kaks membraani. Kloroplastide maatriksit nimetatakse "stroomaks". See sisaldab plastiidi DNA-d, RNA-d, ensüüme ja tärklise lisandeid. Kloroplastid sisaldavad tülakoide, mündikujulisi membraanmoodustisi. Nende sees toimub fotosüntees. See sisaldab ka klorofülli, mis toimib keemiliste reaktsioonide katalüsaatorina. Kloroplastide tülakoidid ühendatakse virnadeks - granaks. Organellid sisaldavad ka lamelle, mis ühendavad üksikuid tülakoide ja pakuvad nende vahel sidet.
Liikumise organellid
Need on iseloomulikud peamiselt üherakulistele organismidele. Nende hulka kuuluvad lipud ja ripsmed. Esimesed esinevad euglenas, trüpanosoomides ja klamüdomonastes. Flagella leidub ka loomade spermas. Ripslastel ja teistel üherakulistel organismidel on ripsmed.
Mikrotuubulid
Need tagavad ainete transpordi, samuti raku püsiva kuju. Mõned teadlased ei klassifitseeri mikrotuubuleid organellideks.
Iga elusorganism koosneb rakkudest, millest paljud on liikumisvõimelised. Selles artiklis räägime liikumisorganellidest, nende ehitusest ja funktsioonidest.
Üherakuliste organismide liikumisorganellid
Kaasaegses bioloogias jagunevad rakud prokarüootideks ja eukarüootideks. Esimeste hulka kuuluvad kõige lihtsamate organismide esindajad, mis sisaldavad ühte DNA ahelat ja millel puudub tuum (sinivetikad, viirused).
Eukarüootidel on tuum ja need koosnevad mitmesugustest organellidest, millest üks on liikumisorganellid.
Üherakuliste organismide liikumisorganellide hulka kuuluvad ripsmed, lipud, niidilaadsed moodustised - müofibrillid, pseudopoodid. Nende abiga saab rakk vabalt liikuda.

Riis. 1. Liikumisorganellide sordid.
Liikumisorganelle leidub ka mitmerakulistes organismides. Näiteks inimestel on bronhide epiteel kaetud paljude ripsmetega, mis liiguvad rangelt samas järjekorras. Sel juhul moodustub nn "laine", mis suudab kaitsta hingamisteid tolmu ja võõrosakeste eest. Spermatosoididel (isaskeha spetsialiseeritud rakud, mis on mõeldud paljunemiseks) on ka lippe.
TOP 4 artiklitkes sellega kaasa loevad
Motoorse funktsiooni saab saavutada ka mikrokiudude (müoneemide) kokkutõmbumise tõttu, mis paiknevad tsütoplasmas naha all.
Liikumisorganellide ehitus ja funktsioonid
Liikumisorganellid on membraanist väljakasvud, mille läbimõõt ulatub 0,25 µm. Oma struktuuri poolest on lipukesed palju pikemad kui ripsmed.
Mõne imetaja sperma lipu pikkus võib ulatuda 100 mikronini, ripsmete suurus aga kuni 15 mikronini.
Vaatamata sellistele erinevustele, sisemine struktuur Need organellid on täiesti identsed. Need on moodustatud mikrotuubulitest, mis on oma struktuurilt sarnased rakukeskuse tsentrioolidega.
Motoorsed liikumised tekivad mikrotuubulite omavahelise libisemise tõttu, mille tagajärjel need painduvad. Nende organellide põhjas on basaalkeha, mis kinnitab need raku tsütoplasma külge. Liikumisorganellide toimimise tagamiseks kulutab rakk ATP energiat.

Riis. 2. Lipu ehitus.
Mõned rakud (amööbid, leukotsüüdid) liiguvad pseudopoodide ehk teisisõnu pseudopoodide tõttu. Erinevalt lipudest ja ripsmetest on pseudopoodid aga ajutised struktuurid. Need võivad kaduda ja ilmuda tsütoplasmas erinevatesse kohtadesse. Nende funktsioonid hõlmavad liikumist ning toidu ja muude osakeste püüdmist.
Lipud koosnevad filamendist, konksust ja põhikehast. Vastavalt nende organellide arvule ja asukohale bakterite pinnal need jagunevad:
- Monotrichid(üks flagellum);
- Amfitrihhia(üks flagellum erinevatel poolustel);
- Lophotrichs(hunnik moodustisi ühel või mõlemal poolusel);
- Peritrichous(paljud flagellad paiknevad üle kogu raku pinna).

Riis. 3. Flagellaatide sordid.
Liikumisorganellide funktsioonide hulgas on järgmised:
- liikumise pakkumine üherakulisele organismile;
- lihaste kokkutõmbumisvõime;
- hingamisteede kaitsereaktsioon võõrosakeste eest;
- vedeliku edenemine.
Flagellaadid mängivad suurt rolli keskkonnas leiduvate ainete ringis, paljud neist on head veekogude reostuse näitajad.
Mida me õppisime?
Üks raku koostisosi on liikumisorganellid. Nende hulka kuuluvad lipukesed ja ripsmed, mis moodustuvad mikrotuubulite abil. Nende funktsioonide hulka kuulub üherakulise organismi liikumise tagamine ja vedelike soodustamine mitmerakulise organismi sees.
Test teemal
Aruande hindamine
Keskmine hinne: 4.7. Saadud hinnanguid kokku: 175.
Lihtsamad loomad on üherakulised organismid, omadused, toitumine, esinemine vees ja inimkehas
üldised omadused
Või üherakulised organismid, nagu nende nimigi ütleb, koosnevad ühest rakust. Algloomade hõimkonda kuulub üle 28 000 liigi. Algloomade ehitust saab võrrelda hulkraksete organismide rakkude ehitusega. Mõlemad põhinevad tuumal ja tsütoplasmal koos erinevate organellide (organellide) ja inklusioonidega. Kuid me ei tohi unustada, et paljurakulise organismi mis tahes rakk on osa mis tahes koest või elundist, kus ta täidab oma spetsiifilisi funktsioone. Kõik mitmerakulise organismi rakud on spetsialiseerunud ega ole võimelised iseseisvaks eksisteerimiseks. Seevastu kõige lihtsamad loomad ühendavad raku ja iseseisva organismi funktsioonid. (Füsioloogiliselt ei sarnane algloomarakk mitte mitmerakuliste loomade üksikute rakkudega, vaid terve mitmerakulise organismiga.
Kõige lihtsam iseloomulikud on kõik mis tahes elusorganismidele omased funktsioonid: toitumine, ainevahetus, eritumine, väliste stiimulite tajumine ja neile reageerimine, liikumine, kasv, paljunemine ja surm.
Algloomad Raku struktuur
Tuum ja tsütoplasma, nagu näidatud, on mis tahes raku, sealhulgas üherakuliste loomade peamised struktuursed ja funktsionaalsed komponendid. Viimase kehas on organellid, skeleti- ja kontraktiilsed elemendid ning mitmesugused kandmised. See on alati kaetud rakumembraaniga, enam-vähem õhuke, kuid elektronmikroskoobis selgelt nähtav. Algloomade tsütoplasma on vedel, kuid selle viskoossus varieerub erinevad tüübid ja varieerub sõltuvalt looma seisundist ja edasi keskkond(selle temperatuur ja keemiline koostis). Enamikul liikidel on tsütoplasma läbipaistev või piimvalge, mõnel aga sinise või roheka värvusega (Stentor, Fabrea saliva). Algloomade tuuma ja tsütoplasma keemilist koostist ei ole täielikult uuritud, seda peamiselt nende loomade väiksuse tõttu. On teada, et tsütoplasma ja tuuma alus, nagu kõigil loomadel, koosneb valkudest. Nukleiinhapped Nad on tihedalt seotud valkudega, nad moodustavad nukleoproteiine, mille roll kõigi organismide elus on äärmiselt suur. DNA (desoksüribonukleiinhape) on osa algloomade tuuma kromosoomidest ja tagab päriliku teabe edastamise põlvest põlve. RNA (ribonukleiinhape) leidub algloomades nii tuumas kui ka tsütoplasmas. See rakendab DNA-s kodeeritud üherakuliste organismide pärilikke omadusi, kuna mängib valkude sünteesis juhtivat rolli.
Ainevahetuses osalevad väga olulised tsütoplasma keemilised komponendid – rasvataolised ained lipiidid. Mõned neist sisaldavad fosforit (fosfatiide), paljud on seotud valkudega ja moodustavad lipoproteiinide komplekse. Tsütoplasmas on ka varutoitaineid inklusioonide - tilkade või graanulite kujul. Need on süsivesikud (glükogeen, paramüül), rasvad ja lipiidid. Need toimivad algloomade keha energiavaruna.
Lisaks orgaanilistele ainetele sisaldab tsütoplasma suures koguses vett ja mineraalsooli (katioonid: K+, Ca2+, Mg2+, Na+, Fe3+ ja anioonid: Cl~, P043“, N03“). Algloomade tsütoplasmas leidub palju ainevahetuses osalevaid ensüüme: proteaase, mis tagavad valkude lagunemise; süsivesikud, mis lagundavad polüsahhariide; lipaasid, mis soodustavad rasvade seedimist; suur number ensüümid, mis reguleerivad gaasivahetust, nimelt aluselised ja happelised fosfataasid, oksüdaasid, peroksidaasid ja tsütokroomoksüdaas.
Varasemad ideed algloomade tsütoplasma fibrillaarse, granulaarse või vahtrakulise struktuuri kohta põhinesid fikseeritud ja värvitud preparaatide uuringutel. Uued meetodid algloomade uurimiseks (pimedas väljas, polariseeritud valguses, kasutades intravitaalset värvimist ja elektronmikroskoopiat) on võimaldanud kindlaks teha, et algloomade tsütoplasma on hüdrofiilsete kolloidide (peamiselt valgukomplekside) kompleksne dünaamiline süsteem, millel on vedel või poolvedel konsistents. Ultramikroskoopilisel uurimisel pimedas väljas näib algloomade tsütoplasma optiliselt tühi, nähtavad on vaid raku organellid ja nende kandmised.
Tsütoplasmaatiliste valkude kolloidne olek tagab selle struktuuri muutlikkuse. Tsütoplasmas toimuvad pidevalt muutused valkude agregeeritud olekus: nad lähevad vedelast olekust (sol) tahkemasse, želatiinsemasse olekusse (geel). Need protsessid on seotud ektoplasma tihedama kihi vabanemisega, kesta - pelliikulite moodustumisega ja paljude algloomade amööboidse liikumisega.
Algloomade tuumad, nagu ka mitmerakuliste rakkude tuumad, koosnevad kromatiinmaterjalist, tuumamahlast ning sisaldavad tuumakesi ja tuumaümbrist. Enamik algloomadest sisaldab ainult ühte tuuma, kuid on ka mitmetuumalisi vorme. Sel juhul võivad tuumad olla samad (mitmetuumalised amööbid perekonnast Pelomyxa, mitmetuumalised lipukesed Polymastigida, Opalinida) või erineda kuju ja funktsiooni poolest. Viimasel juhul räägitakse tuumadiferentseerumisest ehk tuumadualismist. Seega iseloomustab kogu ripslaste klassi ja mõningaid foraminifeere tuumade dualism. st tuumad, mis ei ole kuju ja funktsiooni poolest identsed.
Seda tüüpi algloomad, nagu ka teised organismid, järgivad kromosoomide arvu püsivuse seadust. Nende arv võib olla üksik või haploidne (enamik lipulisi ja eosloomi) või kahekordne või diploidne (ripsloomad, opaalid ja ilmselt sarkoodid). Erinevate algloomaliikide kromosoomide arv on väga erinev: 2-4 kuni 100-125 (haploidses komplektis). Lisaks täheldatakse tuumasid, mille kromosoomikomplektide arv on mitmekordselt suurenenud. Neid nimetatakse polüploidseks. On leitud, et ripslaste suured tuumad ehk makrotuumad ja mõnede radiolaariumide tuumad on polüploidsed. On väga tõenäoline, et ka Amoeba proteuse tuum on polüploidne, kromosoomide arv ulatub sellel liigil 500-ni.
Paljundamine Tuumajaotus
Nii algloomade kui ka mitmerakuliste organismide tuuma jagunemise põhitüüp on mitoos ehk karüokinees. Mitoosi ajal toimub kromosomaalse materjali õige ja ühtlane jaotus jagunevate rakkude tuumade vahel. Selle tagab iga kromosoomi pikisuunaline lõhenemine kaheks tütarkromosoomiks mitoosi metafaasis, kusjuures mõlemad tütarkromosoomid lähevad jaguneva raku erinevatele poolustele.
 Monocystis magna gregariini tuuma mitootiline jagunemine:
Monocystis magna gregariini tuuma mitootiline jagunemine: 1, 2 - profaas; 3 - üleminek metafaasile; 4, 5 - metafaas; 6 - varajane anafaas; 7, 8 - hilja
anafaas; 9, 10 - telofaas.
Monocystis magna gregarina tuuma jagunemisel võib täheldada kõiki mitmerakulistele organismidele iseloomulikke mitootilisi kujundeid. Profaasis on tuumas nähtavad niidilaadsed kromosoomid, osa neist on seotud tuumaga (joon. 1, 1, 2). Tsütoplasmas võib eristada kahte tsentrosoomi, mille keskel on radiaalselt lahknevate tähekiirtega tsentrioolid. Tsentrosoomid lähenevad tuumale, külgnevad selle kestaga ja liiguvad tuuma vastaspoolustele. Tuumaümbris lahustub ja moodustub akromatiini spindel (joon. 1, 2-4). Toimub kromosoomide spiraliseerumine, mille tulemusena need lühenevad oluliselt ja kogunevad tuuma keskele, tuum lahustub. Metafaasis liiguvad kromosoomid ekvatoriaaltasandile. Iga kromosoom koosneb kahest kromatiidist, mis asuvad üksteisega paralleelselt ja mida hoiab koos üks tsentromeer. Iga tsentrosoomi ümber olev tähekuju kaob ja tsentrioolid jagunevad pooleks (joonis 1, 4, 5). Anafaasis jagunevad iga kromosoomi tsentromeerid pooleks ja nende kromatiidid hakkavad spindli pooluste suunas lahknema. Algloomadele on iseloomulik, et tsentromeeridele kinnitunud tõmbevõlli niidid on eristatavad vaid mõnel liigil. Kogu spindel on venitatud ja selle keermed, mis kulgevad pidevalt poolusest pooluseni, pikenevad. Kromosoomideks muutunud kromatiidide eraldumise tagavad kaks mehhanismi: nende lahtitõmbumine tõmbespindli keermete kokkutõmbumise toimel ja pidevate spindlikuermete venitamine. Viimane viib raku pooluste üksteisest eemaldamiseni (joonis 1, 6, 7). kromosoomid spiraalivad ja muutuvad õhemaks ning spindel kaob ja jagunenud tsentrioolide ümber moodustub kaks sõltumatut tsentrosoomi, millel on tähekiired – järgmise mitootilise jagunemise tsentrosoomid (joon. 1, ). 9, 10) aga jaguneb ka tsütoplasma, sealhulgas Monocystis'is, mille tulemusena tekivad elutsüklis ajutiselt mitmetuumalised staadiumid iga tuuma ümber ja korraga moodustuvad paljud väikesed rakud.
Eespool kirjeldatud mitoosiprotsessist on erinevaid kõrvalekaldeid: tuumaümbris võib säilida kogu mitootilise jagunemise ulatuses, tuumaümbrise alla võib tekkida akromatiini spindel ja mõnel juhul tsentrioolid ei moodustu. Kõige olulisemad kõrvalekalded on mõnel euglenidae'il: neil puudub tüüpiline metafaas ja spindel läheb tuumast välja. Metafaasis paiknevad kahest kromatiidist koosnevad kromosoomid piki tuuma telge, ekvatoriaalplaati ei moodustu, tuumamembraan ja tuum säilivad, viimane jaguneb pooleks ja läheb tütartuumadesse. Kromosoomide käitumises mitoosis algloomadel ja mitmerakulistel organismidel ei ole põhimõttelisi erinevusi.
Enne uute uurimismeetodite kasutuselevõttu kirjeldati paljude algloomade tuumade jagunemist amitoosi ehk otsese jagunemisena. Tõelise amitoosi all mõistetakse nüüd tuumade jagunemist ilma kromatiidide (kromosoomide) nõuetekohase eraldamiseta tütartuumadeks. Selle tulemusena moodustuvad mittetäielike kromosoomikomplektidega tuumad. Nad ei ole võimelised normaalseks mitootiliseks jagunemiseks. Selliseid tuumajagunemisi on kõige lihtsamates organismides normaalselt raske oodata. Amitoosi täheldatakse valikuliselt enam-vähem patoloogilise protsessina.
Algloomade keha on üsna keeruline. Ühe raku sees toimub selle üksikute osade diferentseerumine, mis täidavad erinevaid funktsioone. Nii nimetati neid algloomade osi analoogselt mitmerakuliste loomade organitega organellideks või organellideks. Seal on liikumise, toitumise, valguse ja muude stiimulite tajumise organellid, eritavad organellid jne.
Liikumine
Algloomade liikumisorganellid on pseudopoodid ehk pseudopoodid, lipud ja ripsmed. Pseudopoodid moodustuvad enamasti liikumise hetkel ja võivad kaduda kohe, kui algloom liigub. Pseudopoodia on algloomade keha ajutised plasmaväljakasvud, millel ei ole püsivat kuju. Nende kest on esindatud väga õhukese (70-100 A) ja elastse rakumembraaniga. Pseudopoodiad on iseloomulikud sarkoodiatele, mõnedele viburloomadele ja eosloomadele.
Lipud ja ripsmed on tsütoplasma väliskihi püsivad väljakasvud, mis on võimelised rütmiliselt liigutama. Nende organellide ülipeent struktuuri uuriti elektronmikroskoobi abil. Leiti, et need on ehitatud peaaegu samamoodi. Lipu ehk tsiliumi vaba osa ulatub välja raku pinnalt.
Sisemine osa on sukeldatud ektoplasmasse ja seda nimetatakse basaalkehaks või blefaroplastiks. Lipu või tsiliumi üliõhukestel lõikudel võib eristada 11 pikisuunalist fibrillit, millest 2 asuvad keskel ja 9 piki perifeeriat (joonis 2). Mõne liigi tsentraalsetel fibrillidel on spiraalsed triibud. Iga perifeerne fibrill koosneb kahest ühendatud torust või subfbrillist. Perifeersed fibrillid lähevad basaalkehasse, kuid tsentraalsed fibrillid ei jõua selleni. Lipumembraan läheb algloomade keha membraani.
Vaatamata ripsmete ja lippude struktuuri sarnasusele, on nende liikumise olemus erinev. Kui lipukesed teevad keerulisi kruviliigutusi, saab ripsmete tööd kõige hõlpsamini võrrelda aerude liikumisega.
Mõne alglooma tsütoplasmas on lisaks basaalkehale ka parabasaalkeha. Basaalkeha on kogu lihas-skeleti süsteemi alus; lisaks reguleerib see algloomade mitootilise jagunemise protsessi. Parabasaalkeha mängib osa algloomade ainevahetuses, kohati see kaob ja võib siis uuesti ilmuda.
Meeleelundid
Algloomadel on võime määrata valguse intensiivsust (valgustatust), kasutades valgustundlikku organelli - ocellust. Merelipulise Chromulina psammobia silma üliõhukese struktuuri uuring näitas, et see sisaldab tsütoplasmasse sukeldatud modifitseeritud lipu.
Tõttu erinevad tüübid toitumine, millest tuleb üksikasjalikult juttu hiljem, on algloomadel väga lai valik seedeorganelle: lihtsatest seedevakuoolidest või vesiikulitest selliste spetsiifiliste moodustisteni nagu rakusuu, suulehter, neel, pulber.
Väljaheidete süsteem
Enamikule algloomadest on iseloomulik võime taluda ebasoodsaid keskkonnatingimusi (ajutiste reservuaaride väljakuivamine, kuumus, külm jne) tsüstidena. Ettevalmistumisel tsütoosiks eraldab algloom märkimisväärse koguse vett, mis toob kaasa tsütoplasma tiheduse suurenemise. Toiduosakeste jäänused visatakse välja, ripsmed ja lipud kaovad ning pseudopoodid tõmmatakse tagasi. Üldine ainevahetus väheneb, moodustub kaitsev kest, mis koosneb sageli kahest kihist. Paljude vormide tsüstide tekkele eelneb varutoitainete kogunemine tsütoplasmasse.
Algloomad ei kaota tsüstides elujõulisust väga pikka aega. Katsetes ületasid need perioodid perekonna Oicomonas (Protomonadida) puhul 5 aastat, Haematococcus pluvialise puhul 8 aastat ja Peridinium cinctum'i puhul ületas tsüstide maksimaalne ellujäämisperiood 16 aastat.
Tsüstidena kanduvad algloomad tuulega märkimisväärsete vahemaade taha, mis seletab algloomade fauna homogeensust kogu maakeral. Seega on tsüstidel mitte ainult kaitsefunktsioon, vaid need on ka algloomade leviku peamised vahendid.
Algloomade keha koosneb tsütoplasmast ja ühest või mitmest tuumast. Tuum on ümbritsetud topeltmembraaniga ja sisaldab kromatiini, mis sisaldab desoksüribonukleiinhapet (DNA), mis määrab raku geneetilise informatsiooni. Enamikul algloomadest on väikese kromatiinisisaldusega vesikulaarne tuum, mis on kogutud piki tuuma perifeeriat või tuumasisesesse kehasse, karüosoomi. Ripslaste mikrotuumad on massiivsed tuumad, milles on palju kromatiini. Enamiku algloomade raku tavalisteks komponentideks on mitokondrid ja Golgi aparaat.
Amoeboidsete vormide (sarcodidae, aga ka mõnede teiste rühmade elufaasid) kehapind on kaetud umbes 100 A paksuse rakumembraaniga. Paljude flagellaatide keha on kaetud periplastiga, mis on moodustatud pikisuunaliste fibrillide jadast, mis on sulanud pelliikliga. Paljudel algloomadel on spetsiaalsed tugifibrillid, näiteks trüpanosoomide ja trihhomonaadide lainelise membraani tugifibrill.
Tihedatel ja kõvadel kestadel on puhkevormid algloomad, tsüstid. Testate amööbid, foraminifera ja mõned teised algloomad on suletud majadesse või kestadesse.
Erinevalt mitmerakulise organismi rakust on algloomarakk terviklik organism. Keha mitmekülgsete funktsioonide täitmiseks saab algloomade kehale spetsialiseeruda struktuursed moodustised ja organellid. Eesmärgi järgi jagunevad algloomade organellid liikumise, toitumise, eritumise jne organellideks.
Algloomade liikumise organellid on väga mitmekesised. Amoeboidsed vormid liiguvad läbi tsütoplasmaatiliste eendite, pseudopoodide moodustumise. Sellist liikumisviisi nimetatakse amoeboidiks ja seda leidub paljudes algloomade rühmades (sarkoodid, eosloomade aseksuaalsed vormid jne). Spetsiaalsed liikumisorganellid on lipud ja ripsmed. Lipud on iseloomulikud lipuliste klassile, aga ka teiste klasside esindajate sugurakud. Enamikus vormides on neid vähe (1 kuni 8). Ripslaste, mis on ripsmete liikumise organellid, arv võib ühes isendis ulatuda mitme tuhandeni. Elektronmikroskoopilised uuringud on näidanud, et algloomade, metaloomade ja taimerakkude lipud ja ripsmed on ehitatud ühe tüübi järgi. Nende aluseks on fibrillide kimp, mis koosneb kahest kesksest ja üheksast paarilisest perifeersest fibrillist.
Žgutt on ümbritsetud membraaniga, mis on rakumembraani jätk. Tsentraalsed fibrillid esinevad ainult nööri vabas osas ja perifeersed fibrillid ulatuvad sügavale tsütoplasmasse, moodustades põhitera - blefaroplasti. Žguti saab ühendada tsütoplasmaga märkimisväärse vahemaa kaugusel õhukese membraani - lainelise membraani abil. Ripslaste tsiliaarne aparaat võib saavutada märkimisväärse keerukuse ja eristuda tsoonideks, mis täidavad iseseisvaid funktsioone. Ripsmed ühinevad sageli rühmadena, moodustades ogasid ja membraane. Iga tsilium algab tsütoplasma pinnakihis asuvast basaalterast, kinetosoomist. Kinetosoomide kogum moodustab infratsiiliatsiooni. Knnetosoomid paljunevad ainult kaheks jagunedes ega saa uuesti tekkida. Lipuaparaadi osalise või täieliku vähenemise korral jääb infratsiilia alles ja seejärel tekivad uued ripsmed.
Algloomade liikumine toimub ajutiste või püsivate liikumisorganellide abil. Esimeste hulka kuuluvad pseudopoodid ehk pseudopoodid - ajutiselt moodustunud ektoplasma väljakasvud, näiteks amööbis, millesse endoplasm näib “voolavat”, mille tõttu tundub, et kõige lihtsam ise “voolab” ühest kohast teise. Püsivad liikumise organellid on piitsad ehk lipud ja ripsmed.
Kõik need organellid on algloomade protoplasma väljakasvud. Žgutil on piki oma telge tihedam elastne niit, mis on justkui vedelama plasma puhul riietatud. Algloomade kehas on nööri põhi ühendatud basaalgraanuliga, mida peetakse tsentrosoomi homoloogiks. Žguti vaba ots tabab ümbritsevat vedelikku, kirjeldades ringikujulisi liigutusi.
Erinevalt ripsmetest on ripsmed väga lühikesed ja äärmiselt arvukad. Ripsmed painduvad kiiresti ühele küljele ja sirutuvad seejärel aeglaselt; nende liikumine toimub järjestikku, mille tõttu vaatleja silm jääb väreleva leegi mulje ja liikumist ennast nimetatakse virvendamiseks.
Mõnel algloomal võivad samaaegselt olla pseudopoodid ja žgutt või pseudopodia ja ripsmed. Teistel algloomadel võib oma elutsükli erinevatel etappidel olla erinev liikumisviis.
Mõne alglooma puhul diferentseeruvad protoplasmas kontraktiilsed kiud ehk müoneemid, tänu mille tööle saab algloomade keha kiiresti kuju muuta.

Esimesel juhul toimub toidu allaneelamine pseudopoodide töö, nn fagotsüütilise toitumise abil, näiteks algloomade tsüstide ja bakterite allaneelamine soolestiku amööbide või ripsmetega, mis ajavad osakesed rakusuhu (tsütostoomi). näiteks ripsloomad Balantidium coll ja tärklise terad). Endosmootne toitumine on iseloomulik algloomadele, millel puuduvad toitumisorganellid, näiteks trüpanosoomid, leishmania, gregariinid, mõned ripsloomad ja paljud teised. jne. Toitumine toimub sellistel juhtudel keskkonnast lahustunud orgaaniliste ainete imendumise tõttu; Seda toitumisviisi nimetatakse ka saprofüütiliseks.
Allaneelatud toitained sisenevad endoplasmasse, kus need seeditakse. Kasutamata jäägid visatakse välja või ükskõik kuhu alglooma keha pinnale või selle teatud piirkonda (analoogselt roojamisprotsessiga).
Algloomade endoplasmas ladestuvad varutoitained glükogeeni, paraglükogeeni (külmas vees ja alkoholis lahustumatu), rasva ja muude ainete kujul.
Endoplasma sisaldab ka eritusaparaati, kui see antud algloomaliigil morfoloogiliselt üldse väljendub. Ekskretsiooni, aga ka osmoregulatsiooni ja osaliselt hingamise organellid on pulseerivad vakuoolid, mis rütmiliselt kokku tõmbudes tühjendavad oma vedelat sisu, mis koguneb endoplasma külgnevatest osadest taas vakuooli. Endoplasmas on algloomade tuum. Paljudel algloomadel on kaks või enam tuuma, millel on erinevates algloomades erinev struktuur.
Tuum on kõige lihtsama vajalik komponent, sest kõik eluprotsessid saavad toimuda ainult selle juuresolekul; Algloomade protoplasma tuumavabad lõigud võivad katsetingimustes püsida vaid mõnda aega.
Algloomadel on spetsiifilisus ka vektorite suhtes. Mõned liigid kohanduvad ainult ühe konkreetse vektoriga, samas kui teistel võib kandjateks olla mitu liiki, mis sageli kuuluvad ühte klassi.