Бактериялар патшалығының тірі ағзаларының биосферадағы рөлі. Аннотация: Прокариоттардың күкірт айналымына қатысуы Сабақтың тіректері
Сұрақ 1. Прокариоттардың биосферадағы рөлін сипаттаңыз.
Прокариоттар фотосинтез жүргізеді, осылайша атмосфераға оттегін шығарады. Хемосинтез прокариоттар арасында кең таралған. Сонымен қатар, бактериялық организмдер арасында азотты бекітетін формалар бар: бұл атмосфералық ауадан азотты тікелей сіңіруге қабілетті және осылайша биологиялық айналымға молекулалық азотты тарта алатын біздің планетамыздағы тірі ағзалардың жалғыз тобы.
Прокариоттардың тағы бір маңызды қызметі бар: органикалық қосылыстардың жойылуы (минералдануы) арқылы бейорганикалық заттарды қоршаған ортаға қайтару.
Жалпы биосфера деңгейінде прокариоттардың, ең алдымен, бактериялардың тағы бір өте маңызды функциясы бар - концентрация. Олар қоршаған ортадан кейбір элементтерді өте төмен концентрацияларда да белсенді түрде бөліп алуға қабілетті.
Прокариоттардың қасиеттері мен функцияларының әртүрлілігі сонша, олар негізінен тұрақты жұмыс істейтін экожүйелерді құруға қабілетті (яғни тек олардың қатысуымен).
2-сұрақ. Табиғи ортаға оның ластаушыларымен күресу үшін прокариоттық препараттарды енгізудің қандай қауіпті екенін түсіндіріңіз.
Прокариоттарды қоршаған ортаға енгізу кезінде туындайтын проблемалық мәселелердің бірі жергілікті микрофлора мен енгізілген микрофлораның арасындағы қарым-қатынастың сипатын анықтау болып табылады. Микробиоценоздардағы қалыптасқан тепе-теңдіктің өзгеруі нәтижесінде енгізілген микроорганизмдердің табиғи биогеоценоздардың күйіне кері әсер ету мүмкіндігін жоққа шығаруға болмайды. Бұл жағдай аса сақтықты талап етеді және мұқият зерттеуден кейін ғана прокариоттарды адам қажеттіліктері үшін және биосфераны ластаушы заттармен күресу үшін пайдалану керек.
3-сұрақ. Азотфиксациялаушы бактериялардың барлығында молекулалық азоттың байланысуын катализдейтін нитрогеназа ферменті бірдей болатыны белгілі. Бұл фактінің тірі табиғат эволюциясында қандай маңызы бар?
Нитрогеназа – екі ақуыздың кешенінен тұратын көп өлшемді фермент. Әртүрлі азот бекіткіштерден алынған азотогеназалар олардың молекулалық салмағы мен металдық құрамы бойынша біршама ерекшеленеді. Бұл факт азотты түзетін бактериялардың шығу тегі бірлігін көрсетеді. Нитрогеназа бактериялардың тектік түрінде пайда болды, содан кейін дивергенция пайда болды, бұл азотты бекітетін бактериялардың әртүрлілігіне әкелді, олардың әрқайсысында нитрогеназаны кодтайтын ген болды.
Азотты бекіту процесі қалыпты атмосфералық қысымда және қалыпты температурада жүреді, бірақ көп энергияны қажет етеді.
Нитрогеназаның белсенді жұмыс істеуі үшін микроаэрофильді жағдайлар қажет. Молекулалық оттегі екі нитрогеназа протеиніне де зиянды әсер етеді. Түйіндегі микроаэрофильді жағдайлар оттегіге диффузды тосқауылмен қамтамасыз етіледі. Ауадан аммиак өндіретін нитрогеназа кешені өте үнемді әрекет етеді. Егер мекендейтін ортада аммоний немесе нитрат иондары жеткілікті болса, ол жұмысын тоқтатады.
Азот барлық тірі организмдер үшін өте маңызды элемент болып табылады. Азоттың негізгі қоры – жер атмосферасы. Эукариотты организмдер атмосферадан тікелей азотты сіңіре алмайды. Молекулярлық азоттың байланысуын катализдейтін нитрогеназа ферментінің азотты түзетін бактерияларда болуына байланысты олармен симбиозда өмір сүретін өсімдіктер өздеріне оңай жететін азотты ала алады. Көптеген өсімдіктер, жануарлар және саңырауқұлақтар азотты түзетін прокариоттармен симбиозға түсу қабілетіне ие.
4-сұрақ. 70-жылдардың басында американдық биолог және эколог Барри Коммонер. ХХ ғасыр Экология заңы ретінде «Табиғат жақсы біледі» деген диссертацияны алға тартты. Не айтқыңыз келгенін түсіндіріңіз. Осы мәселе бойынша өз пікіріңізді білдіріңіз. Жауабыңызды мысалдар арқылы түсіндіріңіз.
Барри Коммонер табиғаттың механизмдері мен функциялары туралы толық сенімді ақпарат болғанша, біз сағаттың құрылымымен таныс емес, бірақ оны түзеткіміз келетін адам сияқты, оларды жақсартуға тырысып, табиғи жүйелерге оңай зиян келтіреміз дейді. Ол өте сақ болуға шақырады. Табиғаттың өзгеруі экономикалық апатты және экологиялық қауіпті. Сайып келгенде, өмір сүруге жарамсыз жағдайлар жасалуы мүмкін. Жақсартудың экологиялық критерийін көрсетпей, табиғатты жақсарту туралы қалыптасқан пікір мағынасыз.
Мен бұл пікірмен келісемін. Табиғаттың бір көрінісі биоценоздар, биогеоценоздар немесе жалпы биосфера үшін одан да жойқын және зиянды құбылыстың салдары деп жиі ойламаймыз. «Ең бастысы, біз өзімізді жақсы сезінеміз» және табиғат күтеді. Олай болмайды!!!
«Орман тәртіпшілері» болып шыққан қасқырларды ату мысалдарын келтіруге болады. Қытайда торғайлар жойылды, олар егінді жояды, бірақ құстарсыз егінді зиянды жәндіктер құртады деп ешкім ойлаған жоқ.
Бұл эукариоттық жасушаның пайда болуы биологиялық эволюциядағы ең маңызды оқиғалардың бірі болып табылады. Эукариот пен прокариоттық организмдердің айырмашылығы геномдық реттеудің жетілдірілген жүйесі болып табылады. Ал осының арқасында бір жасушалы организмдердің бейімделгіштігі, олардың геномға тұқым қуалаушылық өзгерістерін енгізбей өзгермелі орта жағдайларына бейімделу қабілеті артты. Бейімделу қабілетінің арқасында эукариоттар көп жасушалы бола алды – көп жасушалы организмде геномы бірдей жасушалар жағдайларға байланысты морфологиясы мен қызметі жағынан мүлде басқа ұлпалар түзеді.
Бұл ароморфоз архей мен протерозойдың (2,6 - 2,7 млрд жыл бұрын) тоғысында орын алды, оны биомаркерлер – тек эукариоттық жасушаларға тән стероидты қосылыстардың қалдықтары анықтады. Эукариоттардың пайда болуы оттегі революциясымен сәйкес келеді.
Эукариоттардың прокариоттардың бірнеше сорттарының симбиозы нәтижесінде пайда болғаны жалпы қабылданған. Шамасы, митохондриялар альфа-протеобактериялардан (аэробты эубактериялар), пластидтер цианобактериялардан және цитоплазмадан белгісіз архебактериялардан пайда болған. Ядроның, цитоскелеттің және жгутиканың шығу тегі туралы жалпы қабылданған теория әлі жоқ. Жердегі тіршіліктің пайда болуы туралы гипотезалар жасушаның пайда болуы туралы мәселені нақтылай алмады. Егер прокариоттардың пайда болуы туралы олардың пайда болуын дәлелді түрде сипаттайтын гипотезалар іс жүзінде болмаса, эукариот жасушаларының шығу тегі туралы бірнеше көзқарастар бар.
Эукариоттардың пайда болуының негізгі гипотезалары:
1. Симбиотикалық гипотезаекі тұжырымдамаға негізделген. Осы концепциялардың біріншісі бойынша тірі табиғаттағы ең іргелі ерекшелік бактериялар мен нағыз ядролары бар жасушалардан тұратын организмдер – протистерден, жануарлардан, саңырауқұлақтардан және өсімдіктерден тұрады. Екінші концепция эукариоттық жасушалардың кейбір бөліктерінің шығу тегі симбиоздардың эволюциясы - әртүрлі түрдегі организмдер арасында тұрақты ассоциациялардың пайда болуы. Органеллалардың үш класы - митохондриялар, кірпікшелер және фотосинтездеуші пластидтер симбиоз нәтижесінде прокариот иесі жасушаларына енген еркін тіршілік ететін бактериялардан пайда болды деп болжанады. Бұл теория негізінен Мендельдік генетиканы Дарвиннің табиғи сұрыпталу идеясымен байланыстырған генетиктер, экологтар және цитологтар әзірлеген неодарвиндік идеяларға сүйенеді. Ол сондай-ақ молекулалық биологияға, әсіресе белоктар мен аминқышқылдарының тізбегінің құрылымы туралы мәліметтерге, жер бетіндегі тіршіліктің ең алғашқы іздерін зерттейтін микропалеонтологияға және бұл ғылымдар биологиялық текті газдармен айналысатындықтан, атмосфералық физика мен химияға сүйенеді.
2.Инвагинация гипотезасыэукариоттық жасушаның тектік формасы аэробты прокариот болғанын айтады. Ішінде жасуша мембранасына бекітілген бірнеше геномдар болды. Корпускулярлы органоидтар мен ядро қабықшаның бөлімдерінің инвагинациялануынан және байлануынан пайда болды, содан кейін ядроға, митохондрияларға және хлоропластарға функционалды мамандану. Содан кейін эволюция процесінде ядролық геном күрделеніп, цитоплазмалық мембраналар жүйесі пайда болды. Бұл гипотеза қабықшаларда ядроның, митохондриялардың, хлоропласттардың және екі мембрананың болуын түсіндіреді. Бірақ ол корпускулярлық органеллалардағы ақуыз биосинтезі процесінің бөлшектеріндегі және эукариоттық жасушаның цитоплазмасындағы айырмашылықтарды түсіндіруде қиындықтарға тап болады. Митохондриялар мен хлоропласттарда бұл процесс қазіргі прокариот жасушаларындағы процесске дәл сәйкес келеді.
Симбиотикалық (I) және инвагинация (II) гипотезалары бойынша эукариоттық жасушаның шығу тегі:
1 - анаэробты прокариот (ие жасушасы), 2 - митохондриялары бар прокариоттар, 3 - көк-жасыл балдырлар (болжамды хлоропласт), 4 - сиирохета тәрізді бактерия (болжамды жілік), 5 - жілік жасушасы бар қарабайыр эукариот, 6 - өсімдік. 7 - жіліншегі бар жануар жасушасы, 8 - аэробты прокариот (болжамдық митохондрия), 9 - аэробты прокариот (II гипотеза бойынша тектік жасуша), 10 - ядро мен митохондрияны тудырған жасуша мембранасының инвагинациялары, 11 - примитивтік. эукариот 12 - хлоропласт берген жасуша қабықшасының инвагинациясы, 13 - өсімдік жасушасы; а - прокариот жасушасының ДНҚ, б - митохондрия, в - эукариот жасушасының ядросы, d - жілік, е - хлоропласт.
Қолда бар деректер гипотезалардың біріне артықшылық беруге немесе ғалымдардың көпшілігіне сәйкес келетін жаңасын әзірлеуге әлі жеткіліксіз, бірақ соңғы жылдары эукариоттық жасушаның пайда болуының симбиогенетикалық теориясын сенімді түрде дәлелдеу мүмкін болды.
Эукариот жасушаларының эволюциялық мүмкіндіктері прокариоттық жасушаларға қарағанда жоғары. Мұнда жетекші рөл прокариоттардың геномынан үлкенірек эукариоттардың ядролық геномына жатады. Маңызды айырмашылықтар - ядроларда гендердің екі жиынтығының болуына байланысты эукариоттық жасушалардың диплоидтылығы, сонымен қатар кейбір гендердің бірнеше рет қайталануы.
Жасуша белсенділігін реттеу механизмі күрделене түсуде, ол реттеуші гендердің салыстырмалы санының көбеюінен және прокариоттардың дөңгелек «жалаңаш» ДНҚ молекулаларының ДНҚ ақуыздармен байланысқан хромосомалармен алмастырылуынан көрінеді.
Аэробты тыныс алу да көп жасушалы формалардың дамуының алғы шарты болды. Эукариоттық жасушалардың өздері Жерде атмосферадағы O 2 концентрациясы 1% (Пастер нүктесі) жеткеннен кейін пайда болды. Ал бұл концентрация аэробты тыныс алудың қажетті шарты болып табылады.
Әрбір эукариоттық жасушада әртүрлі шығу тегі бар геномдар болатыны белгілі: жануарлар мен саңырауқұлақ жасушаларында бұл ядро мен митохондриялардың геномдары, ал өсімдік жасушаларында бұл да пластидті геномдар. Кішкентай дөңгелек ДНҚ эукариот жасушаларының жгутикасының базальды денесінде де болады.
Молекулалық сағат әдісі бойынша эукариоттар прокариоттармен бір мезгілде пайда болды. Бірақ Жер тарихының маңызды бөлігінде прокариоттардың басым болғаны анық. Эукариоттық өлшемдерге сәйкес келетін алғашқы жасушалар (акритархтар) 3 миллиард жыл болды, бірақ олардың табиғаты әлі анық емес. Эукариоттардың белгілі дерлік қалдықтарының жасы шамамен 2 миллиард жыл. Ал оттегі төңкерісінен кейін ғана планетаның бетінде эукариоттар үшін қолайлы жағдайлар дамыды (шамамен 1 млрд жыл бұрын).
Сірә, эукариоттық жасушалардың негізгі атасы архебактериялар болды, олар тамақ бөлшектерін жұту арқылы қоректенуге көшті. Мұндай жұту үшін қажетті жасуша пішінінің өзгеруі актин мен миозиннен тұратын цитоскелет арқылы қамтамасыз етілді. Мұндай жасушаның тұқым қуалайтын аппараты мембранамен байланысын сақтай отырып, оның өзгеретін бетінен тереңірек жылжыды. Бұл ядролық кеуектері бар ядролық қабықтың пайда болуына себеп болды.
Ие жасушасы сіңірген бактериялар оның ішінде өмір сүруін жалғастыра алады. Осылайша, митохондриялардың ата-бабалары фотосинтетикалық бактериялар тобына айналды - күлгін альфапротеобактериялар. Ие жасушасының ішінде олар фотосинтездеу қабілетін жоғалтып, органикалық заттардың тотығуын өз мойнына алды. Олардың арқасында эукариоттық жасушалар аэробты болды. Басқа фотосинтетикалық жасушалармен симбиоздар өсімдік жасушаларының пластидтерге ие болуына себеп болды. Мүмкін эукариоттық жасушалардың флагеллалары қозғалуға қабілетті жасушалар мен бактериялар арасындағы симбиоздың нәтижесінде пайда болуы мүмкін.
Эукариот жасушаларының тұқым қуалайтын аппараты шамамен прокариоттардың құрылымына ұқсас болды. Бірақ үлкенірек және күрделірек жасушаны басқару қажеттілігіне байланысты хромосомалардың ұйымы кейінірек өзгерді, ДНҚ гистон белоктарымен байланысты болды. Прокариоттық ұйым жасушаішілік симбионттардың геномдарында сақталады.
Симбиогенездің әртүрлі әрекеттерінің нәтижесінде эукариоттық организмдердің әртүрлі топтары пайда болды: эукариот жасуша + цианобактерия = қызыл балдырлар; эукариоттық жасуша + прохлорофитті бактерия = жасыл балдырлар. Тіпті алтын, диатомды, қоңыр және криптомонад балдырларының хлоропласттары екі ретті симбиоздың нәтижесінде пайда болды, бұл 4 мембрананың болуымен дәлелденді.
Эукариоттардың пайда болуы биосфера тарихындағы жағдайлар әсіресе тұрақсыз және болжау мүмкін емес кезеңмен шектелді, прокариоттардың бейімделу стратегиясы (жылдам мутация, көлденең ген алмасу және төзімді клондарды таңдау) тым ысырап және емес. жеткілікті тиімді. Мұндай жағдайда мақсатты түрлендірудің өзгермелілігін дамытуға негізделген түбегейлі әмбебап және үнемді бейімделу стратегиясы үлкен артықшылыққа ие болуы мүмкін.
Мүмкін, эукариоттардың қалыптасуы және олардың жыныстық процесінің дамуы өзгергіштік пен биоәртүрліліктің құрылымын дискретті және «бақыланатын» етті - бұл биоәртүрліліктің жедел өсуіне және түрлердің эволюциялық пластикасы мен экологиялық төзімділігінің артуына әкелуі керек еді. , қауымдастықтар және тұтастай биота.
Эукариоттардың пайда болуын «стандартты» ароморфоз деп атауға болады. Бұл оқиға биологиялық эволюцияның жалпы прогрессивті бағытын айқын көрсетті. Прогресс ұйымның күрделенуінен, тіршіліктің жалпы бейімделу аймағының кеңеюінен, биомасса мен санның өсуінен, организмдердің автономиясының жоғарылауынан ғана емес, сонымен қатар тірі жүйелердің тұрақтылығын арттыруда да байқалды.
Эукариоттардың мысалын қолдана отырып, тіршіліктің жаңа формаларының пайда болуын кейбір жеке филетикалық сызықтардың немесе кладтардың эволюциясының нәтижесі ретінде емес, жоғары дәрежелі жүйелердің дамуының табиғи және болмай қоймайтын әсері ретінде қарастыру керек екендігі анық көрсетілген. - қауымдастықтар, биосфера және, мүмкін, тұтастай алғанда бүкіл планета.
Пайдаланылған дереккөздер:
А.В.Марков, А.М.Куликов. Микробтар қауымдастығындағы интеграциялық процестердің нәтижесінде эукариоттардың пайда болуы
А.В.Марков. Эукариоттардың шығу тегі мәселесі
М.В.Ларина. Эукариоттық жасушалардың пайда болуы туралы гипотезалар. Көп жасушалылықтың пайда болуы
Сабақтың түрі -біріктірілген
Әдістері:ішінара ізденіс, проблемалық баяндау, түсіндірмелі және иллюстративті.
Мақсат:
Студенттерде тірі табиғат, оның жүйелі ұйымдастырылуы мен эволюциясы туралы білімнің тұтас жүйесін қалыптастыру;
Биологиялық мәселелер бойынша жаңа ақпаратқа дәлелді баға бере білу;
Азаматтық жауапкершілікке, дербестікке, бастамашылдыққа тәрбиелеу
Тапсырмалар:
Тәрбиелік: биологиялық жүйелер туралы (жасуша, организм, түр, экожүйе); тірі табиғат туралы қазіргі түсініктердің даму тарихын; биология ғылымындағы көрнекті жаңалықтар; дүниенің қазіргі жаратылыстану картинасын қалыптастырудағы биология ғылымының рөлі; ғылыми танымның әдістері;
Дамужалпы адамзаттық мәдениетке енген биологияның көрнекті жетістіктерін зерттеу үдерісіндегі шығармашылық қабілеттер; әртүрлі ақпарат көздерімен жұмыс істеу барысында қазіргі заманғы ғылыми көзқарастарды, идеяларды, теорияларды, концепцияларды, әртүрлі гипотезаларды (тіршіліктің, адамның мәні мен пайда болуы туралы) дамытудың күрделі және қарама-қайшы жолдары;
Тәрбиетірі табиғатты тану мүмкіндігіне, қоршаған табиғи ортаға және өз денсаулығына қамқорлық қажеттігіне сенімділік; биологиялық мәселелерді талқылау кезінде қарсыластың пікірін құрметтеу
Биологияны оқудың жеке нәтижелері:
1. ресейлік азаматтық тұлғаны тәрбиелеу: патриотизм, Отанға деген сүйіспеншілік пен құрмет, өз Отанына деген мақтаныш сезімі; өз ұлтын білу; көпұлтты ресейлік қоғамның гуманистік және дәстүрлі құндылықтарын ассимиляциялау; Отан алдындағы жауапкершілік пен борыш сезімін тәрбиелеу;
2. білімге деген жауапкершілікті, білім алу мен білімге мотивация негізінде оқушылардың өзін-өзі дамытуға және өзін-өзі тәрбиелеуге дайындығы мен қабілетін қалыптастыру, әлемде бағдарлануға негізделген әрі қарай жеке білім беру траекториясын саналы таңдау және құру. тұрақты когнитивтік қызығушылықтарды ескере отырып, кәсіптер мен кәсіби бейімділіктер;
Биологияны оқытудың метапәндік нәтижелері:
1. оқу мақсаттарын өз бетінше анықтау, оқу-танымдық әрекетте өзіне жаңа мақсаттар қою және тұжырымдау, өзінің танымдық әрекетінің мотивтері мен мүдделерін дамыту;
2. ғылыми-зерттеу және жобалық іс-әрекеттің құрамдас бөліктерін меңгеру, оның ішінде проблеманы көру, сұрақ қою, гипотеза қою;
3. биологиялық ақпараттың әртүрлі көздерімен жұмыс істей білу: әртүрлі дереккөздерден биологиялық ақпаратты табу (оқулық мәтіні, ғылыми-көпшілік әдебиеттер, биологиялық сөздіктер және анықтамалықтар), талдау және
ақпаратты бағалау;
Когнитивті: биологиялық объектілер мен процестердің маңызды белгілерін анықтау; адам мен сүтқоректілер арасындағы қарым-қатынас туралы дәлелдемелерді (аргументтерді) ұсыну; адам мен қоршаған орта арасындағы қарым-қатынас; адам денсаулығының қоршаған орта жағдайына тәуелділігі; қоршаған ортаны қорғау қажеттілігі; биология ғылымының әдістерін меңгеру: биологиялық объектілер мен процестерді бақылау және сипаттау; биологиялық эксперименттер құру және олардың нәтижелерін түсіндіру.
Нормативтік:мақсатқа жету жолдарын, оның ішінде баламалы жолдарды өз бетінше жоспарлау, оқу-танымдық міндеттерді шешудің тиімді жолдарын саналы түрде таңдау қабілеті; мұғаліммен және құрдастарымен оқудағы ынтымақтастықты және бірлескен іс-әрекетті ұйымдастыра білу; жеке және топтық жұмыс: позицияларды үйлестіру және мүдделерді ескере отырып, ортақ шешім табу және қақтығыстарды шешу; ақпараттық-коммуникациялық технологияларды пайдалану саласындағы құзыреттілікті (бұдан әрі – АКТ құзыреттері) қалыптастыру және дамыту.
Коммуникативті:құрбыларымен қарым-қатынас пен ынтымақтастықта коммуникативтік құзыреттілікті қалыптастыру, жасөспірімдік шақтағы гендерлік әлеуметтену ерекшеліктерін түсіну, қоғамдық пайдалы, оқу-зерттеу, шығармашылық және басқа да қызмет түрлері.
Технологиялар : Денсаулықты сақтау, проблемалық, дамыта оқыту, топтық іс-әрекет
Техникалар:талдау, синтездеу, қорытынды жасау, ақпаратты бір түрден екінші түрге аудару, жалпылау.
Тұсаукесер хостингі
«Прокариоттардың күкірт айналымына қатысуы»
Нижний Новгород 2010 ж
Кіріспе
Прокариоттардың (бактериялар мен архейлердің) жер бетіндегі тіршілік үшін маңызы ерекше – олар тіршілікке қажетті негізгі элементтердің (көміртек, оттегі, азот, күкірт, фосфор) циклдік түрленуінде іргелі рөл атқарады. Тірі ағзалар жинақталған элементтердің циклдік түрленулері заттардың айналымын білдіреді. Қазіргі уақытта циклдердің белгілі бір кезеңдерін тек прокариоттар жүзеге асыратыны, биосферадағы негізгі биогендік элементтердің циклдерінің жабылуын қамтамасыз ететіні даусыз дәлелденді. В.И. Вернадский (биосфера туралы ілімнің негізін қалаушылардың бірі) «биосфера өмірдің қабығын – тірі материяның тіршілік ету аймағын білдіреді».
1. Күкірт айналымы
Күкірт – биогенді элемент, тірі заттың қажетті құрамдас бөлігі. Ол ақуыздарда амин қышқылдарының бір бөлігі ретінде кездеседі, ақуыздардағы күкірт мөлшері 0,8-2,4% құрайды. Күкірт сонымен қатар витаминдердің, гликозидтердің, коферменттердің құрамына кіреді және өсімдік эфир майларында кездеседі. Күкірт жер қыртысында, көмірде, тақтатастарда, мұнайларда, табиғи газдарда көптеп кездеседі.
Күкірт – өзгермелі валентті элемент. Бұл оның қозғалғыштығын қамтамасыз етеді. Бейорганикалық қосылыстар түрінде күкірт тотыққан күйде (сульфаттар, политионаттар), тотықсызданған күйде (сульфидтер) және белсенді тотығу-тотықсыздану циклін жүзеге асыратын молекулалық күйде кездеседі. Табиғатта күкірт әртүрлі химиялық және биологиялық өзгерістерге ұшырап, бейорганикалық қосылыстардан органикалық қосылыстарға және кері қарай жылжып, валенттілігі – 2-ден +6-ға дейін өзгереді.
Күкірт қосылыстарының циклдік түрленулері күкірт циклі деп аталады.
Күкірттің айналу цикліне тотығу және тотықсыздану бірліктері, сонымен қатар күкірттің валенттілігін өзгертпей түрленуі жатады. Күкірт циклінің тотығу бөлігі жағдайларға байланысты таза химиялық жолмен де, негізінен микроорганизмдердің қатысуымен болуы мүмкін кезеңдерді қамтиды (бұл жай көзге көрінбейтін организмдер, оның ішінде микроскопиялық эукариоттар: саңырауқұлақтар, балдырлар, қарапайымдар және барлық прокариоттар). Күкірт циклінің тотықсыздандырғыш бөлігі бұл процесте прокариоттардың басым рөлімен биологиялық түрде жүзеге асырылады, ал күкірт атомы максималды тотығу (+ 6) күйінен максималды тотықсыздану (–2) күйіне дейін қалпына келеді. Бірақ бұл процесс әрқашан аяқталмайды, толық емес тотыққан өнімдер қоршаған ортада жиі кездеседі: элементтік күкірт, политионаттар, сульфит.
Осылайша, күкірт айналымы, сондай-ақ заттардың айналымы циклдің жабылуын қамтамасыз ететін прокариоттардың қатысуынсыз мүмкін емес.
2. Қалпына келтіру саласы
Сульфаттың ассимиляциясы.
Сульфатты барлық дерлік өсімдіктер мен микроорганизмдер күкірт көзі ретінде пайдаланады. Ассимиляция кезінде күкірт органикалық қосылыстарға қосылуы үшін сульфат тотықсызданады, өйткені тірі организмдерде күкірт тек сульфгидриль (-SH) немесе дисульфидтік (-S-S-) топтары түрінде тотықсызданған күйде кездеседі. Екі жағдайда да ағзаның өсуіне қажетті мөлшерде күкірті бар қоректік заттар ғана ассимиляцияланады, сондықтан күкірт алмасуының төмендеген өнімдері қоршаған ортаға шығарылмайды. Биосинтез нәтижесінде күкірт негізінен құрамында күкірті бар амин қышқылдарының: цистин, цистеин, метиониннің құрамына кіреді. Құрамында күкірт бар органикалық заттардың құрамына сульфаттардың қатысуы сульфатты ассимиляциялық қалпына келтіру деп аталады.
Күкірттің органикалық қосылыстарының H 2 S түзілуімен конверсиясы.
Күкірт тірі организмдерге негізінен еритін сульфаттар немесе тотықсызданған органикалық күкірт қосылыстары түрінде болады.
Күкіртті органикалық қосылыстардың минералдануы кезінде күкірт бейорганикалық тотықсызданған күйінде Н 2 S түрінде бөлінеді. Аммонификацияға қабілетті сапрофитті микроорганизмдер күкіртті органикалық күкіртті қосылыстардан (тірі ағзалардың зат алмасу өнімдері) бөлуге қатысады. , өлі өсімдіктер мен жануарлар қалдықтары). Аммонификация кезінде құрамында күкірті бар белоктар мен нуклеин қышқылдары ыдырап СО 2, мочевина, органикалық қышқылдар, аминдер және күкірт айналымы үшін маңызды Н 2 S және меркаптандар (тиоспирттер) түзеді. Меркаптандар аэробты жағдайда тотығады да, H2S бөледі.
Белоктардың микроорганизмдермен жойылуы жасушадан тыс процесс ретінде басталады. Бұл жағдайда белоктар протолиттік экзоферменттердің әсерінен жасуша ішіне ене алатын кішірек молекулаларға гидролизденеді және жасушаішілік протеазалар арқылы әрі қарай ыдырауға болатын аминқышқылдарына ыдырайды.
Сульфат пен элементтік күкірттен H 2 S тікелей түзілуі.
Биосферада күкіртсутектің түзілуі негізінен күкірттің дүниежүзілік айналымы үшін үлкен маңызы бар сульфатты қалпына келтіретін бактериялардың белсенділігімен байланысты. Сульфатты қалпына келтіретін бактериялар диссимиляциялық сульфатты қалпына келтіруді жүзеге асырады, бұл органикалық заттардың немесе молекулалық сутегінің тотығуы кезінде сульфат соңғы электронды акцептор (оттегінің орнына) қызметін атқаратын анаэробты тыныс алу. Сондықтан сульфатты қалпына келтіретін бактериялардағы метаболизмнің энергетикалық түрін көбінесе сульфатты тыныс алу деп атайды. Диссимиляциялық сульфатты тотықсыздандыру кезінде сульфатты тотықсыздандыру процесін схемалық түрде келесідей көрсетуге болады: SO 4 2- →SO 3 2- →S 3 O 6 2- →S 2 O 3 2- →S 2-.
Сульфатты тотықсыздандыруға қатысатын ферменттік жүйе екі бөліктен тұрады: біріншісі АТФ-тәуелді процесте сульфатты сульфитке дейін тотықсыздандырады, ал екіншісі алты-электронды тасымалдау арқылы сульфитті сульфидке дейін тотықсыздандырады. Бұл соңғы реакция диссимиляциялық болғандықтан, жасушаны энергиямен қамтамасыз етеді.
Сульфатты қалпына келтіретін бактериялар негізінен облигатты анаэробты бактериялар болып табылады. Сульфатты қалпына келтіретін бактериялардың геохимиялық рөлі өте үлкен, өйткені олардың белсенділігінің арқасында анаэробты аймақтағы инертті қосылыс сульфат күкірттің биологиялық айналымына кең ауқымда қатысады.
Сульфатты қалпына келтіретін бактериялардың белсенділігі әсіресе тоғандар мен бұлақтар түбіндегі лайларда, батпақтарда және теңіз жағалауында байқалады. Теңіз суындағы сульфаттың концентрациясы салыстырмалы түрде жоғары болғандықтан, сульфатты қалпына келтіру теңіз таяздарында органикалық заттардың минералдануының маңызды факторы болып табылады. Мұндай минералданудың белгілері H 2 S иісі және бұл процесс жүретін қою қара лай болып табылады. Шламның қара түсі оның құрамында көп мөлшерде темір темір сульфидінің болуына байланысты. Органикалық заттардың жинақталуы сульфаттың ерекше қарқынды төмендеуіне әкелетін кейбір жағалау аймақтары H 2 S улы әсерінен іс жүзінде жансыз.
Сульфатты қалпына келтіретін бактериялар жүйелі топ емес, физиологиялық болып табылады, өйткені олардың құрамына бір физиологиялық процесті - сульфаттардың қатысуымен анаэробты тыныс алуды, мысалы, Desulfovibrio (vibrios) туысының бактерияларын жүргізуге қабілетті әртүрлі таксономиялық топтардың бактериялары кіреді. Desulfotomaculum (спора түзетін таяқшалар). Архейлер де сульфатты қалпына келтіретін организмдерден табылған. Сульфатты қалпына келтіретін бактериялар тек сульфаттарды ғана емес, тиосульфатты, сульфитті, элементтік күкіртті және басқа да күкірт қосылыстарын қалпына келтіру арқылы көбейе алады.
Кейбір сульфаттарды төмендететін бактериялар үшін күкірт алмасуының негізінен жаңа түрі көрсетілді. Бұл бактериялар органикалық субстраттарда өскенде сульфаттың тотықсыздануы есебінен ғана емес, сонымен қатар тиосульфаттың, сульфиттің, дитиониттің сульфат пен сульфид түзілуімен диспропорциялануы нәтижесінде энергия алуға қабілетті.
Тиосульфаттың жалпы диспропорциялану реакциясы:
S 2 O 3 2- + H 2 0 → SO 4 2- + HS - + H +
Go = – 21,9 кДж/моль S 2 O 3 2-
Жалпы сульфитті диспропорциялау реакциясы:
4SO 3 2- + H + → 3SO 4 2- + HS -
Go = – 58,9 кДж/мольSO 3 2-
Тиосульфатты диспропорциялы бактериялар теңіз шөгінділерінде кең таралғаны анықталды.
Күкіртсутек элементарлы күкіртті тотықсыздандыру кезінде де түзілуі мүмкін. Бүгінгі күні молекулалық күкірттен күкіртсутек түзудің екі механизмі белгілі. Бірінші жағдайда бактериялар мен архейлер молекулалық күкіртті анаэробты тыныс алу (диссимиляциялық күкірт тотықсыздануы) кезінде электронды акцептор ретінде пайдаланады, оның барысында АТФ синтезделеді. Диссимиляциялық күкірт тотықсыздануы – мезофильді және термофильді прокариоттар жүргізетін ферментативті процесс. Екінші жағдайда микроорганизмдер (ашытқылар мен прокариоттар) күкіртті тек ашыту кезінде бөлінген электрондарды шығару үшін пайдаланады (жеңілдетілген ашыту). Бұл АТФ синтезімен бірге жүрмейтін электрондардың бос (бос) разряды.
Молекулярлық күкірттің күкіртсутекке тотықсыздануы, әсіресе күкірттің элементарлы түрі күкірттің ең маңызды түрлерінің бірі болып табылатын және күкірт қосылыстары бастапқыда жанартаудан шыққан гидротермдерде тұратын термофильді микроорганизмдер үшін өте маңызды. Гидротермиялық биоценоздар – тіршілік иелерінің бірегей қауымдастығы. Жоғары температурада (45–50-ден 100°С-қа дейін) дамитын олар негізінен прокариоттар – бактериялар мен архейлерден түзіледі. Гидротермальды сұйықтықтардың микробтық қауымдастығын құрайтын микроорганизмдердің басым көпшілігі басқа жерлерде кездеспейді. Гидротермалардың микробтық қауымдастықтары жер бетіндегі ең ежелгі биоценоздар болып саналады.
3. Тотығу тармағы
Күкірт циклінің бұл бөлігі толығымен күкірттің бейорганикалық қосылыстарының реакцияларынан тұруы мүмкін: S 2- → nS 2- → S 0 → S 2 O 3 2- → SO 3 2- → SO 4 2-, сондай-ақ органикалық реакцияларды қамтуы мүмкін. пішіндер. Органикалық сульфидтердің күкірт атомы әдетте S2 түрінде бөлінгеннен кейін тотығады - бейорганикалық жолмен, дегенмен күкірт атомы органикалық қосылыстардың бөлігі бола отырып, тотыққанда, мысалы, цистеин, таза органикалық тотығу жолы да мүмкін. → цистин. Күкірт қосылыстарының тотығу реакцияларының көпшілігі күшті тотықтырғыштардың (H 2 O 2, O 3, оттегі радикалдары) қатысуымен микроорганизмдердің қатысуынсыз жүруі мүмкін, бірақ микробтық тотығу, әсіресе реагенттің төмен концентрациясында әлдеқайда тиімді.
Күкірт қосылыстарын тотықтыруға қабілетті микроорганизмдердің ішінде үш негізгі физиологиялық топты ажыратады: анаэробты фототрофты бактериялар, аэробты және факультативті анаэробты литотрофты бактериялар, архейлер және әртүрлі гетеротрофты микроорганизмдер.
Анаэробты фототрофты бактериялар.
Бұл өсімдіктер мен цианобактериялардағы аэробты (оттегі) фотосинтезде болатындай, H 2 O орнына электронды донор ретінде әртүрлі тотықсызданған күкірт қосылыстарын қолданып, анаэробты (аноксигенді) фотосинтез жүргізетін бактериялардың ерекше тобы. Тотықсызданған күкірт қосылыстарының ең көп тараған тотығу өнімдері SO 4 2- және S 0, соңғылары кейбір фототрофты бактерияларда жасуша ішінде жинақталуы мүмкін.
Жүйелі түрде фототрофты бактериялар бірнеше топқа бөлінеді: күкіртті және күкіртті емес күлгін және жасыл бактериялар, гелиобактериялар, эритробактериялар (123-сурет). Фототрофты бактериялар пигментті және қоңыр, жасыл немесе күлгін түсті болуы мүмкін. Аноксигенді фотосинтезді жүзеге асыратын фототрофты бактериялардың құрамында оларға сәйкес түс беретін бактериохлорофилл және каротиноидтар болады. Бұл бактериялар жеткілікті жарық түсетін көлдер, тоғандар, лагуналар және минералды бұлақтар сияқты таяз су қоймаларының анаэробты су бағанасында сульфидті тотығуда басым болады. Ерекшелік - эритробактериялар, олар міндетті аэробтар болып табылады және аэробты жағдайда аноксигенді фотосинтезді жүзеге асырады.

Литотрофты күкіртті тотықтырғыш бактериялар және архейлер. Күкірт бактериялары және тион бактериялары.
Кейбір прокариоттар тотықсызданған күкірт қосылыстарының қатысуымен литотрофты өсуге қабілетті. Литотрофты өсу кезінде тотықсызданған күкірт қосылыстары немесе басқа тотықсызданған бейорганикалық қосылыстар (NH 3, H 2, CO, Fe 2+) энергия алмасуында электрон доноры болып табылады, яғни олар электронды тасымалдау тізбегінде тотыққанда АТФ синтезделеді. Литотрофты өсуге тек прокариоттар ғана қабілетті. Литотрофты күкіртке тәуелді прокариоттарда тотықсызданған күкірт қосылыстарының тотығу реакциялары энергия көзі ретінде қызмет етеді. Тотығу реакциялары арнайы ферменттік жүйелердің қатысуымен жүзеге асады. Әртүрлі күкірт қосылыстарының және күкірттің литотрофты прокариоттармен тотығуы әдетте сульфаттардың түзілуіне әкеледі. Бірақ бұл процесс әрқашан аяқтала бермейді, ортада толық емес тотыққан өнімдер (S 0, SO 3 2-, S 2 O 3 2-, S 4 O 6 2-) жиі кездеседі. Литотрофты күкіртке тәуелді прокариоттар тобына негізінен тионды бактериялардың өкілдері, күкіртке өте термоацидофильді архейлер, сутегі бактериялары, түссіз күкірт бактериялары, сонымен қатар күкірт қосылыстарының тотығуынан литотрофты өсуге қабілетті күлгін бактериялардың кейбір өкілдері жатады. қараңғы.
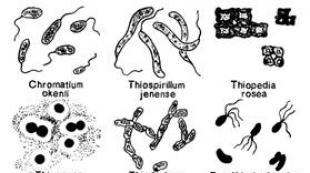
Күкіртсутекті бұлақтарда және құрамында күкіртсутегі бар басқа да су қоймаларында, әдетте, жасушаларында күкірт тамшылары болатын көптеген боялмаған микроорганизмдер кездесетіні бұрыннан белгілі. Күкіртсутегінің концентрациясы салыстырмалы түрде төмен (50 мг/л-ден аз) жерлерде түссіз күкірт бактериялары деп аталатын мұндай микроорганизмдер көбінесе қабықшалар, ақ шөгінділер және басқа ластанулар түрінде массалық жинақтарды түзеді. С.Н. Виноградский (1887) күкірт бактерияларының типтік өкілдерінің бірі, атап айтқанда Беггиатоа жасушаларында тұндырылған күкірт күкіртсутектен түзілетінін және осы микроорганизммен күкірт қышқылына дейін тотыға алатынын дәлелдеді. Жүйелі түрде түссіз күкірт бактериялары гетерогенді топ болып табылады және сипатталған түрлердің және тіпті тектердің барлығын да берік бекітілген деп санауға болмайды. Морфологиясы, қозғалу сипаты, көбею тәсілі және жасуша құрылысы жағынан көп жасушалы да, біржасушалы да (Beggiatoa, Thiothrix, Thiospirillopsis, Thioploca, Achromatium) түссіз күкіртті бактериялардың бірқатар өкілдері көк-жасыл балдырлармен үлкен ұқсастық көрсетеді. Кейбір зерттеушілер, атап айтқанда, Прингшейм (1963) бұл микроорганизмдерді олардың түссіз нұсқалары ретінде қарастырады. Oscillatoria, Thiothrix - Rivularia, Thiospirillopsis - Spirulina көк-жасыл балдырлар Беггиатоа аналогтары болып саналады, ал Achromatium - Synechococcus ұқсас. Көк-жасыл балдырлар қазір бактериялар ретінде жіктелгендіктен, олардың түссіз күкірт бактерияларымен қарым-қатынасы барған сайын ақталуда. Сондай-ақ, кейбір көк-жасыл балдырлардың өз жасушаларында күкіртті тұндыратынын атап өткен жөн, дегенмен бұл қасиет микроорганизмдердің таксономиясын көп қамтамасыз етпейді. Түссіз күкіртті бактерияларға жататын микроорганизмдер тұщы су қоймаларында да, тұзды су қоймаларында да кездеседі. Олардың кейбіреулері төмен температурада жақсы өседі, басқалары (Thiospirillum pistiense) 50°С жоғары температурада термиялық күкіртті бұлақтарда дамиды. Мобильді формаларда хемотаксис бар және оттегі мен күкіртті сутегінің оңтайлы мөлшері бар жерлерге жылжи алады.
Тион деп аталатын бактериялардың көпшілігі типтік хемоавтотрофтар, яғни. олар күкірттің тотықсызданған қосылыстарын тек Н-донорлар ретінде ғана емес, сонымен бірге энергия көзі ретінде де пайдаланады және көмірқышқыл газын ассимиляциялай отырып, таза минералды орталарда өсуге қабілетті. Мұндай бактериялар алғаш рет Неаполь шығанағындағы судан бөлініп алынды (Натансон, 1902) және Тиобациллис (Beijerinck, 1904) атауын алды. Бүгінгі күнге дейін әртүрлі су қоймаларынан, топырақтан, сондай-ақ күкірт пен әртүрлі металдардың шөгінділерінен оқшауланған автотрофты тиобактериялардың көптеген түрлері сипатталған.
Кейбір, негізінен, физиологиялық сипаттамаларға (әртүрлі күкірт қосылыстарын және басқа бейорганикалық субстраттарды тотықтыру қабілеті, органикалық қосылыстарды қолдану, қоршаған ортаның рН мәндеріне, оттегіге қатынасына және т. түрлері ажыратылады. Олардың ішінде ең берік орныққандары болып саналады: Thiobacillus thioparus, T. thiooxidans, T. denitrificans, T. thiocyanoxidans, T. neapolitanus, T. intermedius, T. novellus, T. ferrooxidanas. Бұл микроорганизмдердің барлығы бикарбонат жиі қосылатын қарапайым минералды орталарда өседі. Азоттың көзі әдетте аммоний тұздары болып табылады. Кейбір түрлер мен штаммдар нитраттарды азот көзі ретінде игере алады. Жеке өкілдердің несепнәр мен аминқышқылдарын қолдану қабілеті де көрсетілген. Тұзды су қоймаларынан оқшауланған штамдар өсу үшін патрий хлоридін қажет етеді, ал қаныққан NaCl ерітіндісінде өсетін галофильдер белгілі. Thiobacillus тұқымдас оқшауланған микроорганизмдердің көпшілігінің өсуі үшін оңтайлы температура шамамен 30 ° C құрайды. Дегенмен, табиғи жағдайда олар 55°С-қа дейінгі температурада термалды бұлақтарда кездеседі. Жақында Thiobacillus sp.-ның таза дақылы бөлініп алынды. 50°C өсу үшін оңтайлы.
Қоршаған ортаның қышқылдығына қатысты тиобактериялар бейтарап немесе тіпті сілтілі реакцияда жақсы өсетін организмдерге және ацидофильді түрлерге бөлінеді, яғни. қышқыл ортада дамып, өте төмен рН мәндеріне төзеді.
Бірінші топқа: T. thioparus, T. denitrificans, T. novellus, T. thiocyanoxidans, T. neapolitanus сияқты түрлер жатады. Бұл микроорганизмдер үшін оңтайлы рН мәні 6,0–9,0 аймағында, ал олардың өсуі мүмкін рН мәндерінің аймағы 3,0–6,0-ден 10,0–11,0-ге дейін, ал әртүрлі түрлер мен штаммдар үшін оңтайлы рН мәндері. және. олардың өсуі байқалатын белсенді қышқылдық аймағы айтарлықтай ерекшеленуі мүмкін.
Екінші топқа T.thiooxidans, T.ferroxidans, T. intermedius жатады. Бұл микроорганизмдер үшін оңтайлы рН мәні 2,0–4,0, ал өсу 0,5–2,0-ден 5,0–7,0-ге дейінгі рН кезінде мүмкін болады. Алғашқы екі түрі ең ацидофильді организмдер. Бұл бактериялардың екеуі де 5,0-ден аспайтын рН мәндерінде өседі. Сонымен қатар, T. thiooxidans 1,0 n сәйкес келетін 0-ге жақын рН мәнінде өміршең болып қалатыны көрсетілген. күкірт қышқылы ерітіндісі. Бұл зерттеушілерге белгілі ең ацидофильді микроорганизм болуы мүмкін.
Тионды бактериялардың көпшілігі оттегінің қатысуымен ғана өседі, дегенмен кейбір өкілдерінің өсуі оттегінің төмен деңгейінде мүмкін болады. Бірақ факультативті анаэробтар да белгілі. Оларға T. denirificans жатады. Аэробты жағдайда бұл бактериялар молекулалық оттегінің қатысуымен тотығу процестерін жүргізеді, анаэробты жағдайда олар денитрификацияға ауысады және нитраттарды молекулалық азотқа дейін тотықсыздандырады. Бір қызығы, T. denitrificans, Paracoccus denitrificans сияқты, нитраттарды азот көзі ретінде игере алмайды және өсу үшін азот көзі ретінде аммонийді қажет етеді.
Тионды бактериялар күкіртті сутегі, сульфидтер, сульфит, тиосульфат, тетратионат, тиоцианат (роданид), дитионит сияқты күкіртті қосылыстарды, сондай-ақ толық тотығу кезінде сульфаттар түзетін молекулалық күкіртті тотықтыруға қабілетті. Дегенмен, жеке түрлердің қабілеттері толығымен бірдей емес. Сонымен қатар, күкірттің қандай қосылыстары биологиялық тотығатынын анықтау әрдайым оңай емес, өйткені олардың көпшілігі төмен рН кезінде тұрақсыз және атмосфералық оттегімен де тотығуы мүмкін.
Бейтарап және сілтілі ортада өсетін барлық түрлер, әдетте, күкіртті сутекті, күкіртті және тиосульфатты тотықтырады. T.thiocyanoxidans-қа тән қасиет - тиоцианидті тотықтыру қабілеті. Осы негізде ол жеке түр ретінде ажыратылады, өйткені басқа сипаттамалары бойынша бұл микроорганизм T. thioparus-қа ұқсас. T. thiooxidans сияқты ацидофильді бактерия әдетте құрамында молекулалық күкірті бар ортада өсіріледі. Бұл түрдің күкіртті сутекті және басқа күкірт қосылыстарын тотықтыру қабілеті туралы мәселе толығымен шешілген жоқ, өйткені бұл қосылыстар қышқылдық жағдайда тұрақсыз. T. ferroxidans туралы айтатын болсақ, бұл бактериялардың молекулалық күкіртті де, оның әртүрлі қосылыстарын да тотықтыратыны туралы деректер бар, атап айтқанда: күкіртсутек, тиосульфат, дитионит, тетрасульфат, сульфит. Сонымен қатар, T. ferroxidans суда ерімейтін ауыр металдардың сульфидтерінің тотығуына белсенді қатысады. Оларға пирит (FeS2), халькопирит (CuFeS2), стибнит (SbS2), халькоцит (Cu2S), кавелит (CuS), пирротит (FeS), реалгар (AsS), виоларит (Ni2FeS4) және т.б. минералдар жатады. Қалғандары. бактерияларда не мұндай қабілет жоқ, не әлсіз көрінеді.
T. ferroxidans-тың айрықша қасиеті сонымен қатар темірді темір оксидіне тотықтыру қабілеті болып табылады. Осы негізде бұл бактерияларды темір бактериялары деп санауға болады, бірақ олар Thiobacillus тұқымдасына жатады.
Жоғарыда атап өтілгендей, молекулалық күкірт пен оның әртүрлі қосылыстарының тионды бактериялармен тотығуының соңғы өнімі сульфат болып табылады. Егер процесс осылай жүрсе, яғни. бастапқы субстраттың толық тотығуы жүреді, содан кейін оның нәтижелері келесі теңдеулерде көрсетіледі. Күкіртсутектің тотығуы кезінде:
Аэробты жағдайда күкірт пен тиосульфаттың тотығуы кезінде:

Нитраттарды қолдану арқылы анаэробты жағдайда T.denitrificans күкірт пен тиосульфаттың тотығуы кезінде:

T. thiocyanoxidans тиоцианидтің тотығуы кезінде:

Бірақ тотығу көбінесе аяқталмайды, ортада әр түрлі толық емес тотыққан өнімдер кездеседі. Осылайша, күкіртсутектің тотығуы кезінде кейде молекулалық күкірт пайда болады, тиосульфат пен политионаттар да кездеседі. Молекулярлық күкірттің бактериялармен тотығуы кезінде тиосульфат пен политионаттардың пайда болуы байқалды. Тиосульфаттың тотығуы да жиі политионаттар (тритионат, тетратионат, пейтатиопатат) және элементтік күкірт түзілуімен жүреді. Сульфит те толық тотықпаған өнімдердің бірі болып табылады. Бірақ бұл қосылыстардың барлығы ферментативті процестердің нәтижесі бола алмайды және іс жүзінде бастапқы субстраттың бактериялық тотығуының аралық өнімдеріне жатады. Олардың кейбіреулері таза химиялық жолмен немесе жанама биологиялық реакциялар нәтижесінде түзілген сияқты. Сондықтан әртүрлі күкірт қосылыстарының микроорганизмдер арқылы тотығу жолдарын түсіндіру өте қиын, және барлық реакциялар әлі анықталмаған.
Сульфидтік түрленудің бірінші кезеңінің табиғаты туралы әлі де өте аз деректер бар және күкірт оның тотығуының тікелей немесе жанама өнімі екендігі толық анықталмаған.
Бактериялардың элементарлы күкіртті қолдану мәселесі өте күрделі және оның екі аспектісі бар: микроорганизмдер бұл суда ерімейтін затқа қалай әсер етеді және тотығу процесінің өзі қалай жүреді.
Тиобацилланың күкіртке әсер ету механизміне қатысты екі көзқарас бар.
1. Күкірт бактериялармен тотығуы үшін ол жасушалармен тікелей байланыста болуы керек.
2. Күкіртті бактериялар қоршаған ортаға бөлетін липидті табиғаттағы заттарда алдын ала ерігеннен кейін пайдаланады.
T.thiooxidans молекулалық күкірті бар ортада өскенде, ортада фосфолипидтер кездеседі. Кейбір мәліметтер бойынша бұл фосфатидилинозитол, басқалары бойынша ол сәл өзгеше қосылыстар (фосфатидил-N-метилетанол, фосфатидилглицерин, дифосфатидилглицерин) және олардың жиналуы дақылдардың белсенді өсу фазасына сәйкес келеді. Дегенмен, екінші болжамды дәлелденген деп санауға болмайды. Күкірттің бактериялармен тотығуы үшін оның жасушалармен байланысы да, күкіртті «дымқылдандыратын» кейбір заттардың бөлінуі де маңызды.
Күкірттің тотығу жолына келетін болсақ, деректер де айтарлықтай қарама-қайшы. Күкіртсутегінің тотығуы үшін де қолайлы келесі схема ең ықтимал болып саналады:

X күкіртсутегі немесе күкірт әрекеттесетін глутатион туындысы немесе жасуша мембраналарымен байланысқан тиол деп болжанады. Тиосульфаттың түзілуі бактериялардың қатысуынсыз жүруі мүмкін ферментативті емес процесс болып саналады.
Көптеген зерттеулер тиосульфаттың тотығуын зерттеуге арналған. Бүкіл жасушалармен және тиобацилланың әртүрлі түрлерінің жасушасыз препараттарымен жүргізілген тәжірибелердің деректері тиосульфаттың конверсиясының бастапқы кезеңі оның күкіртсутек пен сульфиттің түзілуімен редуктаза әсерінен тотықсыздануымен байланысты болуы мүмкін екенін көрсетеді:

Немесе элементарлы күкірт пен сульфитке бөлінуімен:

немесе, сайып келгенде, тетратионатқа тотығу арқылы, содан кейін тритионат пен сульфитке айналу арқылы:

Алайда тиосульфатты түрлендірудің соңғы жолы негізгі емес деп болжанады. Тиосульфаттың сульфит түзу үшін ыдырауына келетін болсақ, мұндай реакцияны тиоцианат түріндегі күкіртті байланыстыратын родоназа жүргізуге болады:
Ол тек тионды және басқа бактерияларда ғана емес, жануарларда да кең таралған. Алайда, бұл фермент тионды бактериялардың тиосульфат алмасуына қатыса ма, жоқ па, нақты анықталған жоқ.
Сульфиттің тотығу жолы әлдеқайда көп зерттелген. Пек зертханасында T.thioparus-пен жүргізілген жұмыс сульфиттің аденозинмонофосфатпен (АМФ) әрекеттесетінін, нәтижесінде аденилсульфат немесе аденозинфосфосульфат (АПС) деп аталатын қосылыс түзілетінін көрсетті. Келесі кезеңде АПИ мен бейорганикалық фосфат (Pn) арасында аденозиндифосфаттың (АДФ) түзілуімен және бос сульфаттың бөлінуімен реакция жүреді:

Аденилаткиназа ферментінің әрекеті нәтижесінде АДФ-ның екі молекуласы АТФ-ға айналады және қайтадан АМФ түзеді:
Осылайша, сульфитті тотығудың бұл жолы субстратты фосфорлану деп аталатын нәтижесінде энергияның (АТФ) түзілуімен байланысты. Сонымен бірге сульфитті тотығу процесі электрондардың тыныс алу тізбегіне ауысуымен қатар жүруі мүмкін, оның қызметі АТФ синтезімен байланысты.
Бірқатар тионды бактериялар үшін, соның ішінде T. thioparus үшін электрондардың тыныс алу тізбегіне өтуін қамтамасыз ететін сульфитті тотықтырғыш ферменттің әрекеті нәтижесінде сульфитті тотығу АПС түзілмей-ақ мүмкін болатыны дәлелденген. цитохромдар деңгейінде:
![]()
Мүмкін, бір организмде сульфит пен басқа күкірт қосылыстарының тотығу жолдары әртүрлі болуы мүмкін және оның немесе екіншісінің маңыздылығы қоршаған орта жағдайларына және басқа факторларға байланысты. Қазіргі уақытта қолда бар мәліметтерге сүйене отырып, әртүрлі күкірт қосылыстарының тионды бактериялармен тотығуын келесі жалпылама схемамен көрсетуге болады.

Тионды бактериялардың электронды тасымалдау жүйелерінің құрамдас бөліктеріне келетін болсақ, онда барлық деректер бойынша олардың әр түрдегі құрамына әрқашан с типті цитохромдар кіреді. Сондай-ақ b типті цитохромдар және оның цитохромы деп аталатын нұсқасы да ашылды, ал кейбір өкілдерінде - а және d цитохромдары. Сонымен қатар, электронды тасымалдау жүйесіне флавопротеиндер мен убихинондар кіреді. Бірақ нитрификациялаушы бактериялар сияқты электрондардың оттегіге берілуін қамтамасыз ететін тиондық бактериялардың тыныс алу тізбегі салыстырмалы түрде қысқа, өйткені тотыққан субстраттар жеткілікті жоғары тотығу-тотықсыздану потенциалына ие. Сондықтан бұл микроорганизмдер үшін термодинамикалық градиентке қарсы энергияға тәуелді электрон тасымалдау (кері электронды тасымалдау) маңызды. Ол көмірқышқыл газын және басқа да конструктивті процестерді ассимиляциялау үшін қажетті азайған NAD түзілуін қамтамасыз етеді. Сондай-ақ электрондарды тасымалдау жүйелерінің құрамдас бөліктері мен электрондарды тасымалдау жолдары тиоп бактерияларының әртүрлі өкілдерінде айқын айырмашылықтарға ие болуы мүмкін емес, сонымен қатар тотыққан субстраттың табиғатына байланысты болуы мүмкін екенін атап өткен жөн. Сондықтан жалпыланған диаграмма (140-сурет) өте шартты.

Әртүрлі авторлардың есептеулері бойынша тиондық бактериялардың бос энергияны пайдалану тиімділігі 2-ден 37% -ға дейін. Басқа хемоавтотрофтар сияқты, тиондық бактериялар көмірқышқыл газын ең алдымен Кальвин циклі арқылы ассимиляциялайды. Бірақ олардың белгілі бір метаболиттердің түзілуі үшін маңызды басқа да карбоксилдену реакцияларын жүргізу қабілеті бар.
Күкіртке өте термоацидофильді архейлер элементтік күкірт болған кезде де литотрофты өсуге қабілетті және 40–100 °C және рН 1–6 дами алады; бұлар гидротермдердің және жанартаулық кальдераның тұрғындары.
Гетеротрофты күкіртті тотықтырғыш микроорганизмдер (қоректену үшін органикалық заттарды пайдаланатын организмдер).
Кейбір облигатты гетеротрофты бактериялардың, ашытқылардың және микромицеттердің әртүрлі бейорганикалық күкірт қосылыстарын тотықтыру қабілеті біршама уақыт бойы белгілі болды, дегенмен бұл реакциялардың гетеротрофтардың метаболизміндегі маңызы әлі толық зерттелмеген. Гетеротрофты күкіртті тотықтырғыш бактериялардың ішінде тиосульфаттың тетратионатқа дейін толық емес тотығуын жүзеге асыратын псевдомонадтар тобының түрлері басым.
Сондай-ақ С.Н. Виноградский (1887–1889) түссіз күкірт бактерияларының құрамында органикалық заттар өте аз мөлшерде болатын суда өсетінін атап өтті, сондықтан олар көмірқышқыл газын сіңіре алады деп есептеді. Беггиатоаның әртүрлі штаммдарының физиологиясын зерттеуге сүйене отырып, Прингхайм олардың арасында күкіртті сутекті тотықтыратын және СО 2 бекітетін автотрофтар бар және органикалық қосылыстарды қажет ететін өкілдері бар деп есептейді. Бірақ бірқатар гетеротрофты Беггиатоа штамдары да органикалық қосылыстардың қатысуымен күкіртсутекті тотықтырады, мүмкін, энергия шығарады, т. олар хемолитогетеротрофтар. Түссіз күкірт бактерияларымен қатар күкіртсутектің, молекулалық күкірттің және тиосульфаттың тотығуына қатысатын типтік гетеротрофты микроорганизмдер белгілі. Оларға Bacillus, Pseudomonas, Achromobacter, Sphaerotilus өкілдері, сонымен қатар актиномицеттер, зеңдер (Penicillium luteum, Aspergillus niger), ашытқылар және Alternaria жатады. Олардың кейбіреулері, атап айтқанда, жіп тәрізді көп жасушалы бактериялар Sphaerotilus natans күкіртті күкіртті сутегінің қатысуымен жасушаларға орналастырады. Басқалары (Pseudomonas aeruginosa, Ps. fluorescens, Achromobacter stuzeri) тиосульфатты тетратионатқа (Na2S4O6) дейін тотықтыруға қабілетті. Политионаттар мен сульфаттардың түзілуі гетеротрофты микроорганизмдердің аралас дақылдары элементарлы күкіртпен әрекеттескенде де байқалды. Тотығу механизмі және гетеротрофтар үшін бұл процестің биологиялық маңызы әлі де түсініксіз. Кейбір гетеротрофты түссіз күкірт бактериялары органикалық заттардың тотығуы кезінде жасушаларда түзілетін супероксид радикалы - сутегі асқын тотығымен күкірт қосылыстарын тотықтыруға қабілетті: H 2 O 2 + H 2 S → S 0 + 2H 2 O. Бұл процестің физиологиялық мәні. оттегінің толық емес тотықсыздануының улы өнімдерін детоксикациялау болып табылады ( H 2 O 2 , O 2-).
Күкірт қосылыстарының сульфатқа дейін толық тотығуы элементарлы күкіртпен байытылған топырақта тіршілік ететін микромицеттер үшін көбірек тән. Aspergillus, Penicillium, Trichoderma, Fusarium, Mucor және Auerobasidium микроскопиялық саңырауқұлақтардың кейбір түрлері үшін элементтік күкіртті, тиосульфатты және тіпті металл сульфидтерін сульфатқа тотықтыру қабілеті көрсетілген, бірақ мұндай тотығу жылдамдығы бір немесе екі ретті құрайды. литотрофты бактерияларға қарағанда төмен.
күкіртті циклдік түрлендіру микроорганизмі
Қорытынды
Соңғы онжылдықтарда күкірттің табиғи айналымы улы күкірт қосылыстарының жинақталуына және күкірттің табиғи айналымының теңгерімсіздігіне әкелетін антропогендік әсердің күшеюіне ұшырады. Атап айтқанда, күкірт қосылыстарының ауқымды шығарындылары қазба отындарын жағу кезінде, целлюлоза-қағаз және металлургия кәсіпорындарынан бөлінетін күкіртті сутегі мен ұшпа органикалық сульфидтерді жағу кезінде, сондай-ақ коммуналдық қалдықтардың ыдырауы кезінде жылу электр станциялары бөлетін күкірт қос тотығының түзілуіне әкеледі. және ауылшаруашылық сарқынды сулары. Бұл қосылыстар микрограмм концентрациясында да улы болып табылады. Олар ауаны уландырып, атмосфералық химияға әсер етіп, суда еріген оттегінің жетіспеушілігін тудыруы мүмкін.
Сондықтан күкірт қосылыстарының өзгеруіне және улы күкірт қосылыстарын детоксикациялауға қатысатын прокариоттарды қолдану өте өзекті болып көрінеді. Сонымен қатар, микроорганизмдердің өте жоғары каталитикалық белсенділігі олардың жер бетінде болатын химиялық өзгерістерде үлкен рөл атқаруының себебі болып табылады. Микроорганизмдер шағын өлшемдеріне байланысты жануарлар мен жоғары сатыдағы өсімдіктермен салыстырғанда беттік-көлемдік қатынасы жоғары, бұл жасуша мен қоршаған орта арасында субстраттардың және экскреторлық өнімдердің жылдам алмасуына әкеледі.
Сондай-ақ олардың қолайлы жағдайларда көбеюінің жоғары қарқыны және бүкіл биосферада кең таралуы маңызды факторлар болып табылады. Дегенмен, прокариоттардың қоректік заттардың айналуындағы ерекше маңыздылығына қарамастан, заттардың айналымындағы, атап айтқанда күкірт цикліндегі прокариоттардың белсенділігінің ауқымы әлі толық бағаланбаған.
Әдебиет
1. Грабович М.Ю. Прокариоттардың күкірт айналымына қатысуы // Сорос білім беру журналы, 1999, № 12, б. 16–20.
2. Громов Б.В., Павленко Г.В. Бактериялар экологиясы. Л.: Ленинград мемлекеттік университетінің баспасы, 1989. 248 б.
3. Громов Б.В. Архейдің таңғажайып әлемі // Сорос білім беру журналы. 1997. № 4. 23–26 беттер.
4. Кальдера микроорганизмдері / Ред. Г.А. Заварзина. М.: Наука, 1989. 120 б.
5. Кондратьева Е.Н. Хемолитотрофтар және метилотрофтар. М.: ММУ баспасы, 1983. 172 б.
6. Малахов В.В. Vestimentifera - автотрофты жануарлар // Сорос білім беру журналы. 1997. № 9. 18–26 беттер.
Планетаның биосферасы – бөліктері бір-бірімен үйлесімді байланысқан біртұтас мегаорганизм. Тіршіліктің барлық алуан түрлілігі екі суперпатшалыққа бөлінеді - прокариоттар (ядроға дейінгі организмдер) және эукариоттар (ядросы бар). Аса патшалықтар тірі патшалықтарға бөлінеді:
- вирустар;
- бактериялар;
- саңырауқұлақтар;
- өсімдіктер;
- жануарлар.
Бактериялар патшалығы вирустар патшалығымен бірге прокариоттардың суперпатшалығына – ядросыз организмдерге біріктірілген. Тарихи тұрғыдан бұл планетадағы тірі ағзалардың бірінші эшелоны.
Бір жасушалы тірі организмдер шамамен 3,8 миллиард жыл бұрын пайда болды. Бір миллиард жылға жуық уақыт бойы олар планетаның жалғыз тірі тұрғындары болды - олар сәтті өсті, дамыды және бейімделді. Олардың тіршілік әрекетінің нәтижесі планетаның атмосферасында бос оттегінің пайда болуы болды, бұл көп жасушалы организмдердің - саңырауқұлақтардың, өсімдіктердің, гүлдер мен жануарлардың пайда болуына мүмкіндік берді.
Қазіргі уақытта тірі бактериялар барлық жерде өмір сүреді: сирек кездесетін атмосфералық қабаттардан ең терең мұхиттық ойпаттарға дейін, олар Арктикалық мұзда және термиялық гейзерлерде өмір сүреді. Бактериялар тек бос кеңістікті ғана емес, олар саңырауқұлақтар, өсімдіктер немесе жануарлар болсын, басқа организмдердің ішінде өседі.

Табиғатта барлық жануарлар, адамдар да ерекшелік емес, микробтардың мекендейтін ортасы болып табылады:
- тері;
- ауыз қуысы;
- ішектер.
Ғалымдар адам ағзасында өмір сүретін микроорганизм жасушаларының саны өз жасушаларының санынан 10 есе көп екенін анықтады. Осындай жоғары сандық көрсеткіштерге қарамастан, денеде өмір сүретін бактериялардың салмағы 2 кг-нан аспайды - бұл жасуша мөлшерінің айтарлықтай айырмашылығына байланысты.
Бактериялар патшалығының тірі өкілдерінің сансыз түрлері бар, бірақ олардың барлығына ортақ нәрсе:
- айқын ядроның болмауы;
- өте кішкентай (өсімдіктер мен жануарлар жасушаларымен салыстырғанда) жасуша өлшемдері;
- Биологиялық бірлік - бұл жасушаның өзі, олар біріктірілген кезде біз бактериялардың колониясы туралы айтамыз.
Бұл саңырауқұлақтардың, өсімдіктердің және жануарлардың пайда болуына мүмкіндік берген бактериялар патшалығының өкілдері болды. Ғаламшарда пайда болған микроорганизмдер бар жағдайларға бейімделіп қана қоймай, олар өмір сүру ортасын белсенді түрде өзгертіп, сапалы жаңа сипаттамалар жасады.

Табиғаттағы азот пен көміртегінің айналымы тек микроорганизмдердің арқасында жүреді. Ғалымдар биосферадан микробтар жойылса, планетадағы тіршілік өмір сүре алмайтынын анықтады.
Биосфера циклдеріндегі прокариоттардың рөлі
Жер бетінде тіршілік пайда болған кезде бактериялар патшалығының өкілдері биосфераның қалыптасуына белсенді түрде қатысты. Қазіргі биосфераның жұмыс істеу деңгейін ұстап тұру үшін микроорганизмдер қажет - табиғаттағы энергия мен заттардың айналымы микробтармен қамтамасыз етіледі.
Биосфералық процестердегі тірі микробтардың басым рөлінің мысалдары топырақтың құнарлы қабатын құру және оны ұстау болып табылады.
Микроорганизмдердің газ және тотығу функцияларынан басқа геохимиялық қызметтерінің маңызы арта түсуде. Ферменттердің белсенділігі мен концентрация функциялары планетаның геохимиясына айтарлықтай әсер етеді.
Микроорганизмдердің түр алуандығы
Бактериялар патшалығының өкілдері биосфераның барлық деңгейлеріне қоныстанған және микробтардың болуы арқылы планетаның биосферасының жоғарғы және төменгі шекараларын анықтайды. Осындай әртүрлі физикалық параметрлері бар орталарда өмір сүретін микробтар бірқатар сипаттамалармен ерекшеленеді.
- Тірі бактерия жасушасының пішіні бойынша:
- сфералық кокктар;
- таяқша тәрізді;
- бұралған, вибриондар мен спирохеталарға бөлінеді.
- Дененің кеңістікте қозғалысына қарай:
- флагелласыз (броунға ұқсас хаотикалық қозғалыс);
- флагелланы пайдалану (бүкіл периметр бойынша сан біреуден көпке дейін өзгереді).
- Бактериялар патшалығы өкілдерінің метаболикалық ерекшеліктеріне сәйкес:
- бейорганикалық заттардан – автотрофтардан қажетті заттарды синтездеу;
- органикалық заттарды өңдеу – гетеротрофтар.
- Энергия алу әдісі бойынша:
- тыныс алу (аэробты және анаэробты микроорганизмдер);
- ашыту;
- фотосинтез (оттегісіз және оттегісіз).

Микробтар мен вирустар арасындағы қарым-қатынас ерекшеліктері – аттас патшалықтардың өкілдері
Прокариоттардың суперпатшалығы екі патшалықты - бактериялар мен вирустарды біріктіреді, олардың ұқсастықтарынан әлдеқайда көп айырмашылықтары бар. Мысалы, бактериялар тіршілікті қамтамасыз етуге қажетті заттардың барлығын синтездесе, вирустар ақуыз синтезіне мүлде қабілетсіз. Олар тіпті өз түрін өздігінен көбейте алмайды, тек біреудің жасушасына ену арқылы ғана.
Вирустар қабылдаушы жасушаның ДНҚ-сын блоктайды және оны өздерімен ауыстырады - нәтижесінде басып алынған жасуша шабуылшы вирустың көшірмелерін жасайды, бұл әдетте оның өліміне әкеледі.