Glikolizės svarba organizmui. Glikolizė. Aš esu žingsnis. Triozės fosfato izomerų susidarymas
Glikolizė yra pagrindinis daugelio gyvų organizmų įvairių angliavandenių katabolizmo procesas. Būtent tai leidžia generuoti energiją ATP molekulių pavidalu tose ląstelėse, kuriose nevyksta fotosintezė. Anaerobinė glikolizė vyksta esant deguoniui arba be jo.
Proceso specifika
Daugybė šio proceso cheminės esmės tyrimų parodė, kad pradiniai fermentacijos ir kvėpavimo etapai turi panašumų. Šio atradimo dėka mokslininkai paaiškino gyvojo pasaulio vienybę. Anaerobinė glikolizė įvyksta pasibaigus trikarboksirūgšties ciklui ir laisvųjų elektronų perkėlimui. Piruvatas patenka į mitochondrijas ir oksiduojamas iki anglies dioksido. Dėl to iš heksozės išsiskiria laisvoji energija. Fermentai, greitinantys glikolitines reakcijas, daugelyje ląstelių randami tirpioje formoje citozolyje. Heksozės oksidacijos proceso, kuris vyksta esant deguoniui, greitintuvai yra susitelkę mitochondrijų membranose.
Norint padalyti angliavandenių molekulę, kurioje yra šeši anglies atomai, į dvi molekules, būtinas dešimties aktyvių fermentų buvimas. Tyrėjai sugebėjo juos identifikuoti gryna forma, tirti jų fizines ir chemines savybes.

Proceso etapai
Glikolizės procesas vyksta gyvų organizmų ląstelėse. Jį lydi piruvo rūgšties susidarymas ir susideda iš kelių etapų. Kvėpavimo takų sutrikimui būtina suaktyvinti gliukozę. Panašus procesas vyksta, kai anglies atomas yra fosforilinamas sąveikaujant su ATP.
gliukozė + ATP = gliukozė-6-fosfatas + ADP
Šiai cheminei sąveikai atlikti naudojami magnio katijonai ir heksokinazė (fermentas). Tada reakcijos produktas izomerizuojamas į fruktozės-6-fosfatą. Kaip katalizatorius naudojamas fermentas fosfogliukoizomerazė.
Gliukozės glikolizei būdingas kitas etapas, kuriame dalyvauja ATP. Fosforo rūgštis pridedama prie pirmojo fruktozės anglies atomo. Tolesni glikolizės etapai yra susiję su susidariusio fruktozės-1,6-difosfato atsiejimu į triozes, PGA (3-fosfogliceraldehido) susidarymu.
Glikolizė yra kelių etapų procesas, susijęs su energijos išsiskyrimu. Suskaidžius vieną gliukozės molekulę, gaunamos dvi PHA molekulės, todėl jos kartojasi.
Glikolizė yra procesas, kurį apibendrina lygtis:
C6H12O6 + 2ATP + 2NAD + 2Pn + 4ADP → 2PVK 2NADH + 2H+ + 4ATP + 2ADP

Glikolizės reguliavimas
Glikolizė yra svarbus procesas gyvam organizmui. Juo siekiama atlikti dvi funkcijas:
- ATP susidarymas skaidant heksozės molekules;
- statybinių blokų transportavimas tolesniam sintezės procesui.
Šio proceso reguliavimas yra skirtas visiškai patenkinti šiuos gyvos ląstelės poreikius. Reakcijos, kurių metu fermentai veikia kaip katalizatoriai, yra negrįžtamos.
Ypač svarbi reguliavimo funkcija yra fosfofruktokinazė. Šis fermentas atlieka reguliavimo funkciją ir apibūdina proceso greitį.

Fiziologinė reikšmė
Glikolizė gali būti laikoma universaliu gliukozės katabolizmo metodu. Jį aktyviai naudoja pro- ir eukariotiniai organizmai. Glikolizę katalizuojantys fermentai ištirpsta vandenyje ir kaupiasi citozolyje. Kai kurios gyvūnų ląstelės ir audiniai gali katabolizuoti heksozę tik glikolizės būdu. Pavyzdžiui, inkstų kanalų ląstelės ir smegenų neuronai turi panašius gebėjimus.
Yra tam tikrų skirtumų tarp fiziologinio glikolizės vaidmens riebaliniame audinyje ir kepenyse. Virškinimo metu riebaliniame audinyje ir kepenyse šis procesas yra substratų, naudojamų riebalų sintezei, šaltinis.
Kai kurie augalų audiniai kaupia krakmolą gumbuose. Vandens augalai energiją gauna glikolizės būdu.
Anaerobinėmis sąlygomis piruvatas virsta etanoliu ir laktatu. Procesą lydi didelis energijos kiekis.
Glikolizė turi didelę fiziologinę reikšmę adipocitams. Su jo pagalba vietoj oksidacinio proceso vyksta lipogenezė, kuri sumažina oksidacinį stresą.

Medicininė reikšmė
Besikaupiant laktatui, kuris susidaro anaerobinėmis sąlygomis, kraujyje vystosi laktatacidozė. Dėl to sumažėja kraujo aplinkos reakcija, kurią lydi dideli ląstelių metabolizmo sutrikimai. Panašus procesas vyksta patologinių procesų, susijusių su sutrikusiu audinių aprūpinimu deguonimi, metu. Pavyzdžiui, sergant miokardo infarktu, kraujavimu, plaučių embolija. Šį procesą sukelia cukrinis diabetas, kai vietoj aerobinės glikolizės vyksta anaerobinis procesas.
Atsižvelgiant į tai, kad insulinas yra glikolizės greitintuvas, sergant 1 tipo cukriniu diabetu glikolizės procesas sulėtėja. Štai kodėl tie vaistai, kurie stimuliuoja šiam procesui naudojamus fermentus, atlieka ligos gydymo funkciją.

Išvada
Glikolizė yra procesas, būtinas visaverčiam organizmų funkcionavimui. Sergant vėžiu, gliukozės suvartojimas padidėja dešimt kartų, todėl naviko ląstelių gyvybingumas priklauso nuo glikolizės.
Išsamiai ištyrę šio proceso ypatumus, mokslininkams pavyko glikolizę panaudoti ne tik ląstelėms maitinti, bet ir tam tikroms ligoms gydyti.
IN anaerobinis procesas piruvo rūgštis redukuojama iki pieno rūgšties (laktato), todėl mikrobiologijoje anaerobinė glikolizė vadinama pieno fermentacija. Laktatas yra metabolinis aklavietė ir tada niekuo nevirsta, vienintelis laktato panaudojimo būdas yra jį oksiduoti atgal į piruvatą.
Daugelis kūno ląstelių gali anaerobiškai oksiduoti gliukozę. Dėl raudonieji kraujo kūneliai tai vienintelis energijos šaltinis. Ląstelės griaučių raumenys dėl gliukozės skaidymo be deguonies jie gali atlikti galingą, greitą, intensyvus darbas, pavyzdžiui, sprintas, įtampa jėgos sporto šakose. Už fizinio aktyvumo ribų gliukozės oksidacija be deguonies ląstelėse padidėja hipoksijos metu – su Įvairios rūšys anemija, adresu kraujotakos sutrikimai audiniuose, nepriklausomai nuo priežasties.
Glikolizė
Anaerobinė gliukozės transformacija yra lokalizuota citozolis ir apima du etapus iš 11 fermentinių reakcijų.
Pirmasis glikolizės etapas
Pirmasis glikolizės etapas yra parengiamieji, čia sunaudojama ATP energija, aktyvuojama ir iš jos susidaro gliukozė triozės fosfatai.
Pirmoji reakcija Glikolizė yra susijusi su gliukozės pavertimu reaktyviu junginiu dėl 6-ojo anglies atomo, neįtraukto į žiedą, fosforilinimo. Ši reakcija yra pirmoji gliukozės konversijos metu, katalizuojama heksokinazės.
Antroji reakcija būtina pašalinti kitą anglies atomą iš žiedo tolesniam jo fosforilinimui (fermentas gliukozės fosfato izomerazė). Dėl to susidaro fruktozė-6-fosfatas.
Trečia reakcija– fermentas fosfofruktokinazė fosforilina fruktozės-6-fosfatą, kad susidarytų beveik simetriška fruktozės-1,6-difosfato molekulė. Ši reakcija yra pagrindinė reguliuojant glikolizės greitį.
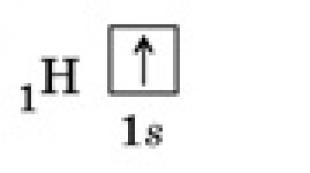
IN ketvirta reakcija fruktozė 1,6-bisfosfatas perpjaunamas per pusę fruktozė-1,6-difosfatas- aldolazės, kad susidarytų du fosforilinti triozės izomerai – aldozės gliceraldehidas(GAF) ir ketozės dioksacetonas(DAF).
Penkta reakcija parengiamasis etapas - gliceraldehido fosfato ir dioksacetono fosfato perėjimas vienas į kitą dalyvaujant triosefosfato izomerazės. Reakcijos pusiausvyra pasislenka dihidroksiacetono fosfato naudai, jo dalis yra 97%, gliceraldehido fosfato dalis yra 3%. Ši reakcija, nepaisant jos paprastumo, lemia tolesnį gliukozės likimą:
- kai ląstelėje trūksta energijos ir suaktyvėja gliukozės oksidacija, dioksiacetono fosfatas paverčiamas gliceraldehido fosfatu, kuris toliau oksiduojamas antrajame glikolizės etape,
- esant pakankamam ATP kiekiui, priešingai, gliceraldehido fosfatas izomerizuojasi į dihidroksiacetono fosfatą, o pastarasis siunčiamas riebalų sintezei.
Antrasis glikolizės etapas

Antrasis glikolizės etapas yra energijos išleidimas, esantis gliceraldehido fosfate, ir saugoti jį formoje ATP.
Šešta reakcija glikolizė (fermentas gliceraldehido fosfato dehidrogenazė) – oksiduojantis gliceraldehido fosfatui ir į jį pridėjus fosforo rūgšties, susidaro didelės energijos junginys iš 1,3-difosfoglicerino rūgšties ir NADH.
IN septintoji reakcija(fermentas fosfoglicerato kinazė) 1,3-difosfoglicerate esančios fosfoesterio jungties energija eikvojama ATP susidarymui. Reakcija gavo papildomą pavadinimą -, kuris paaiškina energijos šaltinį didelės energijos jungtims gauti ATP (iš reakcijos substrato), priešingai nei oksidacinis fosforilinimas (iš elektrocheminis gradientas vandenilio jonai ant mitochondrijų membranos).
Aštunta reakcija– 3-fosfogliceratas, susintetintas ankstesnėje reakcijoje veikiant fosfoglicerato mutazė izomerizuojasi iki 2-fosfoglicerato.
Devinta reakcija– fermentas enolazė abstrahuoja vandens molekulę iš 2-fosfoglicerino rūgšties ir fosfoenolpiruvato sudėtyje susidaro didelės energijos fosfoesterio jungtis.
Dešimtoji reakcija glikolizė yra kita substrato fosforilinimo reakcija– susideda iš didelės energijos fosfato pernešimo piruvatkinaze iš fosfenolpiruvato į ADP ir piruvo rūgšties susidarymo.
bendra apžvalga
Glikolitinis kelias susideda iš 10 nuoseklių reakcijų, kurių kiekvieną katalizuoja atskiras fermentas.
Glikolizės procesą galima suskirstyti į du etapus. Pirmasis etapas, kuris vyksta sunaudojant 2 ATP molekules, susideda iš gliukozės molekulės padalijimo į 2 gliceraldehido-3-fosfato molekules. Antrame etape vyksta nuo NAD priklausoma gliceraldehido-3-fosfato oksidacija, kartu su ATP sinteze. Pati glikolizė yra visiškai anaerobinis procesas ty reakcijoms įvykti nereikia deguonies.
Glikolizė yra vienas iš seniausių medžiagų apykaitos procesų, žinomas beveik visuose gyvuose organizmuose. Manoma, kad pirmykščiuose prokariotuose glikolizė atsirado daugiau nei prieš 3,5 milijardo metų.
Lokalizacija
Eukariotinių organizmų ląstelėse citozolyje yra dešimt fermentų, kurie katalizuoja gliukozės skilimą į PVC, o visi kiti fermentai, susiję su energijos apykaitą, - mitochondrijose ir chloroplastuose. Gliukozė į ląstelę patenka dviem būdais: nuo natrio priklausomu simptomu (daugiausia enterocitams ir inkstų kanalėlių epiteliui) ir palengvinta gliukozės difuzija naudojant baltymus-nešiklius. Šių pernešėjų baltymų darbą kontroliuoja hormonai ir, visų pirma, insulinas. Insulinas stipriausiai stimuliuoja gliukozės transportavimą raumenyse ir riebaliniame audinyje.
Rezultatas
Glikolizės rezultatas yra vienos gliukozės molekulės pavertimas dviem piruvinės rūgšties (PVA) molekulėmis ir dviejų redukuojančių ekvivalentų susidarymas kofermento NAD∙H pavidalu.
Visa glikolizės lygtis yra tokia:
Gliukozė + 2NAD + + 2ADP + 2P n = 2NAD∙H + 2PVK + 2ATP + 2H2O + 2H+.Jei ląstelėje nėra deguonies arba jo trūksta, piruvo rūgštis redukuojama į pieno rūgštį bendroji lygtis glikolizė bus tokia:
Gliukozė + 2ADP + 2P n = 2 laktatas + 2ATP + 2H 2 O.Taigi, anaerobinio vienos gliukozės molekulės skilimo metu, bendra grynoji ATP išeiga yra dvi molekulės, gautos ADP substrato fosforilinimo reakcijose.
Aerobiniuose organizmuose galutiniai glikolizės produktai toliau virsta biocheminiais ciklais, susijusiais su ląstelių kvėpavimu. Dėl to, visiškai oksidavus visus vienos gliukozės molekulės metabolitus paskutiniame ląstelių kvėpavimo etape - oksidacinis fosforilinimas, atsirandantis ant mitochondrijų kvėpavimo grandinė esant deguoniui, kiekvienai gliukozės molekulei susintetinama papildomai po 34 arba 36 ATP molekules.
Kelias
Pirmoji reakcija glikolizė yra fosforilinimas gliukozės molekulės, atsirandančios dalyvaujant audiniams specifiniam fermentui heksokinazei, sunaudojant 1 ATP molekulę; susidaro aktyvi gliukozės forma - gliukozė-6-fosfatas (G-6-F):
Kad reakcija vyktų, terpėje būtinas Mg 2+ jonų buvimas, su kuriuo kompleksiškai susieta ATP molekulė. Ši reakcija yra negrįžtama ir yra pirmoji Pagrindinė glikolizės reakcija.
Gliukozės fosforilinimas turi du tikslus: pirma, dėl to, kad plazminė membrana, pralaidi neutraliai gliukozės molekulei, nepraleidžia neigiamo krūvio turinčių G-6-P molekulių, fosforilinta gliukozė yra užrakinta ląstelės viduje. Antra, fosforilinimo metu gliukozė paverčiama aktyvia forma, kuri gali dalyvauti biocheminėse reakcijose ir būti įtraukta į medžiagų apykaitos ciklus.
Kepenų heksokinazės izofermentas gliukokinazė yra svarbus reguliuojant gliukozės kiekį kraujyje.
Kitoje reakcijoje ( 2 ) veikiant fermentui fosfogliukoizomerazei G-6-P paverčiamas į fruktozė 6-fosfatas (F-6-F):

Šiai reakcijai nereikia energijos ir reakcija yra visiškai grįžtama. Šiame etape fruktozė taip pat gali būti įtraukta į glikolizės procesą fosforilinant.
Po to beveik iš karto viena po kitos vyksta dvi reakcijos: negrįžtamas fruktozės-6-fosfato fosforilinimas ( 3 ) ir susidariusio grįžtamojo aldolinio skilimo fruktozė 1,6-bifosfatas (F-1.6-bF) į dvi trijules ( 4 ).
P-6-P fosforilinimą atlieka fosfofruktokinazė, sunaudojant kitos ATP molekulės energiją; tai antrasis pagrindinė reakcija glikolizės, jos reguliavimas lemia visos glikolizės intensyvumą.

Aldolo skilimas F-1.6-bF atsiranda veikiant fruktozės-1,6-bisfosfato aldolazei:

Dėl ketvirtosios reakcijos dihidroksiacetono fosfatas Ir gliceraldehido-3-fosfatas, o pirmasis yra beveik iš karto paveiktas fosfotriozės izomerazės eina į antrą ( 5 ), kuris dalyvauja tolesnėse transformacijose:

Kiekviena gliceraldehido fosfato molekulė yra oksiduojama NAD+ dalyvaujant gliceraldehido fosfato dehidrogenazė prieš 1,3-difosfogliceratas (6 ):

Toliau su 1,3-difosfogliceratas 1 padėtyje turintis didelės energijos jungtį, fermentas fosfoglicerato kinazė perneša fosforo rūgšties liekaną į ADP molekulę (reakcija 7 ) - susidaro ATP molekulė:

Tai pirmoji substrato fosforilinimo reakcija. Nuo šio momento gliukozės skilimo procesas nustoja būti nepelningas energijos požiūriu, nes energijos sąnaudas pirmojo etapo yra kompensuojamos: vietoj dviejų, sunaudojamų reakcijose, susintetinamos 2 ATP molekulės (po vieną kiekvienam 1,3-difosfogliceratui). 1 Ir 3 . Šiai reakcijai įvykti reikalingas ADP buvimas citozolyje, tai yra, kai ląstelėje yra ATP perteklius (ir trūksta ADP), jos greitis mažėja. Kadangi ATP, kuris nėra metabolizuojamas, ląstelėje nenusėda, o tiesiog sunaikinamas, ši reakcija yra svarbus glikolizės reguliatorius.
Tada nuosekliai: susidaro fosfoglicerolio mutazė 2-fosfogliceratas (8 ):

Enolazės formos fosfenolpiruvatas (9 ):

Galiausiai, antroji ADP substrato fosforilinimo reakcija įvyksta, kai susidaro piruvato ir ATP enolio forma ( 10 ):

Reakcija vyksta veikiant piruvato kinazei. Tai paskutinė pagrindinė glikolizės reakcija. Enolinės piruvato formos izomerizacija į piruvatą vyksta nefermentiniu būdu.
Nuo pat susikūrimo F-1.6-bF Vyksta tik reakcijos, kurios išskiria energiją 7 Ir 10 , kuriame vyksta ADP substrato fosforilinimas.
Tolimesnis vystymas
Galutinis glikolizės metu susidarančio piruvato ir NAD∙H likimas priklauso nuo organizmo ir sąlygų ląstelėje, ypač nuo deguonies ar kitų elektronų akceptorių buvimo ar nebuvimo.
Anaerobiniuose organizmuose piruvatas ir NAD∙H toliau fermentuojami. Pieno rūgšties fermentacijos metu, pavyzdžiui, bakterijose, fermentas laktatdehidrogenazė piruvatą redukuoja į pieno rūgštį. Mielėse panašus procesas yra alkoholinė fermentacija, kai galutiniai produktai yra etanolis ir anglies dioksidas. Taip pat žinomas sviesto rūgšties ir citrinos rūgšties fermentavimas.
Sviesto rūgšties fermentacija:
Gliukozė → sviesto rūgštis + 2 CO 2 + 2 H 2 O.
Alkoholinė fermentacija:
Gliukozė → 2 etanolis + 2 CO 2 .
Citrinų rūgšties fermentacija:
Gliukozė → citrinos rūgštis + 2 H 2 O.
Fermentacija yra svarbi maisto pramonėje.
Aerobuose piruvatas paprastai patenka į trikarboksirūgšties ciklą (Krebso ciklą), o NAD∙H galiausiai oksiduojasi deguonimi kvėpavimo grandinėje mitochondrijose oksidacinio fosforilinimo proceso metu.
Nors žmogaus metabolizmas daugiausia yra aerobinis, intensyviai dirbančiuose skeleto raumenyse vyksta anaerobinė oksidacija. Esant ribotai prieigai prie deguonies, piruvatas paverčiamas pieno rūgštimi, kaip tai vyksta pieno rūgšties fermentacijos metu daugelyje mikroorganizmų:
PVK + NAD∙H + H + → laktatas + NAD + .
Raumenų skausmas, atsirandantis kurį laiką po neįprastai intensyvaus fizinio krūvio, yra susijęs su juose besikaupiančia pieno rūgštimi.
Pieno rūgšties susidarymas yra aklavietės metabolizmo atšaka, tačiau tai nėra galutinis metabolizmo produktas. Veikiant laktatdehidrogenazei, pieno rūgštis vėl oksiduojasi, susidaro piruvatas, kuris dalyvauja tolesnėse transformacijose.
Glikolizės reguliavimas
Yra vietinis ir bendras reglamentas.
Vietinis reguliavimas atliekamas keičiant fermentų aktyvumą, veikiant įvairiems metabolitams ląstelės viduje.
Glikolizės reguliavimas kaip visuma, iš karto visam organizmui, vyksta veikiant hormonams, kurie, veikdami per antrinių pasiuntinių molekules, keičia tarpląstelinį metabolizmą.
Insulinas vaidina svarbų vaidmenį skatinant glikolizę. Gliukagonas ir adrenalinas yra svarbiausi hormoniniai glikolizės inhibitoriai.
Insulinas skatina glikolizę:
- heksokinazės reakcijos aktyvinimas;
- fosfofruktokinazės stimuliavimas;
- piruvato kinazės stimuliavimas.
Kiti hormonai taip pat turi įtakos glikolizei. Pavyzdžiui, somatotropinas slopina glikolitinių fermentų veiklą, o skydliaukės hormonai yra stimuliatoriai.
Glikolizė reguliuojama keliais pagrindiniais etapais. Heksokinazės katalizuojamos reakcijos ( 1 ), fosfofruktokinazė ( 3 ) ir piruvato kinazė ( 10 ) pasižymi dideliu laisvosios energijos sumažėjimu ir yra praktiškai negrįžtami, todėl jie yra veiksmingi glikolizės reguliavimo taškai.
Heksokinazės reguliavimas
Heksokinazė slopina reakcijos produktas – gliukozė-6-fosfatas, kuris allosteriškai jungiasi su fermentu, pakeisdamas jo aktyvumą.
Dėl to, kad didžioji dalis G-6-P ląstelėje susidaro skaidant glikogeną, heksokinazės reakcija iš tikrųjų nėra būtina glikolizei, o gliukozės fosforilinimas nėra išskirtinės reikšmės reguliuojant. glikolizės. Heksokinazės reakcija yra svarbus žingsnis reguliuojant gliukozės koncentraciją kraujyje ir ląstelėje.
Fosforilinta gliukozė praranda gebėjimą pernešti per membraną nešiklio molekulėmis, todėl susidaro sąlygos jai kauptis ląstelėje. G-6-P heksokinazės slopinimas riboja gliukozės patekimą į ląstelę, užkertant kelią pernelyg dideliam jos kaupimuisi.
Gliukozės-6-fosfatas neslopina kepenų gliukokinazės (IV heksokinazės izotipo), o kepenų ląstelės ir toliau kaupia gliukozę net ir su dideliu G-6-P kiekiu, iš kurio vėliau sintetinamas glikogenas. Palyginti su kitais izotipais, gliukokinazė išsiskiria didele Michaelio konstantos reikšme, tai yra, fermentas visu pajėgumu veikia tik esant didelei gliukozės koncentracijai, kuri beveik visada būna po valgio.
Gliukozės-6-fosfatas gali būti paverstas atgal į gliukozę, veikiant gliukozės-6-fosfatazei. Fermentai gliukokinazė ir gliukozės-6-fosfatazė dalyvauja palaikant normalią gliukozės koncentraciją kraujyje.
Fosfofruktokinazės reguliavimas
Fosfofruktokinazės reakcijos intensyvumas turi lemiamą įtaką visam glikolizės pralaidumui, o fosfofruktokinazės stimuliavimas laikomas svarbiausiu reguliavimo etapu.
Fosfofruktokinazė (PFK) yra tetramerinis fermentas, kuris pakaitomis egzistuoja dviejose konformacinėse būsenose (R ir T), kurios yra pusiausvyroje ir pakaitomis pereina iš vienos į kitą. ATP yra ir substratas, ir alosterinis FFK inhibitorius.
Kiekvienas iš FFK subvienetų turi dvi ATP surišimo vietas: substrato vietą ir slopinimo vietą. Substrato vieta vienodai gali prijungti ATP bet kurioje tetramero konformacijoje. Nors slopinimo vieta jungiasi tik su ATP, kai fermentas yra T konformacijos būsenoje, kitas PPA substratas yra fruktozė 6-fosfatas, kuris pirmiausia jungiasi su fermentu R būsenoje. Esant didelei ATP koncentracijai, slopinimo vieta yra užimta, perėjimai tarp fermentų konformacijų tampa neįmanomi, dauguma fermentų molekulių stabilizuojamos T būsenoje, negali prijungti P-6-P. Tačiau fosfofruktokinazės slopinimą ATP slopina AMP, kuris jungiasi su fermento R konformacijomis, taip stabilizuodamas P-6-P prisijungimo fermento būseną.
Svarbiausias allosterinis glikolizės ir gliukoneogenezės reguliatorius yra fruktozė 2,6-bifosfatas, kuri nėra tarpinė šių ciklų grandis. Fruktozė 2,6-bisfosfatas allosteriškai aktyvina fosfofruktokinazę.
Fruktozės-2,6-bifosfato sintezę katalizuoja specialus bifunkcinis fermentas – fosfofruktokinazė-2/fruktozė-2,6-bifosfatazė (PFK-2/F-2,6-BPase). Nefosforilintas baltymas yra žinomas kaip fosfofruktokinazė-2 ir turi katalizinį poveikį fruktozės 6-fosfatui, sintetindamas fruktozę 2-6-bifosfatą. Dėl to žymiai stimuliuojamas FPA aktyvumas ir stipriai slopinamas fruktozės-1,6-bifosfatazės aktyvumas. Tai yra, esant FFK-2 aktyvumui, šios reakcijos tarp glikolizės ir gliukoneogenezės pusiausvyra pasislenka į pirmąją - sintetinamas fruktozė-1,6-bifosfatas.
Fosforilintas bifunkcinis fermentas neturi kinazės aktyvumo, priešingai, jo molekulėje suaktyvinama vieta, kuri hidrolizuoja P2,6BP į P6P ir neorganinį fosfatą. Bifunkcinio fermento fosforilinimo metabolinis poveikis yra toks, kad nutrūksta allosterinė PPA stimuliacija, pašalinamas allosterinis F-1,6-BPazės slopinimas ir pusiausvyra pereina į gliukoneogenezę. Gaminamas F6P, o vėliau – gliukozė.
Bifunkcinio fermento tarpusavio konversijas vykdo nuo cAMP priklausoma proteinkinazė (PK), kurią savo ruožtu reguliuoja kraujyje cirkuliuojantys peptidiniai hormonai.
Sumažėjus gliukozės koncentracijai kraujyje, slopinamas ir insulino susidarymas, o gliukagono išsiskyrimas, atvirkščiai, skatinamas, o jo koncentracija kraujyje smarkiai padidėja. Gliukagonas (ir kiti kontrainsuliniai hormonai) jungiasi prie receptorių plazmos membrana kepenų ląstelės, sukeldamos membranos adenilato ciklazės aktyvavimą. Adenilato ciklazė katalizuoja ATP pavertimą cikliniu AMP. cAMP prisijungia prie reguliuojančio baltymo kinazės subvieneto, sukeldamas katalizinių jo subvienetų, kuriuos fosforilina daugybė fermentų, įskaitant bifunkcinę FFK-2/F-2,6-BPazę, atpalaidavimą ir aktyvavimą. Tuo pačiu metu sustoja gliukozės suvartojimas kepenyse ir suaktyvėja gliukoneogenezė bei glikogenolizė, atkuriama normoglikemija.
Piruvato kinazė
Kitas žingsnis, kuriame vykdomas glikolizės reguliavimas, yra paskutinė reakcija – piruvatkinazės veikimo stadija. Taip pat buvo aprašyta keletas izofermentų, turinčių reguliavimo ypatybes piruvato kinazei.
Kepenų piruvato kinazė(L tipo) yra reguliuojamas fosforilinimo, allsterinių efektorių ir genų ekspresijos reguliavimu. Fermentą slopina ATP ir acetil-CoA, o aktyvuoja fruktozė-1,6-bisfosfatas. Piruvatkinazės slopinimas ATP yra panašus į ATP poveikį PPA. ATP prisijungimas prie fermento slopinimo vietos sumažina jo afinitetą fosfenolpiruvatui. Kepenų piruvato kinazę fosforilina ir slopina baltymų kinazė, todėl ji taip pat yra kontroliuojama hormonų. Be to, kepenų piruvato kinazės aktyvumas reguliuojamas kiekybiškai, tai yra, keičiant jos sintezės lygį. Tai lėtas, ilgalaikis reguliavimas. Padidėjęs angliavandenių kiekis maiste skatina piruvatkinazę koduojančių genų ekspresiją, todėl ląstelėje padidėja fermento lygis.
M tipo piruvato kinazė, randamas smegenyse, raumenyse ir kituose gliukozės reikalaujančiuose audiniuose, nereguliuoja proteinkinazė. Tai esminga, nes šių audinių metabolizmą lemia tik vidiniai poreikiai ir nepriklauso nuo gliukozės kiekio kraujyje.
Raumenų piruvato kinazės neveikia išoriniai veiksniai, tokie kaip sumažėjęs gliukozės kiekis kraujyje ar hormonų išsiskyrimas. Ekstraląstelinės sąlygos, sukeliančios fosforilinimą ir kepenų izofermento slopinimą, nekeičia M tipo piruvatkinazės aktyvumo. Tai yra, glikolizės intensyvumą dryžuotuose raumenyse lemia tik ląstelės viduje esančios sąlygos ir nepriklauso nuo bendro reguliavimo.
Reikšmė
Glikolizė yra išskirtinės svarbos katabolinis kelias. Jis suteikia energijos ląstelių reakcijoms, įskaitant baltymų sintezę. Glikolizės tarpiniai produktai naudojami riebalų sintezei. Piruvatas taip pat gali būti naudojamas alaninui, aspartatui ir kitiems junginiams sintetinti. Glikolizės dėka mitochondrijų veikla ir deguonies prieinamumas neriboja raumenų jėgos trumpalaikių ekstremalių apkrovų metu.
taip pat žr
Nuorodos
- Glikolizė
Anaerobinė glikolizė – tai sudėtingas fermentinis nuoseklių gliukozės virsmų procesas, vykstantis žmogaus ir gyvūnų audiniuose nenaudojant deguonies (28 pav.).
Grįžtamąjį piruvo rūgšties pavertimą pieno rūgštimi katalizuoja laktatdehidrogenazė:
Bendras glikolizės rezultatas išreiškiamas tokia lygtimi: C 6 H 12 O 6 + 2H 3 PO 4 + 2ADP = 2C 3 H 6 O 3 + 2ATP + 2H 2 O
Taigi grynoji ATP išeiga anaerobinės glikolizės metu yra 2 mol ATP 1 moliui gliukozės. Būtent anaerobinės glikolizės dėka žmogaus ir gyvūno organizmas tam tikrą laiką deguonies trūkumo sąlygomis gali atlikti daugybę fiziologinių funkcijų.
Šis procesas bakterijose vadinamas pieno fermentacija: juo gaminami fermentuoti pieno produktai. Anaerobinė glikolizė vyksta ląstelių citozolyje, kuriame yra visi tam reikalingi fermentai, ir jai nereikia mitochondrijų kvėpavimo grandinės. ATP anaerobinės glikolizės procese susidaro dėl substrato fosforilinimo reakcijų.
Mielėse anaerobinėmis sąlygomis vyksta panašus procesas - alkoholinė fermentacija, šiuo atveju piruvinė rūgštis dekarboksilinama, kad susidarytų acetaldehidas, kuris vėliau redukuojamas į etilo alkoholį:
CH3-CO-COOH → CH3-CHO + CO2;
CH3-CHO + NAD.H+H+ → CH3-CH2-OH + NAD+.

28 pav. Anaerobinės gliukozės glikolizės schema
10.6. Aerobinis gliukozės skilimas
Aerobinis gliukozės skilimas susideda iš trijų etapų:
1) gliukozės pavertimas piruvo rūgštimi (piruvatu) – aerobinė glikolizė. Ši dalis yra panaši į aukščiau aptartą anaerobinės glikolizės procesą, išskyrus paskutinę jo stadiją (piruvato pavertimą pieno rūgštimi);
2) bendras katabolizmo kelias;
3) mitochondrijų elektronų transportavimo grandinė – audinių kvėpavimo procesas.
Bendras katabolizmo kelias
Bendras katabolizmo kelias susideda iš dviejų etapų.
1 etapas – oksidacinis piruvo rūgšties dekarboksilinimas. Tai sudėtingas daugiapakopis procesas, katalizuojamas daugelio fermentų sistemos – piruvato dehidrogenazės komplekso; lokalizuota mitochondrijose (vidinėje membranoje ir matricoje) ir gali būti išreikšta apibendrinta bendra schema:
CH 3 -CO-COOH + HS-KoA + NAD + → CH 3 -CO-SkoA + NADH + H + + CO 2.
2 etapas – Krebso ciklas (citrato ciklas, arba trikarboksilo ir dikarboksilo rūgščių ciklas) (29 pav.); lokalizuota mitochondrijose (matricoje). Šiame cikle acetilo liekana, įtraukta į acetil-CoA, sudaro keletą pirminių vandenilio donorų. Tada vandenilis, dalyvaujant dehidrogenazėms, patenka į kvėpavimo grandinę. Dėl susieto citrato ciklo ir kvėpavimo grandinės veikimo acetilo liekana oksiduojasi iki CO 2 ir H 2 O. Bendra visos gliukozės virsmų sekos aerobinio skilimo metu lygtis yra tokia:
C 6 H 12 O 2 + 6 O 2 → 6 CO 2 + 6 H 2 O
Aerobinio skilimo energetinis poveikis yra 38 ATP molekulių sintezė, suskaidžius 1 gliukozės molekulę. Taigi energetine prasme visiškas gliukozės oksidavimas į anglies dioksidą ir vandenį yra efektyvesnis procesas nei anaerobinė glikolizė. Deguonis slopina anaerobinę glikolizę, todėl, esant deguonies pertekliui, augalų ir gyvūnų audiniuose stebimas perėjimas nuo anaerobinės glikolizės (fermentacijos) į kvėpavimą (aerobinę glikolizę), t.y. ląstelių perjungimas į efektyvesnį ir ekonomiškesnį energijos gavimo būdą (Pastero efektas). Anaerobinės glikolizės vaidmuo aprūpinant organizmą energija ypač didelis trumpalaikio intensyvaus darbo metu, kai deguonies transportavimo į mitochondrijas mechanizmo galios neužtenka aerobinei glikolizei užtikrinti. Taigi bėgimą ~ 30 sekundžių (200 m) visiškai užtikrina anaerobinė glikolizė, tuo tarpu anaerobinės glikolizės greitis didėjant kvėpavimui mažėja, o aerobinio irimo greitis didėja. Po 4-5 min. bėgimas (1,5 km) – pusę energijos suteikia anaerobinis procesas, pusę – aerobinis procesas. Po 30 min. (10 km bėgimas) – beveik visa energija tiekiama aerobiniu procesu.
Raudonieji kraujo kūneliai iš viso neturi mitochondrijų, o jų ATP poreikį visiškai patenkina anaerobinė glikolizė.
(iš graikų glykys – saldus ir lizė – skilimas, skilimas) – vienas iš trijų pagrindinių (glikolizės, Krebso ciklas ir Entnerio-Dudorofo kelias) energijos gamybos gyvuose organizmuose būdų. Tai anaerobinis (t. y. nereikalaujantis laisvojo O 2) fermentinio nehidrolizinio angliavandenių (daugiausia gliukozės ir glikogeno) skaidymo gyvūnų audiniuose procesas, lydimas adenozino trifosforo rūgšties (ATP) sintezės ir baigiasi pieno rūgšties susidarymas. Glikolizė svarbi raumenų ląstelėms, spermatozoidams ir augantiems audiniams (įskaitant navikus), nes užtikrina energijos kaupimąsi, kai nėra deguonies. Bet žinoma ir glikolizė esant O2 (aerobinė glikolizė) – raudonuosiuose kraujo kūneliuose, akies tinklainėje, vaisiaus audiniuose iškart po gimimo ir žarnyno gleivinėje. G. ir K. Corey, taip pat tokie biochemijos pradininkai kaip O. Meyerhoffas ir G. Embdenas labai prisidėjo prie glikolizės tyrimo. Glikolizė buvo pirmoji visiškai iššifruota biocheminių reakcijų seka (nuo XIX a. pabaigos iki 1940 m.). Heksozės monofosfato šunto arba pentozės fosfato kelias kai kuriose ląstelėse (eritrocituose, riebaliniame audinyje) taip pat gali atlikti energijos tiekėjo vaidmenį.
Be gliukozės, glikolizės procese gali dalyvauti glicerolis, kai kurios aminorūgštys ir kiti substratai. Raumeniniame audinyje, kur pagrindinis glikolizės substratas yra glikogenas, procesas prasideda 2 ir 3 reakcijomis ( cm. schema) ir vadinama glikogenolize. Dažnas tarpinis produktas tarp glikogenolizės ir glikolizės yra gliukozės-6-fosfatas. Atvirkštinis glikogeno susidarymo kelias vadinamas glikogeneze.
Glikolizės metu susidarę produktai yra substratai vėlesnėms oksidacinėms transformacijoms ( cm. Trikarboksirūgšties ciklas arba Krebso ciklas). Procesai, panašūs į glikolizę, yra pieno rūgšties, sviesto rūgšties, alkoholio ir glicerolio fermentacija, kuri vyksta augalų, mielių ir bakterijų ląstelėse. Atskirų glikolizės etapų intensyvumas priklauso nuo rūgštingumo – pH – pH (optimalus pH 7-8), temperatūros ir terpės joninės sudėties. Glikolizės reakcijų seka ( cm. schema) buvo gerai ištirtas ir nustatyti tarpiniai produktai. Tirpūs glikolitiniai fermentai, esantys ląstelių sultyse, išskiriami kristaline arba išgryninta forma.
Fermentai, atliekantys atskirus glikolizės etapus:
1. Heksokinazė KF2.7.1.1 (arba gliukokinazė KF2.7.1.2)
2. Glikogeno fosforilazė KF2.4.1.1
3. Fosfogliukomutazė KF2.7.5.1
4. Gliukozės fosfato izomerazė KF5.3.1.9
5. Fosfofruktokinazė KF2.7.1.11
6. Fruktozės bisfosfato aldolazė KF4.1.2.13
7. Triofosfato izomerazė KF5.3.1.1
8, 9. Gliceraldehido fosfato dehidrogenazė KF1.2.1.12
10. Fosfogliceratkinazė KF2.7.2.3
11. Fosfogliceromutazė KF2.7.5.3
12. Enolazė KF4.2.1.11
13. Piruvato kinazė KF2.7.1.40
14. Laktato dehidrogenazė KF1.1.1.27
Glikolizė prasideda nuo cukrų fosforo darinių susidarymo, kuris prisideda prie ciklinės substrato formos pavertimo acikline, labiau reaktyvia. Viena iš reakcijų, reguliuojančių glikolizės greitį, yra 2 reakcija, katalizuojama fermento fosforilazės. Pagrindinis reguliavimo vaidmuo glikolizėje priklauso fermentui fosfofruktokinazei (5 reakcija), kurio aktyvumą slopina ATP ir citratas, tačiau stimuliuoja jo skilimo produktai. Centrinė glikolizės grandis yra glikolitinė oksidoredukcija (8–10 reakcijos), tai yra redokso procesas, vykstantis oksiduojant 3-fosfogliceraldehidą į 3-fosfoglicerino rūgštį ir redukuojant kofermentą nikotinamido adenino dinukleotidą (NAD). Šias transformacijas atlieka 3-fosfogliceraldehido dehidrogenazė (DPGA), dalyvaujant fosfoglicerato kinazei. Tai vienintelis oksidacinis glikolizės etapas, tačiau jai nereikia laisvo deguonies, reikalingas tik NAD + buvimas, kuris redukuojamas iki NAD-H 2.
Dėl oksidoredukcijos (redokso proceso) išsiskiria energija, kuri kaupiasi (energetinio junginio ATP pavidalu) substrato fosforilinimo procese. Antroji reakcija, kuri užtikrina ATP susidarymą, yra 13 reakcija – piruvo rūgšties susidarymas. Anaerobinėmis sąlygomis glikolizė baigiasi pieno rūgšties susidarymu (14 reakcija), veikiant laktato dehidrogenazei ir dalyvaujant redukuotam NAD, kuris oksiduojamas į NAD (NAD-H 2) ir vėl gali būti naudojamas oksidaciniame etape. . Aerobinėmis sąlygomis piruvo rūgštis oksiduojasi mitochondrijose Krebso ciklo metu.
Taigi, suskaidžius 1 molekulę gliukozės, susidaro 2 molekulės pieno rūgšties ir 4 molekulės ATP. Tuo pačiu metu pirmose glikolizės stadijose (žr. 1, 5 reakcijas) 1 gliukozės molekulei sunaudojama 2 ATP molekulės. Glikogenolizės procese susidaro 3 ATP molekulės, nes nereikia eikvoti ATP, kad susidarytų gliukozės-6-fosfatas. Pirmosios devynios glikolizės reakcijos atspindi jos endergoninę (energijos absorbcijos) fazę, o paskutinės devynios reakcijos – eksergoninę (energijos išsiskyrimo) fazę. Glikolizės proceso metu išsiskiria tik apie 7% teorinės energijos, kurią galima gauti visiškai oksiduojant gliukozę (iki CO 2 ir H 2 O). Tačiau bendras energijos kaupimo ATP pavidalu efektyvumas yra 35–40%, o praktiškomis korinio ryšio sąlygomis jis gali būti didesnis.
Gliceraldehido fosfato dehidrogenazė ir laktatdehidrogenazė yra susietos viduje (vienai reikia NAD +, kitai gamina NAD +), kuri užtikrina šio kofermento cirkuliaciją. Tai gali būti pagrindinė galutinės dehidrogenazės biocheminė reikšmė.
Visos glikolizės reakcijos yra grįžtamos, išskyrus 1, 5 ir 13. Tačiau iš jų fosforo darinių galima gauti gliukozę (reakcija 1) arba fruktozės monofosfatą (5 reakcija) fosforo rūgšties hidroliziniu pašalinimu, esant atitinkamiems fermentams; 13 reakcija yra praktiškai negrįžtama, matyt, dėl aukšta energija fosforo grupės hidrolizė (apie 13 kcal/mol). Todėl gliukozės susidarymas iš glikolizės produktų vyksta kitu būdu.
Esant O 2, glikolizės greitis mažėja (Pastero efektas). Yra pavyzdžių, kai kai kuriuose intensyviai glikolizuojančiuose audiniuose glikolizės būdu slopinamas audinių kvėpavimas (Crabtree efektas). Anaerobinių ir aerobinių oksidacinių procesų ryšio mechanizmai iki galo neištirti. Vienu metu vykstantis glikolizės ir glikogenezės procesų reguliavimas vienareikšmiškai lemia anglies srautą kiekvienu iš šių takų, priklausomai nuo organizmo poreikių. Kontrolė vykdoma dviem lygiais – hormoniniu (aukštesniems gyvūnams per reguliavimo kaskadas, dalyvaujant antriesiems pasiuntiniams) ir metaboliniu (visuose organizmuose).
Igoris Rapanovičius