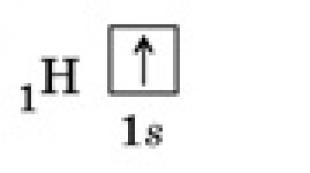
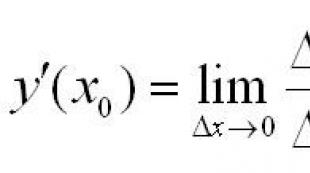Kinetiken för enzymatiska reaktioner. Beroende av hastigheten för enzymatiska reaktioner på koncentrationen av substrat, enzymer, temperatur Vilken är ordningen för den enzymatiska reaktionen med enzym
Hastigheten för enzymatiska reaktioner beror på koncentrationen av sub-
skikt Detta beroende är komplext, vilket för vissa enzymer beskrivs av en parabolisk kurva (fig. 29).
Figur 29 – Beroende av hastigheten för enzymatisk reaktion
på substratkoncentration
Beroendets paraboliska natur förklaras av det faktum att när enzymet interagerar med substratet bildas ett enzym-substratkomplex. Inledningsvis, när substratkoncentrationen ökar, ökar koncentrationen av enzym-substratkomplex i reaktionsblandningen, vilket visar sig i en parallell ökning av reaktionshastigheten. Vid en viss koncentration av substratet (mättande) inträffar en sorts "mättnad" av alla aktiva centra av enzymmolekyler i reaktionsblandningen. Hastigheten för den enzymatiska reaktionen vid mättande koncentration blir maximal. Med en ytterligare ökning av substrathalten i reaktionsblandningen förändras den inte.
Från grafen över beroendet av hastigheten för en enzymatisk reaktion på koncentrationen av substratet, beräknas två viktiga indikatorer:
1. Maximal reaktionshastighet (V max). Det definieras som reaktionshastigheten vid substratets mättande koncentration. Det maximala hastighetsvärdet återspeglar enzymets katalytiska kraft. Större enzymer V max är mer kraftfulla katalysatorer. De katalyserar omvandlingen av ett större antal substratmolekyler per tidsenhet. Den maximala hastigheten uttrycks av antalet varv av enzymet. Omsättningstalet uppskattas av antalet substratmolekyler som omvandlas av enzymet per tidsenhet (s -1). För de flesta enzymer ligger omsättningstalet inom 10 4. Samtidigt finns det enzymer för vilka fart betydligt mer (600 000 för karbanhydras) eller mindre än detta värde (100 för kymotrypsin).
2. Michael är konstant (TILL m). Michaelis-konstanten är koncentrationen av substrat vid vilken reaktionshastigheten är hälften av maximal. Magnitud TILL m reflekterar enzymets affinitet för substratet. Ju högre detta värde är, desto mindre affinitet har enzymet för substratet. TILL m uttrycks i mol substrat. Alltså värdet TILL m i förhållande till glukos för glukokinasenzymet är 10 mmol och för hexokinas - 0,01 mmol. Hexokinas uppvisar en större affinitet för glukos än glukokinas vid samma substratkoncentration, det katalyserar fosforyleringen av glukos i en högre hastighet.
Baserat på en matematisk analys av kurvan för beroendet av hastigheten för en enzymatisk reaktion på koncentrationen av substratet, härledde L. Michaelis och M. Menten (1913) en formel som gör att man kan utvärdera sambandet mellan reaktionshastigheten, den maximala hastigheten och Michaelis-konstanten. Den definieras för närvarande som Michaelis-Menten-ekvationen.
V o = V max [ S]/K m + [ S],
Var V o – reaktionshastighet, S– substratkoncentration.
Allmänna egenskaper hos enzymer
Trots förekomsten av vissa skillnader i struktur, funktion och intracellulär lokalisering kännetecknas enzymer av ett antal gemensamma egenskaper. Dessa inkluderar beroendet av manifestationen av deras katalytiska aktivitet på temperatur (termisk labilitet) och pH i miljön, såväl som substratspecificitet.
En karakteristisk egenskap hos enzymer är termolabilitet. Detta fenomen kan illustreras med en graf över hastigheten för en enzymatisk reaktion mot reaktionsblandningens temperatur (fig. 30).

Figur 30 – Beroende av hastigheten för enzymatisk reaktion på temperaturen
reaktionsmedium ( t opt – optimal temperatur; V- hastighetsreaktion)
Som framgår av den presenterade grafen, vid en temperatur nära 4 o C, sker praktiskt taget inga enzymatiska reaktioner. Av denna anledning kan biologiska föremål förvaras i kyla under en viss tid innan man genomför biokemiska studier. Det är kylan som gör att livsmedelsprodukter kan bevaras från autolys (självsmältning).
En ökning av temperaturen åtföljs av en ökning av hastigheten för den enzymatiska reaktionen. Anledningen till detta är en ökning av den kinetiska energin hos substratet och enzymmolekylerna, vilket ökar interaktionshastigheten mellan dem. Ett liknande fenomen observeras upp till en temperatur som motsvarar enzymets temperaturoptimum. Enzymets temperaturoptimum motsvarar den temperatur vid vilken hastigheten för den enzymatiska reaktionen är maximal. För enzymer från varmblodiga djur är det vanligtvis 28 o C eller 37 o C.
En ytterligare ökning av reaktionsblandningens temperatur leder till en gradvis minskning av hastigheten för den enzymatiska reaktionen. Detta fenomen beror på processen för termisk denaturering av proteinpolypeptidkedjan. Denaturering åtföljs av en förändring i strukturen av enzymets aktiva centrum, vilket resulterar i en minskning av enzymets affinitet för substratet. Vid temperaturer över 55 o C förlorar de flesta enzymer helt sina katalytiska egenskaper (inaktiveras). I detta avseende används uppvärmning till 55–56 o C i stor utsträckning för pastöriseringsproceduren, vilket ökar hållbarheten för livsmedelsprodukter (mjölk, etc.).
Miljöns pH har stor inverkan på hastigheten för den enzymatiska reaktionen. Som framgår av den visade figuren. 31 grafen, liknar den till formen en graf över beroendet av hastigheten för en enzymatisk reaktion på temperaturen.

Figur 31 – Hastighetsberoende ( V) enzymatisk reaktion
på miljöns pH (pH opt – enzymets pH-optimum)
En kraftig minskning av hastigheten för enzymatisk reaktion vid extrema pH-värden är associerad med fenomenet denaturering av polypeptidkedjan i en proteinmolekyl under påverkan av syror och alkalier. Enzymet uppvisar maximal katalytisk kraft vid ett pH-värde som bestäms av termen pH-optimum enzym. De flesta kända enzymer har ett optimalt pH i intervallet från 5,0 till 7,5. Samtidigt finns det många exempel på enzymer där det optimala pH-värdet flyttas till området med sura eller alkaliska pH-värden. Dessa enzymer inkluderar:
Orsaken till förekomsten av ett beroende av hastigheten för enzymatiska reaktioner på pH beror på det faktum att mediets pH-värde har en uttalad effekt på graden av jonisering av de funktionella grupperna i substratet. Funktioner för jonisering av bärnstenssyramolekylen vid olika surhetsgrad i miljön (pH):

Samtidigt påverkar miljöns pH också graden av jonisering av aminosyraradikaler som utgör enzymets aktiva centrum:

Om bildningen av enzym-substratkomplexet stabiliseras på grund av elektrostatiska interaktioner, blir pH:s roll för att ge optimala förhållanden för den enzymatiska reaktionens förlopp tydlig (fig. 24).
Hastigheten för reaktioner som katalyseras av enzymer, i vilkas interaktion med substrat elektrostatiska interaktioner inte är signifikanta, beror i mindre utsträckning på mediets pH. I fig. Figur 32 visar beroendet av hastigheten för proteinhydrolys av papain. I interaktionen av detta enzym med substratet blir hydrofoba interaktioner av primär betydelse. Som kan ses från den presenterade grafen har papain i allmänhet inte ett klart definierat pH-optimum.

Figur 32 - Effekt av pH på hastigheten för proteinhydrolys av papain.
Enzymer har en viss specificitet angående substrat. Specificitet avser enzymers förmåga att katalysera transformationen av ett eller en grupp av strukturellt liknande substrat. Det finns flera typer av enzymspecificitet.
· Absolut specificitet. Det hänvisar till förmågan hos ett enzym att katalysera omvandlingen av endast ett substrat. Enzymer med absolut specificitet inkluderar arginas, urikasrestriktionsenzymer, etc.
· Relativ specificitet. Det betyder förmågan hos ett enzym att katalysera omvandlingen av en grupp av substrat liknande struktur (så kallade proteolytiska enzymer hydrolyserar olika proteiner, lipasestrar av glycerol och högre fettsyror, hexokinas fosforylerar olika monosackarider). I det här fallet bestäms specificiteten av att enzymet endast påverkar en viss typ av bindning (proteolytiska enzymer hydrolyserar peptidbindningen, lipas hydrolyserar esterbindningen etc.).
· Stereospecificitet . Denna term hänvisar till förmågan hos ett enzym att katalysera omvandlingen av en stereoisomer av ett substrat. Således uppvisar enzymer involverade i omvandlingen av monosackarider specificitet med avseende på deras D-stereoisomerer, och enzymer involverade i omvandlingen av aminosyror - till deras L-stereo-isomerer.
Enzymaktivitet
Det speciella med enzymer som katalysatorer är att de kan ändra sina katalytiska egenskaper under påverkan av olika yttre faktorer. Ett mått på styrkan hos enzymers katalytiska verkan är deras aktivitet. Enzymernas förmåga att förändra sin aktivitet under olika förhållanden är mycket biologiskt meningsfull. Denna egenskap gör det möjligt för en levande cell att anpassa tillståndet för metaboliska processer till cellernas omedelbara behov, vilket kan förändras avsevärt under påverkan av olika yttre faktorer.
Bestämning av enzymaktivitet spelar en viktig roll i deras karakterisering. Det finns några allmänna principer för att kvantifiera enzymaktivitet. Enzymaktivitet kan bestämmas enligt följande:
· antingen genom ackumuleringshastigheten i reaktionsblandningen där enzymet i reaktionsprodukten finns;
· eller genom hastigheten för försvinnandet av substratet för den enzymatiska reaktionen från reaktionsblandningen.
Båda dessa metoder är likvärdiga och kan användas i praktiken. Vid bestämning av enzymaktivitet måste dock följande förhållanden iakttas: i reaktionsblandningen i vilken enzymaktiviteten bestäms,
· temperaturen måste motsvara enzymets temperaturoptimum;
· mediets pH måste motsvara pH-optimum för detta enzym;
· koncentrationen av substratet får inte vara mindre än den mättande;
· kofaktorer måste vara närvarande om detta enzym har några;
Enzymaktivatorer måste finnas.
Således bestäms enzymets aktivitet under optimala förhållanden. Under dessa förhållanden är enzymets aktivitet proportionell mot dess innehåll i testprovet och kan därför användas för att indirekt uppskatta dess koncentration.
Enzymaktivitet uttrycks kvantitativt i verksamhetsenheter. En enhet av enzymaktivitet (U) är den enzymaktivitet vid vilken, under dess inflytande, 1 µmol av reaktionsprodukten bildas (eller 1 µmol av substratet försvinner) per minut. I SI-systemet är enheten för enzymatisk aktivitet katal (kat). 1 katal motsvarar enzymaktivitet där en mol reaktionsprodukt bildas (en mol substrat försvinner) per sekund.
Det specifika aktivitetsvärdet används också för att karakterisera enzymer. Denna enhet återspeglar enzymets aktivitet per massenhet och uttrycks i µmol/min mg protein. Specifika aktivitetsenheter används för att bedöma renheten hos enzympreparat. Ju högre specifik aktivitet, desto renare enzympreparat.
Enzymkinetik studerar inverkan av olika faktorer (S- och E-koncentrationer, pH, temperatur, tryck, inhibitorer och aktivatorer) på hastigheten för enzymatiska reaktioner. Huvudmålet med att studera kinetiken för enzymatiska reaktioner är att få information som möjliggör en djupare förståelse av enzymers verkningsmekanism.
Kinetisk kurva låter dig bestämma den initiala reaktionshastigheten V 0 .
Substratmättnadskurva.
Beroende av reaktionshastighet på enzymkoncentration.
Beroende av reaktionshastighet på temperatur.
Reaktionshastighetens beroende av pH.
|
|
Det optimala pH-värdet för verkan av de flesta enzymer ligger inom det fysiologiska området 6,0-8,0. Pepsin är aktivt vid pH 1,5-2,0, vilket motsvarar surheten i magsaften. Arginas, ett leverspecifikt enzym, är aktivt vid 10,0. Inverkan av pH på hastigheten för en enzymatisk reaktion är associerad med tillståndet och graden av jonisering av jonogena grupper i enzymet och substratmolekylerna. Denna faktor bestämmer konformationen av proteinet, tillståndet för det aktiva centret och substratet, bildandet av enzym-substratkomplexet och själva katalysprocessen. |

Matematisk beskrivning av substratmättnadskurvan, Michaelis konstant .
|
|
Ekvationen som beskriver substratmättnadskurvan föreslogs av Michaelis och Menton och bär deras namn (Michaelis-Mentens ekvation): V = (V MAX *[ S])/(Km+[ S]) , där Km är Michaelis-konstanten. Det är lätt att räkna ut att när V = V MAX /2 Km = [S], d.v.s. Km är substratkoncentrationen vid vilken reaktionshastigheten är ½ V MAX. För att förenkla bestämningen av V MAX och Km kan Michaelis-Mentens ekvation räknas om. 1/V = (Km+[S])/(V MAX *[S]), 1/V = Km/(V MAX *[S]) + 1/V MAX , |
|
|
1/ V = Km/ V MAX *1/[ S] + 1/ V MAX Lineweaver-Burks ekvation. Ekvationen som beskriver Lineweaver-Burk-diagrammet är ekvationen för en rät linje (y = mx + c), där 1/V MAX är skärningen av den räta linjen på y-axeln; Km/V MAX - tangent för den räta linjen; skärningen av den räta linjen med abskissaxeln ger värdet 1/Km. Lineweaver-Burk-plotten låter dig bestämma Km från ett relativt litet antal punkter. Denna graf används också vid bedömning av effekten av inhibitorer, vilket kommer att diskuteras nedan. Km-värdet varierar kraftigt: från 10 -6 mol/l för mycket aktiva enzymer, till 10 -2 för lågaktiva enzymer. |


Km-uppskattningar har praktiskt värde. Vid substratkoncentrationer som är 100 gånger större än Km, kommer enzymet att arbeta med nästan maximal hastighet, så den maximala hastigheten V MAX kommer att återspegla mängden aktivt enzym som finns närvarande. Denna omständighet används för att uppskatta enzymhalten i beredningen. Dessutom är Km en egenskap hos ett enzym som används för att diagnostisera enzymopatier.
Hämning av enzymaktivitet.
En extremt karakteristisk och viktig egenskap hos enzymer är deras inaktivering under påverkan av vissa inhibitorer.
Inhibitorer - dessa är ämnen som orsakar partiell eller fullständig hämning av reaktioner som katalyseras av enzymer.
Hämning av enzymatisk aktivitet kan vara irreversibel eller reversibel, konkurrenskraftig eller icke-konkurrerande.
Irreversibel hämning - detta är ihållande inaktivering av enzymet, som är resultatet av kovalent bindning av en inhibitormolekyl på det aktiva stället eller i ett annat speciellt centrum som förändrar enzymets konformation. Dissociationen av sådana stabila komplex med regenereringen av det fria enzymet är praktiskt taget uteslutet. För att övervinna konsekvenserna av en sådan hämning måste kroppen syntetisera nya enzymmolekyler.
Reversibel hämning – kännetecknad av jämviktskomplexbildning av inhibitorn med enzymet på grund av icke-kovalenta bindningar, som ett resultat av vilket sådana komplex kan dissocieras med återställande av enzymaktivitet.
Klassificeringen av inhibitorer i konkurrenskraftiga och icke-konkurrenskraftiga baseras på om den är försvagad ( konkurrenshämning ) eller inte försvagad ( icke-konkurrensmässig hämning ) deras hämmande effekt när substratkoncentrationen ökar.
Konkurrenskraftiga hämmare - dessa är som regel föreningar vars struktur liknar strukturen hos substratet. Detta gör att de kan binda på samma aktiva ställe som substraten, vilket förhindrar enzymet från att interagera med substratet redan vid bindningsstadiet. Efter bindning kan inhibitorn omvandlas till en produkt eller förbli på det aktiva stället tills dissociation inträffar.
Reversibel kompetitiv hämning kan representeras som ett diagram:
E↔ E-I → E + P 1
S (inaktiv)
Graden av enzyminhibering bestäms av förhållandet mellan substrat- och enzymkoncentrationer.
Ett klassiskt exempel på denna typ av hämning är hämningen av succinatdehydrogenas (SDH) aktivitet av malat, som tränger undan succinat från substratstället och förhindrar dess omvandling till fumarat:

Kovalent bindning av inhibitorn till det aktiva stället resulterar i inaktivering av enzymet (irreversibel hämning). Exempel irreversibel konkurrenshämning kan fungera som inaktivering av triosefosfatisomeras med 3-kloracetolfosfat. Denna inhibitor är en strukturell analog av substratet, dihydroxiacetonfosfat, och binder irreversibelt till glutaminsyraresten på det aktiva stället:
Vissa inhibitorer verkar mindre selektivt och interagerar med en specifik funktionell grupp på det aktiva stället för olika enzymer. Således leder bindningen av jodacetat eller dess amid till SH-gruppen i aminosyran cystein, belägen i enzymets aktiva centrum och deltar i katalys, till en fullständig förlust av enzymaktivitet:
R-SH + JCH2COOH → HJ + R-S-CH2COOH
Därför inaktiverar dessa hämmare alla enzymer som har SH-grupper involverade i katalys.
Irreversibel hämning av hydrolaser under inverkan av nervgaser (sarin, soman) beror på deras kovalenta bindning till serinresten i det aktiva centret.
Den kompetitiva inhiberingsmetoden har fått bred tillämpning i medicinsk praxis. Sulfonamidläkemedel, p-aminobensoesyraantagonister, kan fungera som ett exempel på metaboliserade kompetitiva inhibitorer. De binder till dihydropteratsyntetas, ett bakteriellt enzym som omvandlar p-aminobensoat till folsyra, nödvändigt för bakterietillväxt. Bakterien dör till följd av att den bundna sulfanilamiden omvandlas till en annan förening och att folsyra inte bildas.
Icke-konkurrenskraftiga hämmare binder vanligtvis till enzymmolekylen på ett annat ställe än substratets bindningsställe, och substratet konkurrerar inte direkt med inhibitorn. Eftersom inhibitorn och substratet binder till olika centra är bildningen av både E-I-komplexet och S-E-I-komplexet möjlig. S-E-I-komplexet bryts också ner för att bilda en produkt, men med en långsammare hastighet än E-S, så reaktionen kommer att sakta ner men inte stoppa. Följande parallella reaktioner kan alltså inträffa:
E↔ E-I ↔ S-E-I → E-I + P
Reversibel icke-kompetitiv hämning är relativt sällsynt.
Icke-konkurrerande hämmare kallas allosterisk till skillnad från konkurrenskraftiga ( isosterisk ).
Reversibel hämning kan studeras kvantitativt med hjälp av Michaelis-Menten-ekvationen.
Med kompetitiv hämning förblir V MAX konstant och Km ökar.
|
|
|


Med icke-konkurrensmässig hämning minskar V MAX medan Km förblir oförändrad.
|
|
|


Om en reaktionsprodukt hämmar enzymet som katalyserar dess bildning kallas denna hämningsmetod retroinhibering eller återkopplingshämning . Till exempel hämmar glukos glukos-6-fosfatas, vilket katalyserar hydrolysen av glukos-6-fosfat.
Den biologiska betydelsen av denna hämning är regleringen av vissa metabola vägar (se nästa lektion).
PRAKTISK DEL
Uppgift för studenter
1. Studera denatureringen av proteiner under inverkan av lösningar av mineraliska och organiska syror och vid upphettning.
2. Detektera koenzym NAD i jäst.
3. Bestäm amylasaktivitet i urin (blodserum).
9. STANDARDER FÖR SVAR PÅ PROBLEM, testfrågor som används för att kontrollera kunskaper i klassen (kan användas som bilaga)
10. ART OCH OMFATTNING AV EVENTUELLT UTBILDNINGS- OCH FORSKNINGSARBETE PÅ ÄMNET
(Ange specifikt arten och formen av UIRS: förbereda abstrakta presentationer, utföra oberoende forskning, simuleringsspel, fylla i en medicinsk historia med hjälp av monografisk litteratur och andra former)
Enzymkinetik studerar hastigheten för reaktioner som katalyseras av enzymer beroende på olika förhållanden (koncentration, temperatur, pH, etc.) för deras interaktion med substratet.
Emellertid är enzymer proteiner som är känsliga för påverkan av olika yttre påverkan. Därför, när de studerar hastigheten för enzymatiska reaktioner, tar de främst hänsyn till koncentrationerna av reagerande ämnen och försöker minimera påverkan av temperatur, pH i miljön, aktivatorer, inhibitorer och andra faktorer och skapa standardförhållanden. För det första är detta pH-värdet i miljön som är optimalt för ett givet enzym. För det andra rekommenderas att hålla en temperatur på 25°C, där så är möjligt. För det tredje uppnås fullständig mättnad av enzymet med substratet. Denna punkt är särskilt viktig eftersom vid låga substratkoncentrationer deltar inte alla enzymmolekyler i reaktionen (Fig. 6.5, A), vilket innebär att resultatet blir långt ifrån det maximala möjliga. Den största kraften i den katalyserade reaktionen, allt annat lika, uppnås om varje enzymmolekyl deltar i omvandlingen, d.v.s. vid en hög koncentration av enzym-substratkomplexet (Fig. 6.5, V). Om substratkoncentrationen inte säkerställer fullständig mättnad av enzymet (Fig. 6.5, b), så når reaktionshastigheten inte sitt maximala värde.
Ris. 65.
A - vid låg substratkoncentration; 6 - med otillräcklig substratkoncentration; V - när enzymet är helt mättat med substrat
Hastigheten för en enzymatisk reaktion mätt under ovanstående förhållanden och fullständig mättnad av enzymet med substratet kallas maximal hastighet för enzymatisk reaktion (V).
Hastigheten för den enzymatiska reaktionen, bestäms när enzymet inte är helt mättat med substratet, anges v.
Enzymkatalys kan förenklas med följande diagram:

där F är ett enzym; S - substrat; FS - enzym-substratkomplex.
Varje steg i denna process kännetecknas av en viss hastighet. Måttenheten för hastigheten för en enzymatisk reaktion är antalet mol substrat som omvandlas per tidsenhet(samma som hastigheten för en normal reaktion).
Enzymets interaktion med substratet leder till bildandet av ett enzym-substratkomplex, men denna process är reversibel. Hastigheterna för framåt- och bakåtreaktioner beror på koncentrationerna av reaktanterna och beskrivs med motsvarande ekvationer:
I jämvikt är ekvation (6.3) giltig, eftersom hastigheterna för framåt- och bakåtreaktionerna är lika.
Genom att ersätta hastighetsvärdena för framåt (6.1) och bakåt (6.2) reaktioner med ekvation (6.3), får vi likheten:
Jämviktstillståndet kännetecknas av en lämplig jämviktskonstant K p, lika med förhållandet mellan konstanterna för framåt- och bakåtreaktionen (6.5). Det reciproka av jämviktskonstanten kallas substratkonstant Ks, eller dissociationskonstanten för enzym-substratkomplexet:

Av ekvation (6.6) framgår att substratkonstanten minskar vid höga koncentrationer av enzym-substratkomplexet, d.v.s. med stor stabilitet. Följaktligen karaktäriserar substratkonstanten affiniteten hos enzymet och substratet och förhållandet mellan hastighetskonstanterna för bildning och dissociation av enzym-substratkomplexet.
Fenomenet enzymmättnad med substrat studerades av Leonor Michaelis och Maud Mepten. Baserat på matematisk bearbetning av resultaten härledde de ekvationen (6.7), som fick sina namn, från vilken det är tydligt att vid en hög substratkoncentration och ett lågt värde på substratkonstanten tenderar hastigheten för den enzymatiska reaktionen till det maximala . Denna ekvation är dock begränsad eftersom den inte tar hänsyn till alla parametrar:
Enzym-substratkomplexet under reaktionen kan genomgå transformationer i olika riktningar:
- dissociera till modersubstanser;
- omvandlas till en produkt från vilken enzymet separeras oförändrat.
Därför, för att beskriva den övergripande verkan av den enzymatiska processen, konceptet Michaelis konstanter Kt, som uttrycker förhållandet mellan hastighetskonstanterna för alla tre reaktionerna av enzymatisk katalys (6.8). Om båda termerna divideras med reaktionshastighetskonstanten för bildandet av enzym-substratkomplexet får vi uttryck (6.9):

En viktig följd följer av ekvation (6.9): Michaelis-konstanten är alltid större än substratkonstanten med mängden k 2 /k v
Numeriskt K t lika med koncentrationen av substratet vid vilken reaktionshastigheten är hälften av den maximala möjliga hastigheten och motsvarar mättnaden av enzymet med substratet, som i fig. 6,5, b. Eftersom det i praktiken inte alltid är möjligt att uppnå fullständig mättnad av enzymet med substratet, är det just K t används för jämförande karakterisering av enzymers kinetiska egenskaper.
Hastigheten för den enzymatiska reaktionen när enzymet inte är helt mättat med substratet (6.10) beror på koncentrationen av enzym-substratkomplexet. Proportionalitetskoefficienten är reaktionskonstanten för frisättningen av enzymet och produkten, eftersom detta ändrar koncentrationen av enzym-substratkomplexet:
Efter transformationer, med hänsyn till ovanstående beroenden, beskrivs hastigheten för den enzymatiska reaktionen när enzymet inte är helt mättat med substratet med ekvation (6.11), dvs. beror på koncentrationerna av enzymet, substratet och deras affinitet K s:
Det grafiska beroendet av hastigheten för en enzymatisk reaktion på koncentrationen av substratet är inte linjärt. Som framgår av fig. 6.6, med ökande substratkoncentration observeras en ökning i enzymaktivitet. Men när maximal mättnad av enzymet med substratet uppnås, blir hastigheten för den enzymatiska reaktionen maximal. Därför är den hastighetsbegränsande faktorn för reaktionen bildningen av ett enzym-substratkomplex.
Praxis har visat att substratkoncentrationer som regel uttrycks i värden mycket mindre än enhet (10 6 -10 3 mol). Det är ganska svårt att arbeta med sådana mängder i beräkningar. Därför föreslog G. Lineweaver och D. Burke att uttrycka det grafiska beroendet av hastigheten för en enzymatisk reaktion inte i direkta koordinater, utan i omvända. De utgick från antagandet att för lika storheter är deras inverser också lika:

Ris. 6.6.
Efter att ha transformerat uttryck (6.13) får vi ett uttryck som kallas Lineweaver-Burks ekvation (6.14):
Det grafiska beroendet för Lineweaver-Burk-ekvationen är linjär (Fig. 6.7). Enzymets kinetiska egenskaper bestäms enligt följande:
- segmentet avskuret på ordinataaxeln är lika med 1/V;
- segmentet avskuret på abskissaxeln är lika med -1 /Till t.

Ris. 6.7.
Man tror att Lineweaver-Burk-metoden gör det möjligt att bestämma den maximala reaktionshastigheten mer exakt än i direkta koordinater. Värdefull information om enzymhämning kan också hämtas från denna graf.
Det finns andra sätt att transformera Michaelis-Mentens ekvation. Grafiska beroenden används för att studera påverkan av olika yttre påverkan på den enzymatiska processen.
Denna gren av enzymologi studerar inverkan av olika faktorer på hastigheten för en enzymatisk reaktion. Med tanke på den allmänna ekvationen för enzymatisk katalys av den reversibla reaktionen att omvandla ett substrat till en produkt (1),
De viktigaste faktorerna som påverkar hastigheten för en enzymatisk reaktion bör nämnas: substratkoncentration [S], enzymkoncentration [E] och reaktionsproduktkoncentration [P].
Interaktionen mellan vissa enzymer och deras substrat kan beskrivas med en hyperbolisk kurva över beroendet av hastigheten för enzymatisk reaktion V på koncentrationen av substratet [S] (Fig. 19):

Fig. 19. Beroende av hastigheten för enzymatisk reaktion på koncentrationen av substratet.
På denna kurva kan tre sektioner särskiljas, vilket kan förklaras av bestämmelserna om mekanismen för interaktion mellan enzymet och substratet: OA - en sektion med direkt proportionellt beroende av V på [S], enzymets aktiva centra fylls gradvis med substratmolekyler med bildandet av ett instabilt komplex ES; avsnitt AB - kurvlinjärt beroende av V på [S], fullständig mättnad av enzymets aktiva centra med substratmolekyler har ännu inte uppnåtts. ES-komplexet är instabilt innan det når övergångstillståndet. Sannolikheten för omvänd dissociation till E och S är fortfarande hög; sektion BC - beroendet beskrivs av en nollordningens ekvation, sektionen är parallell med [S]-axeln, fullständig mättnad av aktiva enzymer med substratmolekyler har uppnåtts, V=V max.
Den karakteristiska formen på kurvan beskrivs matematiskt av Briggs-Haldane-ekvationen:
V=V max ● [S]/ Km + [S] (2),
där Km är Michaelis-Menten-konstanten, numeriskt lika med substratkoncentrationen vid vilken hastigheten för den enzymatiska reaktionen är lika med halva Vmax.
Ju lägre Km för enzymet, desto högre affinitet har enzymet för substratet, desto snabbare uppnås övergångstillståndet för substratet, och det förvandlas till en reaktionsprodukt. Att hitta Km-värden för varje gruppspecifikt enzymsubstrat är viktigt för att bestämma den biologiska rollen för detta enzym i cellen.
För de flesta enzymer är det omöjligt att konstruera en hyperbolisk kurva (Fig. 19) I detta fall används metoden med dubbla reciproka (Lineweaver-Burk), dvs. ett grafiskt beroende av 1/[V] på 1/[S] plottas (fig. 20). Metoden att konstruera sådana kurvor i ett experiment är mycket bekväm när man studerar effekten av olika typer av inhibitorer på enzymaktivitet (se vidare i texten).

Fig.20. Graf av 1/[V] kontra 1/[S] (Lineweaver-Burk-metoden),
där y är gränssnittet - och x är gränssnittet -  , tangens av vinkeln α - .
, tangens av vinkeln α - .
Beroende av hastigheten för enzymatisk reaktion V på enzymkoncentrationen [E].
Detta grafiska beroende (fig. 21) anses vid optimal temperatur och pH i miljön, vid substratkoncentrationer som är betydligt högre än mättnadskoncentrationen av enzymets aktiva centra.

Ris. 21. Inverkan av enzymkoncentration på hastigheten för enzymatisk reaktion.
Beroende av hastigheten för en enzymatisk reaktion på koncentrationen av en kofaktor eller koenzym. För komplexa enzymer bör det beaktas att en brist på koenzymformer av vitaminer vid hypovitaminos och en kränkning av intaget av metalljoner i kroppen nödvändigtvis leder till en minskning av koncentrationen av motsvarande enzymer som är nödvändiga för kursen av metaboliska processer. Därför bör man dra slutsatsen att enzymets aktivitet är direkt beroende av koncentrationen av kofaktorn eller koenzymet.
Produktkoncentrationens inverkan på hastigheten för enzymatisk reaktion. För reversibla reaktioner som inträffar i människokroppen måste man ta hänsyn till att produkterna från den direkta reaktionen kan användas av enzymet som substrat för den omvända reaktionen. Därför beror flödesriktningen och ögonblicket för att nå Vmax på förhållandet mellan koncentrationerna av de initiala substraten och reaktionsprodukterna. Till exempel aktiviteten av alaninaminotransferas, som katalyserar transformationen:
Alanin + Alfa-ketoglutarat ↔ Pyruvat + Glutamat
beror i cellen på koncentrationsförhållandet:
[alanin + alfa-ketoglutarat] / [pyruvat + glutamat].
MEKANISM FÖR ENZYMVERKNING. TEORIER OM ENZYMKATALYS
Enzymer, liksom icke-proteinkatalysatorer, ökar hastigheten för en kemisk reaktion på grund av deras förmåga att minska aktiveringsenergin för denna reaktion. Aktiveringsenergin för en enzymatisk reaktion beräknas som skillnaden mellan energivärdet i systemet för den pågående reaktionen som har nått övergångstillståndet och energin som bestäms i början av reaktionen (se grafiskt beroende i fig. 22).

Ris. 22. Grafiskt beroende av energitillståndet för en kemisk reaktion utan ett enzym (1) och i närvaro av ett enzym (2) på reaktionstiden.
V. Henrys och i synnerhet L. Michaelis, M. Mentens arbete med att studera mekanismen för monosubstrat reversibla enzymatiska reaktioner gjorde det möjligt att postulera att enzym E först reversibelt och relativt snabbt kombineras med sitt substrat S för att bilda ett enzym- substratkomplex (ES):
E+S<=>ES (1)
Bildningen av ES sker på grund av vätebindningar, elektrostatiska, hydrofoba interaktioner, i vissa fall kovalenta, koordinationsbindningar mellan sidoradikalerna av aminosyrarester i det aktiva centret och de funktionella grupperna i substratet. I komplexa enzymer kan funktionen av kontakt med substratet också utföras av den icke-proteiniska delen av strukturen.
Enzym-substratkomplexet sönderfaller sedan i en andra, långsammare, reversibel reaktion för att producera reaktionsprodukt P och fritt enzym E:
ES<=>EP<=>E+P (2)
För närvarande, tack vare de ovan nämnda forskarnas arbete, såväl som Keilin D., Chance B., Koshland D. (teorin om "inducerad korrespondens"), finns det teoretiska bestämmelser om fyra huvudpunkter i verkningsmekanismen av ett enzym på ett substrat, som bestämmer enzymers förmåga att påskynda kemiska reaktioner:
1. Orientering och förhållningssätt . Enzymet kan binda en substratmolekyl på ett sådant sätt att bindningen som angrips av enzymet inte bara är belägen i närheten av den katalytiska gruppen, utan också korrekt orienterad med avseende på den. Sannolikheten för att ES-komplexet kommer att nå övergångstillståndet genom orientering och närhet ökar kraftigt.
2. Stress och belastning : framkallad korrespondens. Fästningen av ett substrat kan orsaka konformationsförändringar i enzymmolekylen, vilket leder till spänningar i strukturen av det aktiva centret, och även något deformerar det bundna substratet, vilket underlättar uppnåendet av ett övergångstillstånd av ES-komplexet. En så kallad inducerad överensstämmelse uppstår mellan molekylerna E och S.
Hastigheten för enzymatiska reaktioner beror på koncentrationen av enzymet, substrat, temperatur, pH och närvaron av aktivatorer och inhibitorer.
Under förhållanden med överskott av substrat, reaktionshastigheten direkt proportionerlig enzymkoncentration (Fig. 3.2).
Ris. 3.2. Beroende av reaktionshastighet på enzymkoncentration.
Beroende av reaktionshastighet på substratkoncentration presenteras i figur 3.3.
Ris. 3.3. Beroende av reaktionshastighet på substratkoncentration.
Det finns 3 avsnitt på grafen. Vid låg substratkoncentration (sektion A) reaktionshastigheten är direkt proportionell mot substratkoncentrationen och följer första ordningens kinetik. Plats på b(blandad ordning reaktion) detta beroende kränks. Plats på c reaktionshastigheten är maximal och beror inte på substratkoncentrationen.
En enzymatisk reaktion kännetecknas av bildandet av ett enzym-substratkomplex, som bryts ner för att bilda det fria enzymet och reaktionsprodukten.
I denna ekvation är k 1 hastighetskonstanten för bildningen av enzym-substratkomplexet, k 2 är dissociationskonstanten för enzym-substratkomplexet för att bilda ett fritt enzym och substrat, och k 3 är hastighetskonstanten för dissociationen av enzym-substratkomplexet till det fria enzymet och reaktionsprodukten.
Michaelis och Menten föreslog en ekvation som beskriver reaktionshastighetens beroende av substratkoncentrationen.
v är reaktionshastigheten vid en given substratkoncentration; Ks – dissociationskonstant för enzym-substratkomplexet; Vmax – maximal reaktionshastighet.
Ks=k-2/ki, dvs. förhållandet mellan den omvända reaktionskonstanten och den framåtriktade reaktionskonstanten.
Denna ekvation beskriver dock endast avsnittet A på grafen och tar inte hänsyn till reaktionsprodukternas inverkan på den enzymatiska processens hastighet.
Haldane och Briggs ersatte dissociationskonstanten i ekvationen med Michaelis-konstanten (Km).
Michael är konstant numeriskt lika med substratkoncentration, vid vilken reaktionshastigheten är hälften av maximum. Michaelis-konstanten kännetecknar affiniteten hos enzymet och substratet. En hög affinitet hos ett enzym för ett substrat kännetecknas av ett lågt Km-värde och vice versa.
Att använda den graf som Michaelis och Menten föreslagit är obekvämt. För en mer bekväm grafisk representation transformerade G. Lineweaver och D. Burke Haldane och Briggs ekvation med hjälp av metoden med dubbla reciproka, baserat på principen att om det finns likhet mellan två kvantiteter, så kommer de reciproka också att vara lika.
Grafisk representation av reaktionshastighetens beroende av pH har en klockform. Det pH-värde vid vilket enzymet uppvisar maximal aktivitet kallas optimalt pH(Fig. 5.4 A) . För de flesta enzymer är det optimala pH 6-8. Undantaget är pepsin, vars optimum är 2,0. När pH ändras i en eller annan riktning från det optimala, minskar reaktionshastigheten på grund av joniseringen av de funktionella grupperna i enzymet och substratet, vilket stör bildningen av enzym-substratkomplexet.
Ris. 3.4. Reaktionshastighetens beroende av pH (A) och temperatur (B).
Hastigheten för en kemisk reaktion ökar med 2 gånger med ökningen temperatur med 10°C. Men på grund av enzymets proteinnatur, med en ytterligare ökning av temperaturen, sker denaturering av enzymet. Den temperatur vid vilken reaktionshastigheten är maximal kallas temperaturoptimal(Fig. 3.4. B) . För de flesta enzymer är den optimala temperaturen 37-40°C. Undantaget är muskelmyokinas, som tål uppvärmning upp till 100°C.
Enzymaktivatorer– dessa är ämnen 1) som bildar enzymets aktiva centrum (Co 2+, Mg 2+, Zn 2+, Fe 2+, Ca 2+); 2) underlätta bildningen av enzym-substratkomplexet (Mg2+); 3) reducering av SH-grupper (glutation, cystein, merkaptoetanol); 4) stabilisering av den naturliga strukturen hos proteinenzymet. Enzymatiska reaktioner aktiveras vanligtvis av katjoner (i det periodiska systemet från 19 till 30). Anjoner är mindre aktiva, även om klorjoner och anjoner av vissa andra halogener kan aktivera pepsin, amylas och adenylatcyklas. Proteiner kan vara aktivatorer: apoprotein A-I (LCAT), apoprotein C-II (LPL).
Verkningsmekanism för aktivatorer:
1) delta i bildandet av enzymernas aktiva centrum;
2) underlätta bindningen av substratet och enzymet;
3) delta i bildandet av enzymets naturliga struktur.
Inhibitorer– ämnen som orsakar partiell eller fullständig hämning av reaktioner som katalyseras av enzymer.
Inhibitorer klassificeras i ospecifik Och specifik. Verkan av ospecifika inhibitorer är inte relaterad till enzymers verkningsmekanism. Dessa inhibitorer orsakar denaturering av enzymproteinet (värme, syror, alkalier, salter av tungmetaller, etc.).
Specifika inhibitorer påverkar verkningsmekanismen för enzymer. Specifika inhibitorer är indelade i 2 grupper: reversibel och irreversibel. Irreversibla inhibitorer orsakar en permanent, irreversibel förändring eller modifiering av enzymets funktionella grupper genom tät eller kovalent bindning. Denna grupp inkluderar: 1) metallhämmare enzymer (HCN, RCN, HF, CO, etc.). Dessa föreningar binder till metaller med variabel valens (Cu eller Fe), som ett resultat av vilket processen för elektronöverföring längs andningskedjan av enzymer störs. Därför kallas dessa hämmare luftvägsgifter. 2) hämmare av enzymer innehållande SH-grupper(monoidoacetat, dijodacetat, jodoacetamid, arsenik och kvicksilverföreningar). 3) hämmare av enzymer som innehåller en OH-grupp i det aktiva centret (organofosforföreningar, insekticider). Dessa hämmare hämmar först och främst aktiviteten av kolinesteras, ett enzym som spelar en primär roll i nervsystemets aktivitet.
Reversibel hämning kan kvantifieras med användning av Michaelis-Menten-ekvationen. Reversibla inhibitorer är indelade i konkurrenskraftiga och icke-konkurrenskraftiga.
Konkurrenskraftiga hämmare– Det är ämnen som till strukturen liknar substratet. Inhibitorn binder till enzymets aktiva ställe och förhindrar bildningen av enzym-substratkomplexet.
Ett klassiskt exempel på kompetitiv hämning är hämningen av succinatdehydrogenas av malonsyra. Succinatdehydrogenas katalyserar oxidationen av bärnstenssyra (succinat) genom dehydrering till fumarsyra.
Om malonsyra (en inhibitor) tillsätts till mediet, kommer den, som ett resultat av dess strukturella likhet med det verkliga substratet succinat, att reagera med det aktiva stället för att bilda ett enzym-inhibitorkomplex, men reaktionen kommer inte att inträffa.
Effekten av inhibitorn elimineras av ökar substratkoncentrationen. Med kompetitiv hämning förändras kinetiken för enzymatiska reaktioner: Km ökar, V max förblir konstant(Fig. 3.5).
Ris. 3.5. Effekt av kompetitiva inhibitorer på hastigheten för enzymatisk reaktion
Den kompetitiva inhiberingsmetoden har funnit tillämpning i medicinsk praxis som antimetaboliter.
Till exempel används sulfonamidläkemedel för att behandla vissa infektionssjukdomar orsakade av bakterier. Dessa läkemedel liknar strukturellt para-aminobensoesyra, som bakteriecellen använder för att syntetisera folsyra, vilket är nödvändigt för bakteriers liv. På grund av denna strukturella likhet blockerar sulfonamid verkan av enzymet genom att ersätta para-aminobensoesyra från komplexet med enzymet som syntetiserar folsyra.
Icke-konkurrerande hämmare –ämnen som inte strukturellt liknar substrat. Icke-kompetitiva inhibitorer binder inte till det aktiva stället, utan till en annan plats i enzymmolekylen, till exempel i det allosteriska centret. Detta ändrar konformationen av det aktiva centret på ett sådant sätt att interaktionen mellan substratet och det störs.
För icke-konkurrensmässig hämning: V max minskar, men K m ändras inte(Fig. 3.6).