Bakteriyalar qirolligi tirik organizmlarining biosferadagi o'rni. Annotatsiya: Prokariotlarning oltingugurt siklidagi ishtiroki
Savol 1. Prokariotlarning biosferadagi rolini aytib bering.
Prokaryotlar fotosintezni amalga oshiradilar va shu bilan atmosferaga kislorod chiqaradilar. Xemosintez prokariotlar orasida keng tarqalgan. Bundan tashqari, bakterial organizmlar orasida azot biriktiruvchi shakllar mavjud: bu bizning sayyoramizdagi azotni to'g'ridan-to'g'ri atmosfera havosidan o'zlashtirishga qodir bo'lgan va shu tariqa molekulyar azotni biologik tsiklga jalb qiladigan yagona tirik organizmlar guruhidir.
Prokariotlarning yana bir muhim vazifasi ham bor: organik birikmalarni yo‘q qilish (minerallash) orqali anorganik moddalarni atrof-muhitga qaytarish.
Umuman olganda, biosfera darajasida prokaryotlar, birinchi navbatda, bakteriyalar yana bir muhim funktsiyaga ega - kontsentratsiya. Ular juda past konsentratsiyalarda ham atrof-muhitdan ma'lum elementlarni faol ravishda ajratib olishga qodir.
Prokaryotlarning xususiyatlari va funktsiyalari shunchalik xilma-xilki, ular printsipial jihatdan barqaror ishlaydigan ekotizimlarni yaratishga qodir (ya'ni faqat ularning ishtiroki bilan).
2-savol. Prokaryotik dorilarni tabiiy muhitga uning ifloslantiruvchi moddalariga qarshi kurashish uchun kiritish qanday xavfli ekanligini tushuntiring.
Prokariotlarni atrof-muhitga kiritishda yuzaga keladigan muammoli masalalardan biri mahalliy mikroflora va kiritilgan mikroflora o'rtasidagi munosabatlarning tabiatini aniqlashdir. Mikrobiotsenozlarda mavjud muvozanatning o'zgarishi natijasida kiritilgan mikroorganizmlarning tabiiy biogeotsenozlar holatiga salbiy ta'sir qilish ehtimolini istisno qilib bo'lmaydi. Bu holat juda ehtiyotkorlikni talab qiladi va faqat sinchkovlik bilan tadqiqotdan so'ng prokaryotlardan inson ehtiyojlari uchun va biosferani ifloslantiruvchi moddalarga qarshi kurashda foydalanishni talab qiladi.
Savol 3. Ma'lumki, barcha eng xilma-xil azotni biriktiruvchi bakteriyalar molekulyar azotning bog'lanishini katalizlovchi bir xil ferment, nitrogenazaga ega. Bu faktning tirik tabiat evolyutsiyasida qanday ahamiyati bor?
Nitrogenaza ikkita oqsil majmuasidan tashkil topgan ko'p o'lchovli fermentdir. Turli xil azot fiksatorlaridan olingan nitrogenazlar o'zlarining molekulyar og'irliklari va metall tarkibi bilan bir oz farq qiladi. Bu fakt azot saqlovchi bakteriyalarning kelib chiqish birligini ko'rsatadi. Nitrogenaza bakteriyalarning ajdodlari shaklida paydo bo'ldi, keyin divergentsiya sodir bo'ldi, bu esa azotni biriktiruvchi bakteriyalarning xilma-xilligiga olib keldi, ularning har birida nitrogenazani kodlovchi gen mavjud edi.
Azotni biriktirish jarayoni normal atmosfera bosimi va normal haroratda sodir bo'ladi, lekin juda ko'p energiya talab qiladi.
Nitrogenazning faol ishlashi uchun mikroaerofil sharoitlar talab qilinadi. Molekulyar kislorod ikkala nitrogenaza oqsiliga ham zararli ta'sir ko'rsatadi. Tugundagi mikroaerofil sharoitlar kislorodga diffuz to'siq bilan ta'minlanadi. Havodan ammiak ishlab chiqaradigan nitrogenaza kompleksi juda tejamkor ishlaydi. Agar yashash joyida ammoniy yoki nitrat ionlari etarli bo'lsa, u ishlashni to'xtatadi.
Azot barcha tirik organizmlar uchun mutlaqo zarur element hisoblanadi. Azotning asosiy ombori yer atmosferasidir. Eukaryotik organizmlar azotni bevosita atmosferadan o'zlashtira olmaydi. Azot biriktiruvchi bakteriyalarda molekulyar azotning bog‘lanishini katalizlovchi nitrogenaza fermenti mavjudligi sababli ular bilan simbiozda yashovchi o‘simliklar o‘zlari uchun oson bo‘lgan azotni olishlari mumkin. Ko'pgina o'simliklar, hayvonlar va zamburug'lar azotni biriktiruvchi prokariotlar bilan simbiozga kirish qobiliyatiga ega.
4-savol. 70-yillarning boshlarida amerikalik biolog va ekolog Barri Kommoner. XX asr Ekologiya qonuni sifatida tezisni ilgari surdi: "Tabiat eng yaxshi biladi". Nima demoqchi ekanligingizni tushuntiring. Ushbu masala bo'yicha fikringizni bildiring. Javobingizni misollar bilan tushuntiring.
Barri Kommonerning ta'kidlashicha, tabiatning mexanizmlari va funktsiyalari haqida mutlaqo ishonchli ma'lumotlar mavjud bo'lmaguncha, biz soatning tuzilishi bilan tanish bo'lmagan, lekin uni tuzatishni xohlaydigan odam kabi, ularni yaxshilashga harakat qilib, tabiiy tizimlarga osonlikcha zarar etkazamiz. U juda ehtiyot bo'lishga chaqiradi. Tabiatning o'zgarishi iqtisodiy jihatdan halokatli va ekologik xavflidir. Oxir-oqibat, hayot uchun mos bo'lmagan sharoitlar yaratilishi mumkin. Tabiatni yaxshilashning ekologik mezonini ko'rsatmasdan turib, tabiatni yaxshilash haqidagi mavjud fikr ma'nosizdir.
Men bu fikrga qo'shilaman. Biz ko'pincha tabiatning bir ko'rinishi biotsenozlar, biogeotsenozlar yoki umuman biosfera uchun ko'proq halokatli va zararli hodisaning oqibati deb o'ylamaymiz. "Asosiysi, biz o'zimizni yaxshi his qilamiz" va tabiat kutadi. Siz bunday qila olmaysiz!!!
Siz "o'rmon buyurtmachilari" bo'lib chiqqan bo'rilarni otish misollarini keltirishingiz mumkin. Xitoyda chumchuqlar yo'q qilindi, ular go'yoki ekinlarni yo'q qiladi, ammo hech kim qushsiz ekinlar zararli hasharotlar tomonidan yo'q qilinadi deb o'ylamagan.
Aynan eukaryotik hujayraning paydo bo'lishi biologik evolyutsiyadagi eng muhim voqealardan biridir. Eukaryotik va prokaryotik organizmlar o'rtasidagi farq genomni tartibga solishning yanada rivojlangan tizimidir. Va shu tufayli bir hujayrali organizmlarning moslashuvchanligi, ularning genomga irsiy o'zgarishlar kiritmasdan o'zgaruvchan atrof-muhit sharoitlariga moslashish qobiliyati oshdi. Moslashish qobiliyati tufayli eukariotlar ko'p hujayrali bo'lishga muvaffaq bo'ldi - ko'p hujayrali organizmda bir xil genomga ega bo'lgan hujayralar, sharoitlarga qarab, morfologiyasi va funktsiyasi jihatidan butunlay boshqacha bo'lgan to'qimalarni hosil qiladi.
Ushbu aromorfoz arxey va proterozoy (2,6 - 2,7 milliard yil oldin) burilishlarida sodir bo'lgan, bu biomarkerlar tomonidan aniqlangan - faqat eukaryotik hujayralarga xos bo'lgan steroid birikmalarining qoldiqlari. Eukariotlarning paydo bo'lishi kislorod inqilobiga to'g'ri keladi.
Eukariotlar prokariotlarning bir nechta navlarining simbiozi natijasida paydo bo'lganligi odatda qabul qilinadi. Ko'rinib turibdiki, mitoxondriyalar alfa-proteobakteriyalardan (aerob eubakteriyalar), plastidlar siyanobakteriyalardan va sitoplazma noma'lum arxebakteriyalardan kelib chiqqan. Yadro, sitoskeleton va flagellaning kelib chiqishi haqida umumiy qabul qilingan nazariya hali mavjud emas. Yerda hayotning kelib chiqishi haqidagi farazlar hujayraning kelib chiqishi haqidagi savolga oydinlik kiritmadi. Agar prokaryotlarning kelib chiqishi to'g'risida ularning paydo bo'lishini to'g'ri tavsiflovchi farazlar deyarli bo'lmasa, eukaryotik hujayralarning kelib chiqishiga nisbatan bir nechta nuqtai nazarlar mavjud.
Eukariotlarning kelib chiqishi haqidagi asosiy farazlar:
1. Simbiotik gipoteza ikki tushunchaga asoslanadi. Ushbu tushunchalarning birinchisiga ko'ra, tirik tabiatdagi eng asosiy farq bakteriyalar va haqiqiy yadroli hujayralardan tashkil topgan organizmlar - protistlar, hayvonlar, zamburug'lar va o'simliklar o'rtasidagi farqdir. Ikkinchi tushuncha eukaryotik hujayralarning ba'zi qismlarining kelib chiqishi simbiozlarning evolyutsiyasi - har xil turdagi organizmlar o'rtasida doimiy assotsiatsiyalarning shakllanishi edi. Taxminlarga ko'ra, organellalarning uchta sinfi - mitoxondriyalar, kiprikchalar va fotosintetik plastidlar - erkin yashovchi bakteriyalardan kelib chiqqan bo'lib, ular simbioz natijasida prokaryotik xost hujayralariga kiritilgan. Bu nazariya ko'p jihatdan Mendel genetikasini Darvinning tabiiy tanlanish g'oyasi bilan bog'lagan genetiklar, ekologlar va sitologlar tomonidan ishlab chiqilgan neodarvin g'oyalariga tayanadi. Shuningdek, u molekulyar biologiyaga, ayniqsa oqsillar va aminokislotalar ketma-ketligi haqidagi ma'lumotlarga, Yerdagi hayotning eng qadimgi izlarini o'rganuvchi mikropaleontologiyaga va bu fanlar biologik kelib chiqadigan gazlar bilan shug'ullanar ekan, atmosfera fizikasi va kimyosiga tayanadi.
2.Invaginatsiya gipotezasi eukaryotik hujayraning ajdod shakli aerob prokariot bo'lganligini aytadi. Ichkarida hujayra membranasiga biriktirilgan bir nechta genom bor edi. Korpuskulyar organellalar va yadro qobiq bo'limlarining invaginatsiyasi va bog'lanishi, so'ngra yadro, mitoxondriya va xloroplastlarga funktsional ixtisoslashuvi natijasida paydo bo'lgan. Keyinchalik evolyutsiya jarayonida yadro genomi murakkablashib, sitoplazmatik membranalar tizimi paydo bo'ldi. Bu gipoteza qobiqlarda yadro, mitoxondriya, xloroplastlar va ikkita membrananing mavjudligini tushuntiradi. Ammo u korpuskulyar organellalardagi oqsil biosintezi jarayoni va eukariot hujayra sitoplazmasidagi tafovutlarni tushuntirishda qiyinchiliklarga duch keladi. Mitoxondriya va xloroplastlarda bu jarayon zamonaviy prokaryotik hujayralardagi jarayonga to'liq mos keladi.
Simbiotik (I) va invaginatsiya (II) gipotezalariga ko'ra eukaryotik hujayraning kelib chiqishi:
1 - anaerob prokariot (xo'jayin hujayra), 2 - mitoxondriyali prokariotlar, 3 - ko'k-yashil suv o'tlari (taxminan xloroplast), 4 - siiroketaga o'xshash bakteriya (taxminan flagellum), 5 - flagellumli ibtidoiy eukariot, 6 - o'simlik 7 - flagellumli hayvon hujayrasi, 8 - aerob prokariot (taxminiy mitoxondriya), 9 - aerob prokariot (II gipoteza bo'yicha progenitör hujayra), 10 - yadro va mitoxondriyani yaratgan hujayra membranasining invaginatsiyasi, 11 - primitiv. eukariot 12 - xloroplast, 13 - o'simlik hujayrasini bergan hujayra membranasining invaginatsiyasi; a - prokariot hujayra DNKsi, b - mitoxondriya, c - eukariot hujayra yadrosi, d - flagellum, e - xloroplast.
Mavjud ma'lumotlar gipotezalardan biriga ustunlik berish yoki ko'pchilik olimlarga mos keladigan yangisini ishlab chiqish uchun hali etarli emas, ammo so'nggi yillarda eukaryotik hujayraning kelib chiqishining simbiogenetik nazariyasini ishonchli tarzda isbotlash mumkin bo'ldi.
Eukaryotik hujayralarning evolyutsion qobiliyati prokaryotik hujayralarga qaraganda yuqori. Bu erda etakchi rol eukariotlarning yadro genomiga tegishli bo'lib, u prokariotlarning genomidan kattaroqdir. Muhim farqlar - yadrolarda ikkita gen to'plamining mavjudligi sababli eukaryotik hujayralarning diploidligi, shuningdek, ba'zi genlarning ko'p marta takrorlanishi.
Hujayra faoliyatini tartibga solish mexanizmi murakkablashib bormoqda, bu tartibga soluvchi genlarning nisbiy sonining ko'payishi va prokaryotlarning dumaloq "yalang'och" DNK molekulalarini DNK oqsillar bilan bog'langan xromosomalar bilan almashtirishda namoyon bo'ladi.
Aerob nafas olish ham ko'p hujayrali shakllarning rivojlanishi uchun zarur shart bo'lib xizmat qildi. Eukaryotik hujayralar atmosferada O 2 kontsentratsiyasi 1% ga (Paster nuqtasi) etganidan keyin Yerda paydo bo'lgan. Va bu kontsentratsiya aerob nafas olish uchun zaruriy shartdir.
Ma'lumki, har bir eukaryotik hujayra turli xil kelib chiqadigan genomlarni o'z ichiga oladi: hayvonlar va qo'ziqorin hujayralarida bu yadro va mitoxondriya genomlari, o'simlik hujayralarida esa bular ham plastid genomlaridir. Kichik dumaloq DNK ham eukaryotik hujayralar flagellasining bazal tanasida mavjud.
Molekulyar soat usuliga ko'ra, eukariotlar prokariotlar bilan bir vaqtda paydo bo'lgan. Ammo Yer tarixining muhim qismida prokaryotlar hukmronlik qilgani aniq. Eukaryotik o'lchamlarga mos keladigan birinchi hujayralar (akritarxlar) 3 milliard yoshda, ammo ularning tabiati hali ham aniq emas. Eukaryotlarning deyarli ma'lum qoldiqlari taxminan 2 milliard yil. Va faqat kislorod inqilobidan so'ng, sayyora yuzasida (taxminan 1 milliard yil oldin) eukaryotlar uchun qulay sharoitlar paydo bo'ldi.
Ehtimol, eukaryotik hujayralarning asosiy ajdodi arxebakteriyalar bo'lib, ular oziq-ovqat zarralarini yutish orqali ovqatlanishga o'tgan. Bunday yutish uchun zarur bo'lgan hujayra shaklining o'zgarishi aktin va miyozindan tashkil topgan sitoskelet tomonidan ta'minlangan. Bunday hujayraning irsiy apparati membrana bilan aloqasini saqlab, o'zgaruvchan yuzasidan chuqurroq harakat qildi. Va bu allaqachon yadro teshiklari bo'lgan yadro qobig'ining paydo bo'lishiga olib keldi.
Xost hujayra tomonidan so'rilgan bakteriyalar uning ichida mavjud bo'lishi mumkin. Shunday qilib, mitoxondriyalarning ajdodlari fotosintetik bakteriyalar guruhiga aylandi - binafsha alfaproteobakteriyalar. Xost hujayra ichida ular fotosintez qilish qobiliyatini yo'qotib, organik moddalarning oksidlanishini o'z zimmalariga oladilar. Ular tufayli eukaryotik hujayralar aerobga aylandi. Boshqa fotosintetik hujayralar bilan simbiozlar o'simlik hujayralarining plastidlarga ega bo'lishiga olib keldi. Ehtimol, eukaryotik hujayralarning flagellasi mezbon hujayralar va harakatga qodir bo'lgan bakteriyalar o'rtasidagi simbioz natijasida paydo bo'lgan.
Eukaryotik hujayralarning irsiy apparati prokariotlarniki bilan taxminan bir xil tuzilgan. Ammo kattaroq va murakkabroq hujayrani boshqarish zarurati tufayli xromosomalarning tashkil etilishi keyinchalik o'zgardi va DNK giston oqsillari bilan bog'landi. Prokaryotik tashkilot hujayra ichidagi simbiontlarning genomlarida saqlanadi.
Turli xil simbiogenez harakatlari natijasida eukaryotik organizmlarning turli guruhlari paydo bo'ldi: eukaryotik hujayra + siyanobakteriyalar = qizil suv o'tlari; eukaryotik hujayra + proxlorofit bakteriya = yashil suv o'tlari. Hatto oltin, diatom, jigarrang va kriptomonad suvo'tlarining xloroplastlari ham ketma-ket ikkita simbioz natijasida paydo bo'lgan, bu 4 ta membrananing mavjudligidan dalolat beradi.
Eukariotlarning paydo bo'lishi biosfera tarixidagi sharoitlar ayniqsa beqaror va oldindan aytib bo'lmaydigan davrga to'g'ri keldi, prokaryotlarning adaptiv strategiyasi (tez mutatsiya, gorizontal gen almashinuvi va chidamli klonlarni tanlash) haddan tashqari isrof bo'lib chiqdi. etarlicha samarali. Bunday vaziyatda, maqsadga muvofiq o'zgartirish o'zgaruvchanligini rivojlantirishga asoslangan printsipial jihatdan universalroq va iqtisodiy moslashuvchan strategiya katta afzalliklarga ega bo'lishi mumkin.
Ehtimol, eukaryotlarning shakllanishi va ularning jinsiy jarayonining rivojlanishi o'zgaruvchanlik va biologik xilma-xillik strukturasini yanada diskret va "boshqariladigan" qildi - bu bioxilma-xillikning tez o'sishiga va turlarning evolyutsion plastikligi va ekologik bardoshliligining oshishiga olib kelishi kerak edi. , jamoalar va umuman biota.
Eukaryotlarning paydo bo'lishini haqli ravishda "standart" aromorfoz deb atash mumkin. Ushbu hodisa biologik evolyutsiyaning umumiy progressiv yo'nalishini aniq ko'rsatdi. Taraqqiyot nafaqat tashkilotning murakkablashishi, hayotning umumiy moslashuv zonasining kengayishi, biomassa va sonlarning o'sishi, organizmlarning avtonomiyasining oshishi, balki tirik tizimlarning barqarorligini oshirishda ham namoyon bo'ldi.
Eukariotlar misolidan foydalanib, hayotning yangi shakllarining paydo bo'lishini ba'zi bir filetik chiziqlar yoki to'qimalarning evolyutsiyasi natijasi sifatida emas, balki yuqori tartibli tizimlar rivojlanishining tabiiy va muqarrar ta'siri sifatida ko'rish kerakligi aniq ko'rsatilgan. - jamoalar, biosfera va, ehtimol, butun sayyora.
Foydalanilgan manbalar:
A. V. Markov, A. M. Kulikov. Mikroblar jamoasidagi integratsiya jarayonlari natijasida eukariotlarning kelib chiqishi
A. V. Markov. Eukariotlarning kelib chiqishi muammosi
M. V. Larina. Eukaryotik hujayralarning kelib chiqishi haqidagi farazlar. Ko'p hujayralilikning paydo bo'lishi
Dars turi - birlashtirilgan
Usullari: qisman izlash, muammoni taqdim etish, tushuntirish va illyustrativ.
Maqsad:
Talabalarda tirik tabiat, uning tizimli tashkil etilishi va evolyutsiyasi haqidagi bilimlarning yaxlit tizimini shakllantirish;
Biologik masalalar bo'yicha yangi ma'lumotlarga asosli baho berish qobiliyati;
Fuqarolik mas’uliyatini, mustaqillikni, tashabbuskorlikni tarbiyalash
Vazifalar:
Tarbiyaviy: biologik tizimlar (hujayra, organizm, tur, ekotizim) haqida; tirik tabiat haqidagi zamonaviy g'oyalarning rivojlanish tarixi; biologiya fanidagi ajoyib kashfiyotlar; dunyoning zamonaviy tabiatshunoslik manzarasini shakllantirishda biologiya fanining roli; ilmiy bilish usullari;
Rivojlanish umuminsoniy madaniyatga kirgan biologiyaning ajoyib yutuqlarini o'rganish jarayonida ijodiy qobiliyatlar; turli axborot manbalari bilan ishlash jarayonida zamonaviy ilmiy qarashlar, g‘oyalar, nazariyalar, tushunchalar, turli farazlarni (hayotning, insonning mohiyati va kelib chiqishi haqida) rivojlantirishning murakkab va qarama-qarshi yo‘llari;
Tarbiya tirik tabiatni bilish imkoniyatiga, tabiiy muhitga va o'z salomatligiga g'amxo'rlik qilish zarurligiga ishonch; biologik muammolarni muhokama qilishda raqibning fikrini hurmat qilish
Biologiyani o'rganishning shaxsiy natijalari:
1. Rossiyaning fuqarolik o'ziga xosligini tarbiyalash: vatanparvarlik, vatanga muhabbat va hurmat, o'z vatanidan faxrlanish tuyg'usi; o'z milliy mansubligini bilish; ko'p millatli rus jamiyatining gumanistik va an'anaviy qadriyatlarini o'zlashtirish; Vatan oldidagi mas’uliyat va burch tuyg‘ularini tarbiyalash;
2. ta'limga mas'uliyatli munosabatni shakllantirish, o'quvchilarning o'z-o'zini rivojlantirish va o'z-o'zini tarbiyalashga tayyorligi va qobiliyatini o'rganish va bilimga motivatsiya, ongli ravishda tanlash va dunyoda yo'naltirilganlik asosida keyingi individual ta'lim traektoriyasini qurish. barqaror kognitiv manfaatlarni hisobga olgan holda kasblar va kasbiy imtiyozlar;
Biologiyani o'qitishning meta-predmet natijalari:
1. o'z ta'lim maqsadlarini mustaqil ravishda belgilash, o'quv va kognitiv faoliyatda o'z oldiga yangi maqsadlar qo'yish va shakllantirish, o'z bilim faoliyatining motivlari va manfaatlarini rivojlantirish qobiliyati;
2. tadqiqot va loyiha faoliyatining tarkibiy qismlarini, shu jumladan muammoni ko'rish, savollar berish, farazlarni ilgari surish qobiliyatini o'zlashtirish;
3. biologik ma'lumotlarning turli manbalari bilan ishlash qobiliyati: biologik ma'lumotlarni turli manbalardan topish (darslik matni, ilmiy-ommabop adabiyotlar, biologik lug'atlar va ma'lumotnomalar), tahlil qilish va
ma'lumotlarni baholash;
Kognitiv: biologik ob'ektlar va jarayonlarning muhim belgilarini aniqlash; odamlar va sutemizuvchilar o'rtasidagi munosabatlarga dalil (argumentatsiya) taqdim etish; odamlar va atrof-muhit o'rtasidagi munosabatlar; inson salomatligining atrof-muhit holatiga bog'liqligi; atrof-muhitni muhofaza qilish zarurati; biologiya fanining usullarini o'zlashtirish: biologik ob'ektlar va jarayonlarni kuzatish va tavsiflash; biologik tajribalar tashkil etish va ularning natijalarini tushuntirish.
Normativ: maqsadlarga erishish yo'llarini, shu jumladan alternativalarni mustaqil ravishda rejalashtirish, ta'lim va kognitiv muammolarni hal qilishning eng samarali usullarini ongli ravishda tanlash qobiliyati; o'qituvchi va tengdoshlar bilan ta'lim sohasidagi hamkorlikni va birgalikdagi faoliyatni tashkil etish qobiliyati; yakka tartibda va guruhda ishlash: pozitsiyalarni muvofiqlashtirish va manfaatlarni hisobga olgan holda umumiy yechim topish va nizolarni hal qilish; axborot-kommunikatsiya texnologiyalaridan foydalanish sohasidagi kompetentsiyalarni shakllantirish va rivojlantirish (keyingi o'rinlarda AKT kompetentsiyalari deb yuritiladi).
Kommunikativ: tengdoshlari bilan muloqot va hamkorlikda kommunikativ kompetentsiyani shakllantirish, o'smirlik davrida gender sotsializatsiyasining xususiyatlarini tushunish, ijtimoiy foydali, ta'lim va tadqiqot, ijodiy va boshqa faoliyat turlari.
Texnologiyalar : Salomatlikni saqlash, muammoli, rivojlantiruvchi ta'lim, guruh faoliyati
Texnikalar: tahlil, sintez, xulosa chiqarish, axborotni bir turdan ikkinchi turga o‘tkazish, umumlashtirish.
Taqdimot hosting
"Prokariotlarning oltingugurt aylanishidagi ishtiroki"
Nijniy Novgorod 2010 yil
Kirish
Prokaryotlar (bakteriyalar va arxeyalar) Yerdagi hayot uchun alohida ahamiyatga ega - ular hayot uchun zarur bo'lgan asosiy elementlarning (uglerod, kislorod, azot, oltingugurt, fosfor) tsiklik o'zgarishida asosiy rol o'ynaydi. Tirik organizmlar tuzilgan elementlarning tsiklik o'zgarishlari moddalarning aylanishini ifodalaydi. Hozirgi vaqtda tsikllarning ma'lum bosqichlari faqat prokariotlar tomonidan amalga oshirilib, biosferadagi asosiy biogen elementlarning tsikllarining yopilishini ta'minlashi shubhasiz isbotlangan. V.I.ga ko'ra. Vernadskiy (biosfera haqidagi ta'limotning asoschilaridan biri), "biosfera hayotning qobig'ini - tirik materiyaning mavjudligi hududini ifodalaydi".
1. Oltingugurt aylanishi
Oltingugurt biogen element, tirik materiyaning zarur tarkibiy qismidir. U oqsillarda aminokislotalarning bir qismi sifatida topilgan, oqsillardagi oltingugurt miqdori 0,8-2,4% ni tashkil qiladi. Oltingugurt, shuningdek, vitaminlar, glikozidlar, koenzimlar tarkibiga kiradi va o'simlik efir moylarida mavjud. Oltingugurt yer qobig'ida, toshko'mirlarda, slanetslarda, neftlarda va tabiiy gazlarda ko'p miqdorda mavjud.
Oltingugurt o'zgaruvchan valentlikka ega elementdir. Bu uning harakatchanligini ta'minlaydi. Noorganik birikmalar shaklida oltingugurt oksidlangan shaklda (sulfatlar, politiyonatlar), qaytarilgan shaklda (sulfidlar) va molekulyar shaklda bo'lib, faol oksidlanish-qaytarilish aylanishini amalga oshiradi. Tabiatda oltingugurt turli xil kimyoviy va biologik o'zgarishlarga uchraydi, noorganik birikmalardan organiklarga va orqaga o'tadi, valentligini - 2 dan +6 gacha o'zgartiradi.
Oltingugurt birikmalarining siklik o'zgarishlariga oltingugurt aylanishi deyiladi.
Oltingugurtning konversiya aylanishiga oksidlovchi va qaytarilish birliklari, shuningdek, oltingugurtning valentligini o'zgartirmagan holda o'zgarishi kiradi. Oltingugurt aylanishining oksidlanish qismi shartlarga qarab, sof kimyoviy jihatdan ham, organizmlar, asosan mikroorganizmlar ishtirokida sodir bo'lishi mumkin bo'lgan bosqichlarni o'z ichiga oladi (bular yalang'och ko'zga ko'rinmaydigan organizmlar, shu jumladan mikroskopik eukaryotlar: zamburug'lar, suv o'tlari, protozoa va boshqalar). barcha prokaryotlar). Oltingugurt aylanishining kamaytiruvchi qismi bu jarayonda prokaryotlarning asosiy roli bilan asosan biologik tarzda amalga oshiriladi, oltingugurt atomi esa maksimal oksidlanish (+ 6) holatidan maksimal qaytarilish (-2) holatiga qaytadi. Biroq, bu jarayon har doim ham tugamaydi va atrof-muhitda to'liq oksidlanmagan mahsulotlar ko'pincha topiladi: elementar oltingugurt, politiyonatlar, sulfit.
Shunday qilib, oltingugurt aylanishi, shuningdek, moddalarning aylanishi tsiklning yopilishini ta'minlaydigan prokariotlarning ishtirokisiz mumkin emas.
2. Qayta tiklovchi filial
Sulfat assimilyatsiyasi.
Sulfat deyarli barcha o'simliklar va mikroorganizmlar tomonidan oltingugurt manbai sifatida ishlatiladi. Oltingugurtning organik birikmalarga qo'shilishi uchun assimilyatsiya paytida sulfat kamayadi, chunki tirik organizmlarda oltingugurt deyarli faqat sulfidril (-SH) yoki disulfid (-S-S-) guruhlari shaklida kamaytirilgan shaklda bo'ladi. Ikkala holatda ham organizmning o'sishi uchun zarur bo'lgan miqdorda oltingugurt o'z ichiga olgan ozuqa moddalari o'zlashtiriladi, shuning uchun oltingugurt almashinuvining qisqargan mahsulotlari atrof-muhitga chiqarilmaydi. Biosintez natijasida oltingugurt asosan oltingugurt o'z ichiga olgan aminokislotalar tarkibiga kiradi: sistin, sistein, metionin. Oltingugurt o'z ichiga olgan organik moddalar tarkibida sulfatlarning ishtirok etishi sulfatni assimilyatsiya qilish deb ataladi.
Organik oltingugurt birikmalarining H 2 S ga aylanishi.
Oltingugurt tirik organizmlar uchun asosan eruvchan sulfatlar yoki qaytarilgan organik oltingugurt birikmalari shaklida mavjud.
Oltingugurt tarkibidagi organik birikmalarning minerallashuvi jarayonida oltingugurt noorganik qaytarilgan shaklda H 2 S shaklida ajralib chiqadi. Oltingugurtni organik oltingugurt saqlovchi birikmalardan (tirik jonzotlarning metabolik mahsulotlari) ajratishda ammonifikatsiyaga qodir saprofit mikroorganizmlar ishtirok etadi. , o'lik o'simlik va hayvonlar qoldiqlari). Ammonifikatsiya jarayonida oltingugurt o'z ichiga olgan oqsillar va nuklein kislotalar parchalanib, CO 2, karbamid, organik kislotalar, aminlar va oltingugurt aylanishi uchun muhim bo'lgan H 2 S va merkaptanlar (tio spirtlari) hosil qiladi. Merkaptanlar aerob sharoitda ham oksidlanib, H2S ni chiqaradi.
Mikroorganizmlar tomonidan oqsillarni yo'q qilish hujayradan tashqari jarayon sifatida boshlanadi. Bunday holda, oqsillar protolitik ekzofermentlar yordamida hujayra ichiga kira oladigan kichikroq molekulalarga gidrolizlanadi va hujayra ichidagi proteazlar tomonidan keyingi parchalanishga olib keladigan aminokislotalarga parchalanadi.
Sulfat va elementar oltingugurtdan H 2 S ning bevosita shakllanishi.
Biosferada vodorod sulfidining paydo bo'lishi asosan oltingugurtning global aylanishi uchun katta ahamiyatga ega bo'lgan sulfat kamaytiruvchi bakteriyalarning faolligi bilan bog'liq. Sulfat kamaytiruvchi bakteriyalar dissimilyatsion sulfat qaytarilishini amalga oshiradilar, bu anaerob nafas olishdir, bunda sulfat organik moddalar yoki molekulyar vodorod oksidlanish jarayonida oxirgi elektron qabul qiluvchi (kislorod o'rniga) bo'lib xizmat qiladi. Shuning uchun sulfatni kamaytiruvchi bakteriyalarda metabolizmning energiya turi ko'pincha sulfatli nafas olish deb ataladi. Sxematik jihatdan dissimilyatsion sulfat qaytarilishida sulfatning qaytarilish jarayonini quyidagicha tasvirlash mumkin: SO 4 2- →SO 3 2- →S 3 O 6 2- →S 2 O 3 2- →S 2-.
Sulfatning qaytarilishida ishtirok etadigan enzimatik tizim ikki qismdan iborat: birinchisi, ATPga bog'liq jarayonda sulfatni sulfitga qaytaradi, ikkinchisi esa olti elektronli uzatish orqali sulfitni sulfidga qaytaradi. Aynan so'nggi reaktsiya dissimilyatsiya bo'lib, hujayrani energiya bilan ta'minlaydi.
Sulfatni kamaytiruvchi bakteriyalar asosan majburiy anaerob bakteriyalardir. Sulfatni kamaytiruvchi bakteriyalarning geokimyoviy roli juda katta, chunki ularning faolligi tufayli anaerob zonadagi inert birikma sulfat oltingugurtning biologik aylanishida keng miqyosda ishtirok etadi.
Sulfat kamaytiruvchi bakteriyalarning faolligi, ayniqsa, suv havzalari va soylar tubida, botqoqlarda va dengiz qirg'oqlarida joylashgan loyda sezilarli. Dengiz suvida sulfat kontsentratsiyasi nisbatan yuqori bo'lganligi sababli, sulfatning qaytarilishi dengiz sayozlarida organik moddalarning minerallashuvida muhim omil hisoblanadi. Bunday minerallashuv belgilari H 2 S hidi va bu jarayon sodir bo'lgan qora-qora loydir. Loyning qora rangi uning tarkibida ko'p miqdorda temir temir sulfid mavjudligi bilan bog'liq. Organik moddalarning to'planishi ayniqsa sulfatning keskin kamayishiga olib keladigan ba'zi qirg'oq hududlari H 2 S ning toksik ta'siri tufayli deyarli jonsizdir.
Sulfatni kamaytiruvchi bakteriyalar sistematik emas, fiziologik guruhdir, chunki ular bir fiziologik jarayonni - sulfatlar ishtirokida anaerob nafas olishni, masalan, Desulfovibrio (vibrion) avlodining bakteriyalarini amalga oshirishga qodir bo'lgan turli taksonomik guruhlarga mansub bakteriyalarni o'z ichiga oladi. Desulfotomakulum (spora hosil qiluvchi tayoqchalar). Arxeyalar sulfat kamaytiruvchi organizmlar orasida ham topilgan. Sulfat kamaytiruvchi bakteriyalar nafaqat sulfatlarni, balki tiosulfat, sulfit, elementar oltingugurt va boshqa oltingugurt birikmalarini ham kamaytirish orqali ko'payishi mumkin.
Ba'zi sulfat kamaytiruvchi bakteriyalar uchun oltingugurt almashinuvining asosan yangi turi ko'rsatildi. Bu bakteriyalar organik substratlarda o'sganda nafaqat sulfatning kamayishi, balki tiosulfat, sulfit, ditionitning sulfat va sulfid hosil bo'lishi bilan nomutanosibligi natijasida ham energiya olishga qodir.
Umumiy tiosulfat nomutanosiblik reaktsiyasi:
S 2 O 3 2- + H 2 0 → SO 4 2- + HS - + H +
Go = – 21,9 kJ/mol S 2 O 3 2-
Umumiy sulfit nomutanosiblik reaktsiyasi:
4SO 3 2- + H + → 3SO 4 2- + HS -
Go = – 58,9 kJ/molSO 3 2-
Dengiz cho'kindilarida tiosulfatni nomutanosib bo'lgan bakteriyalar keng tarqalganligi aniqlandi.
Vodorod sulfidi elementar oltingugurtni kamaytirish jarayonida ham hosil bo'lishi mumkin. Bugungi kunda molekulyar oltingugurtdan vodorod sulfidini hosil qilishning ikkita mexanizmi ma'lum. Birinchi holda, bakteriyalar va arxeylar molekulyar oltingugurtdan anaerob nafas olish (dissimilyatsiya qiluvchi oltingugurtni kamaytirish) jarayonida elektron qabul qiluvchi sifatida foydalanadilar, bunda ATP sintezlanadi. Dissimilyatsion oltingugurtni kamaytirish - bu mezofil va termofil prokaryotlar tomonidan amalga oshiriladigan fermentativ jarayon. Ikkinchi holda, mikroorganizmlar (xamirturush va prokaryotlar) oltingugurtdan faqat fermentatsiya paytida chiqarilgan elektronlarni tashlash uchun foydalanadilar (engillashtirilgan fermentatsiya). Bu ATP sintezi bilan birga bo'lmagan elektronlarning befoyda (bo'sh) zaryadsizlanishi.
Molekulyar oltingugurtning vodorod sulfidiga aylanishi, ayniqsa, elementar oltingugurt oltingugurtning eng muhim shakllaridan biri bo'lgan va oltingugurt birikmalari dastlab vulqon kelib chiqishi bo'lgan gidrotermlarda yashovchi termofil mikroorganizmlar uchun katta ahamiyatga ega. Gidrotermik biotsenozlar tirik mavjudotlarning noyob jamoalaridir. Yuqori haroratlarda (45-50 dan 100 ° C gacha) rivojlanib, ular asosan prokaryotlar - bakteriyalar va arxeyalardan hosil bo'ladi. Gidrotermal suyuqliklarning mikrobial jamoalarini tashkil etuvchi mikroorganizmlarning katta qismi boshqa joylarda topilmaydi. Gidrotermlarning mikrobial jamoalari Yerdagi eng qadimgi biotsenozlar hisoblanadi.
3. Oksidlanish shoxchasi
Oltingugurt aylanishining bu qismi butunlay noorganik oltingugurt birikmalarining reaktsiyalaridan iborat bo'lishi mumkin: S 2- → nS 2- → S 0 → S 2 O 3 2- → SO 3 2- → SO 4 2-, shuningdek, organik reaktsiyalarni ham o'z ichiga oladi. shakllari. Organik sulfidlarning oltingugurt atomi odatda S2 shaklida ajratilgandan so'ng - noorganik yo'l orqali oksidlanadi, ammo sof organik oksidlanish yo'li ham mumkin bo'lsa-da, oltingugurt atomi organik birikmalarning bir qismi sifatida oksidlanganda, masalan, sistein → sistin. Oltingugurt birikmalarining aksariyat oksidlanish reaktsiyalari kuchli oksidlovchi moddalar (H 2 O 2, O 3, kislorod radikallari) ishtirokida mikroorganizmlar ishtirokisiz sodir bo'lishi mumkin, ammo mikrobial oksidlanish, ayniqsa reagentning past konsentratsiyasida ancha samarali.
Oltingugurt birikmalarini oksidlovchi mikroorganizmlar orasida uchta asosiy fiziologik guruh ajratiladi: anaerob fototrof bakteriyalar, aerob va fakultativ anaerob litotrof bakteriyalar, arxeya va turli xil geterotrof mikroorganizmlar.
Anaerob fototrof bakteriyalar.
Bu o'simliklar va siyanobakteriyalardagi aerob (kislorodli) fotosintezda bo'lgani kabi H 2 O o'rniga elektron donorlar sifatida turli xil qaytarilgan oltingugurt birikmalaridan foydalanib, anaerob (anoksigen) fotosintezni amalga oshiradigan bakteriyalarning o'ziga xos guruhidir. Qaytarilgan oltingugurt birikmalarining eng keng tarqalgan oksidlanish mahsulotlari SO 4 2- va S 0 dir, ikkinchisi ba'zi fototrofik bakteriyalarda hujayra ichida to'planishi mumkin.
Tizimli ravishda fototrof bakteriyalar bir necha guruhlarga bo'linadi: oltingugurtli va oltingugurtsiz binafsha va yashil bakteriyalar, geliobakteriyalar, eritrobakteriyalar (123-rasm). Fototrof bakteriyalar pigmentli bo'lib, jigarrang, yashil yoki binafsha rangga ega bo'lishi mumkin. Anoksigenli fotosintezni amalga oshiradigan fototrof bakteriyalarda bakterioxlorofil va karotinoidlar mavjud bo'lib, ular ularga mos rang beradi. Bu bakteriyalar ko'llar, hovuzlar, lagunalar va mineral buloqlar kabi sayoz suv havzalarining anaerob suv ustunida sulfid oksidlanishida ustunlik qiladi, bu erda etarli yorug'lik kiradi. Istisno - bu majburiy aeroblar bo'lgan va aerob sharoitda anoksigenli fotosintezni amalga oshiradigan eritrobakteriyalar.

Litotrofik oltingugurt oksidlovchi bakteriyalar va arxeya. Oltingugurt bakteriyalari va tion bakteriyalari.
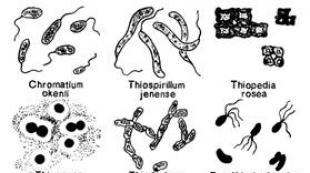
Ba'zi prokaryotlar oltingugurt birikmalari kamaygan holda litotrofik o'sishga qodir. Litotrofik o'sish jarayonida qaytarilgan oltingugurt birikmalari yoki boshqa qaytarilgan noorganik birikmalar (NH 3, H 2, CO, Fe 2+) energiya almashinuvida elektron donorlardir, ya'ni ular elektron tashish zanjirida oksidlanganda ATP sintezlanadi. Faqat prokaryotlar litotrofik o'sishga qodir. Litotrofik oltingugurtga bog'liq prokariotlarda kamaytirilgan oltingugurt birikmalarining oksidlanish reaktsiyalari energiya manbai bo'lib xizmat qiladi. Oksidlanish reaktsiyalari maxsus ferment tizimlari ishtirokida amalga oshiriladi. Har xil oltingugurt birikmalarining va oltingugurtning litotrofik prokaryotlar tomonidan oksidlanishi odatda sulfatlarning hosil bo'lishiga olib keladi. Lekin bu jarayon har doim ham tugallanavermaydi va toʻliq oksidlanmagan mahsulotlar (S 0, SO 3 2-, S 2 O 3 2-, S 4 O 6 2-) koʻpincha muhitda uchraydi. Litotrofik oltingugurtga bog'liq prokaryotlar guruhiga asosan tion bakteriyalari, oltingugurtga juda termoatsidofil qaram arxeyalar, vodorod bakteriyalari, rangsiz oltingugurt bakteriyalari, shuningdek oltingugurt birikmalarining oksidlanishi tufayli litotrofik o'sishga qodir binafsha rangli bakteriyalarning ba'zi vakillari kiradi. qorong'i.
Vodorod sulfidli buloqlarda va vodorod sulfidi bo'lgan boshqa suv havzalarida, qoida tariqasida, ko'p miqdorda rangsiz mikroorganizmlar topilganligi uzoq vaqtdan beri ma'lum bo'lib, ularning hujayralarida oltingugurt tomchilari topiladi. Vodorod sulfidining kontsentratsiyasi nisbatan past bo'lgan joylarda (50 mg / l dan kam) rangsiz oltingugurt bakteriyalari deb ataladigan bunday mikroorganizmlar ko'pincha plyonkalar, oq qatlamlar va boshqa ifloslanishlar shaklida ommaviy to'planishlarni hosil qiladi. S.N. Winogradskiy (1887) oltingugurt bakteriyalarining tipik vakillaridan biri, ya'ni Beggiatoa hujayralarida to'plangan oltingugurt vodorod sulfididan hosil bo'lishini va bu mikroorganizm tomonidan sulfat kislotaga oksidlanishi mumkinligini isbotladi. Tizimli ravishda, rangsiz oltingugurt bakteriyalari, ko'rinishidan, heterojen guruh bo'lib, tavsiflangan barcha turlar va hatto avlodlar ham mustahkam o'rnatilgan deb hisoblanishi mumkin emas. Morfologiyasi, harakatlanish tabiati, koʻpayish usuli va hujayra tuzilishi jihatidan koʻp hujayrali va bir hujayrali (Beggiatoa, Thiothrix, Thiospirillopsis, Thioploca, Ahromatium) rangsiz oltingugurtli bakteriyalarning bir qator vakillari koʻk-yashil suvoʻtlar bilan katta oʻxshashlik koʻrsatadi. Ba'zi tadqiqotchilar, xususan, Pringsheim (1963) bu mikroorganizmlarni ularning rangsiz variantlari deb hisoblashadi. Ko'k-yashil suv o'tlari Oscillatoria, Thiothrix - Rivularia, Thiospirillopsis - Spirulina Beggiatoa analoglari hisoblanadi va Ahromatium Synechococcus ga o'xshaydi. Ko'k-yashil suv o'tlari endi bakteriyalar deb tasniflanganligi sababli, ularning rangsiz oltingugurt bakteriyalari bilan aloqasi tobora oqlanib bormoqda. Shuni ham ta'kidlash kerakki, ba'zi ko'k-yashil suvo'tlar o'z hujayralarida oltingugurtni to'plashi aniqlangan, garchi bu xususiyatning o'zi mikroorganizmlar taksonomiyasini ko'p ta'minlamaydi. Rangsiz oltingugurt bakteriyalariga mansub mikroorganizmlar ham chuchuk, ham sho'r suv havzalarida uchraydi. Ulardan ba'zilari past haroratlarda yaxshi o'sadi, boshqalari (Thiospirillum pistiense) 50 ° C dan yuqori haroratda termal oltingugurtli buloqlarda rivojlanadi. Mobil shakllar kemotaksisga ega va optimal kislorod va vodorod sulfidi tarkibiga ega joylarga o'tishi mumkin.
Tion deb ataladigan bakteriyalarning ko'pchiligi tipik kimyoavtotroflardir, ya'ni. ular kamaytirilgan oltingugurt birikmalaridan nafaqat H-donorlari, balki energiya manbalari sifatida ham foydalanadilar va karbonat angidridni assimilyatsiya qilib, sof mineral muhitda o'sishga qodir. Bunday bakteriyalar birinchi marta Neapol ko'rfazining suvidan ajratilgan (Natanson, 1902) va Thiobacillus (Beijerinck, 1904) nomini oldi. Bugungi kunga qadar turli xil suv havzalaridan, tuproqdan, shuningdek oltingugurt va turli metallar konlaridan ajratilgan avtotrof tiobaksillalarning ko'p turlari tasvirlangan.
Ba'zi, asosan fiziologik xususiyatlarga (turli xil oltingugurt birikmalarini va boshqa noorganik substratlarni oksidlash qobiliyati, organik birikmalardan foydalanish, atrof-muhitning pH qiymatiga, kislorodga munosabatiga va boshqalarga qarab o'sish qobiliyati) ko'p sonli tiobaksilluslar asosida. turlari ajralib turadi. Ulardan quyidagilar eng mustahkam oʻrnashganlari hisoblanadi: Thiobacillus thioparus, T. thiooxidans, T. denitrificans, T. thiocyanoxidans, T. neapolitanus, T. intermedius, T. novellus, T. ferrooxidanas. Bu mikroorganizmlarning barchasi oddiy mineral muhitda o'sadi, bu erda ko'pincha bikarbonat qo'shiladi. Azot manbai odatda ammoniy tuzlari hisoblanadi. Ba'zi turlar va shtammlar nitratlarni azot manbalari sifatida o'zlashtira oladi. Ayrim vakillarning karbamid va aminokislotalardan foydalanish qobiliyati ham ko'rsatilgan. Tuzli suv havzalaridan ajratilgan shtammlar o'sishi uchun patrium xloridni talab qiladi va to'yingan NaCl eritmasida o'sishi mumkin bo'lgan galofillar ma'lum. Thiobacillus jinsining ko'pgina izolyatsiya qilingan mikroorganizmlarining o'sishi uchun optimal harorat taxminan 30 ° C dir. Biroq, tabiiy sharoitda ular 55 ° S gacha bo'lgan haroratda termal buloqlarda topiladi. Yaqinda Thiobacillus sp.ning sof madaniyati ajratildi. 50 ° C da o'sishi uchun optimal bilan.
Atrof muhitning kislotaligiga nisbatan tiobaksillar neytral yoki hatto gidroksidi reaktsiyada yaxshi o'sadigan organizmlarga va atsidofil turlarga bo'linadi, ya'ni. kislotali muhitda rivojlanadi va juda past pH qiymatlariga bardosh beradi.
Birinchi guruhga: T. thioparus, T. denitrificans, T. novellus, T. thiocyanoxidans, T. neapolitanus kabi turlar kiradi. Ushbu mikroorganizmlar uchun optimal pH qiymati 6,0-9,0 mintaqasida va ularning o'sishi mumkin bo'lgan pH qiymatlari zonasi 3,0-6,0 dan 10,0-11,0 gacha, turli turlar va shtammlar uchun optimal pH qiymatlari. va. ularning o'sishi kuzatiladigan faol kislotalilik mintaqasi sezilarli darajada farq qilishi mumkin.
Ikkinchi guruhga T. thiooxidans, T. ferroxidans, T. intermedius kiradi. Ushbu mikroorganizmlar uchun optimal pH qiymati 2,0-4,0 ni tashkil qiladi va o'sish 0,5-2,0 dan 5,0-7,0 gacha bo'lgan pH darajasida mumkin. Birinchi ikki tur eng atsidofil organizmlardir. Ushbu bakteriyalarning ikkalasi ham 5,0 dan oshmaydigan pH qiymatlarida o'sadi. Shu bilan birga, T. thiooxidans 1,0 n ga to'g'ri keladigan 0 ga yaqin pH qiymatida hayotiyligini ko'rsatdi. sulfat kislota eritmasi. Bu tadqiqotchilarga ma'lum bo'lgan eng atsidofil mikroorganizmdir.
Ko'pgina tionik bakteriyalar faqat kislorod ishtirokida o'sadi, garchi ba'zi vakillarining o'sishi past kislorod darajasida mumkin. Ammo fakultativ anaeroblar ham ma'lum. Bularga T. denitrifikans kiradi. Aerob sharoitda bu bakteriyalar molekulyar kislorod ishtirokida oksidlanish jarayonlarini amalga oshiradi, anaerob sharoitda ular denitrifikatsiyaga o'tadi va nitratlarni molekulyar azotga kamaytiradi. Shunisi qiziqki, T. denitrificans, xuddi Paracoccus denitrificans kabi, azot manbai sifatida nitratlarni o'zlashtira olmaydi va o'sishi uchun azot manbai sifatida ammoniyni talab qiladi.
Tionik bakteriyalar vodorod sulfidi, sulfidlar, sulfit, tiosulfat, tetrationat, tiosiyanat (rodanid), ditionit kabi oltingugurt birikmalarini, shuningdek, molekulyar oltingugurtni to'liq oksidlanishda sulfatlar hosil qilish qobiliyatiga ega. Biroq, alohida turlarning qobiliyatlari butunlay bir xil emas. Bundan tashqari, qaysi oltingugurt birikmalarining biologik oksidlanishini aniqlash har doim ham oson emas, chunki ularning ko'plari past pHda beqaror va atmosfera kislorodi bilan ham oksidlanishi mumkin.
Neytral va ishqoriy muhitda o'sadigan barcha turlar, qoida tariqasida, vodorod sulfidi, oltingugurt va tiosulfatni oksidlaydi. T. thioyanoxidansning xarakterli xususiyati tiosiyanidni oksidlash qobiliyatidir. Shu asosda u alohida tur sifatida ajralib turadi, chunki boshqa xususiyatlarda bu mikroorganizm T. thioparusga o'xshaydi. T. thiooxidans kabi atsidofil bakteriya odatda molekulyar oltingugurt bo'lgan muhitda etishtiriladi. Ushbu turning vodorod sulfidi va boshqa oltingugurt birikmalarini oksidlash qobiliyati haqidagi savol to'liq hal etilmagan, chunki bu birikmalar kislotali sharoitda beqaror. T. ferroksidanlarga kelsak, bu bakteriyalar molekulyar oltingugurtni ham, uning turli birikmalarini ham, xususan: vodorod sulfidi, tiosulfat, ditionit, tetrasulfat, sulfitni oksidlashi mumkinligi haqida dalillar mavjud. Bundan tashqari, T. ferroksidans suvda erimaydigan ogʻir metallar sulfidlarining oksidlanishida faol ishtirok etadi. Bularga pirit (FeS2), xalkopirit (CuFeS2), stibnit (SbS2), xalkotsit (Cu2S), kavellit (CuS), pirrotit (FeS), realgar (AsS), violarit (Ni2FeS4) va boshqalar kabi minerallar kiradi. Qolganlari. bakteriyalar yoki bu qobiliyatga ega emas, yoki u zaif ifodalangan.
T. ferroksidanlarning oʻziga xos xususiyati, shuningdek, temir temirni oksidli temirga oksidlash qobiliyatidir. Shu asosda, bu bakteriyalarni temir bakteriyalar deb hisoblash mumkin, garchi ular Thiobacillus jinsiga tegishli.
Yuqorida ta'kidlanganidek, molekulyar oltingugurt va uning turli birikmalarini tion bakteriyalari oksidlanishining yakuniy mahsuloti sulfatdir. Agar jarayon shu tarzda davom etsa, ya'ni. asl substratning to'liq oksidlanishi sodir bo'ladi, keyin uning natijalari quyidagi tenglamalarda aks ettiriladi. Vodorod sulfidining oksidlanishida:
Aerob sharoitda oltingugurt va tiosulfat oksidlanishida:

Oltingugurt va tiosulfatni nitratlar yordamida anaerob sharoitda T. denitrificans oksidlanishida:

T. thiocyanoxidans thioanide oksidlanishida:

Biroq, oksidlanish ko'pincha tugamaydi va muhitda turli xil to'liq oksidlangan mahsulotlar topiladi. Shunday qilib, vodorod sulfidining oksidlanishida ba'zida molekulyar oltingugurt paydo bo'ladi, tiosulfat va politiyonatlar ham topiladi. Molekulyar oltingugurtning bakteriyalar tomonidan oksidlanishi jarayonida tiosulfat va politiyonatlarning paydo bo'lishi kuzatildi. Tiosulfatning oksidlanishi, shuningdek, ko'pincha politiyonatlar (tritionat, tetrationat, peitatiopatat) va elementar oltingugurt hosil bo'lishi bilan birga keladi. Sulfit ham to'liq oksidlanmagan mahsulotlardan biridir. Ammo bu birikmalarning barchasi fermentativ jarayonlarning natijasi bo'lishi mumkin emas va aslida asl substratning bakterial oksidlanishining oraliq mahsulotlariga tegishli. Ulardan ba'zilari aniq kimyoviy yoki yon biologik reaktsiyalar natijasida hosil bo'lgan. Shuning uchun turli xil oltingugurt birikmalarining mikroorganizmlar tomonidan oksidlanish yo'llarini tushuntirish juda qiyin va barcha reaktsiyalar hali aniqlanmagan.
Sulfid o'zgarishining birinchi bosqichining tabiati haqida hali juda kam ma'lumotlar mavjud va oltingugurt uning oksidlanishining bevosita yoki qo'shimcha mahsuloti ekanligi to'liq aniq emas.
Bakteriyalar tomonidan elementar oltingugurtdan foydalanish masalasi juda murakkab va u ikki jihatga ega: mikroorganizmlar ushbu suvda erimaydigan moddaga qanday ta'sir qiladi va oksidlanish jarayoni qanday sodir bo'ladi.
Tiobaksillusning oltingugurtga ta'sir qilish mexanizmiga nisbatan ikkita nuqtai nazar mavjud.
1. Oltingugurt bakteriyalar tomonidan oksidlanishi uchun u hujayralar bilan bevosita aloqada bo'lishi kerak.
2. Oltingugurt bakteriyalar tomonidan atrof muhitga chiqaradigan lipid tabiatli moddalarda oldindan eritilgandan so'ng ishlatiladi.
T. thiooxidans molekulyar oltingugurtli muhitda oʻsganda, muhitda fosfolipidlar topiladi. Ba'zi ma'lumotlarga ko'ra, bu fosfatidilinositol, boshqalarga ko'ra, u bir oz farqli birikmalar (fosfatidil-N-metiletanol, fosfatidilgliserol, difosfatidilgliserin) va ularning to'planishi ekinlarning faol o'sishi bosqichiga to'g'ri keladi. Biroq, ikkinchi taxminni isbotlangan deb hisoblash mumkin emas. Ehtimol, oltingugurtning bakteriyalar tomonidan oksidlanishi uchun uning hujayralar bilan aloqasi ham, oltingugurtni "ho'llaydigan" ba'zi moddalarning chiqishi ham muhimdir.
Oltingugurt oksidlanish yo'liga kelsak, ma'lumotlar ham bir-biriga ziddir. Quyidagi sxema vodorod sulfidining oksidlanishi uchun ham maqbul bo'lgan eng ehtimolli hisoblanadi:

X vodorod sulfidi yoki oltingugurt reaksiyaga kirishadigan glutation hosilasi yoki hujayra membranalari bilan bog'langan tiol deb taxmin qilinadi. Tiosulfat hosil bo'lishi bakteriyalar ishtirokisiz sodir bo'lishi mumkin bo'lgan fermentativ bo'lmagan jarayon hisoblanadi.
Tiosulfat oksidlanishini o'rganishga ko'plab tadqiqotlar bag'ishlangan. Butun hujayralar va turli xil turdagi tiobaksillularning hujayrasiz preparatlari bilan o'tkazilgan tajribalar ma'lumotlari shuni ko'rsatadiki, tiosulfat konversiyasining dastlabki bosqichi uning reduktaza ta'sirida vodorod sulfidi va sulfit hosil bo'lishi bilan kamayishi bilan bog'liq bo'lishi mumkin:

Yoki elementar oltingugurt va sulfitga bo'linish bilan:

yoki, nihoyat, tetrationatga oksidlanish va keyin tritionat va sulfitga aylantirish bilan:

Biroq, tiosulfatni konversiyalashning oxirgi yo'li asosiy emas deb taxmin qilinadi. Tiosulfatning sulfit hosil qilish uchun parchalanishiga kelsak, bunday reaktsiya oltingugurtni tiosiyanat shaklida bog'laydigan rodonaz tomonidan amalga oshirilishi mumkin:
U nafaqat tion va boshqa bakteriyalarda, balki hayvonlarda ham keng tarqalgan. Biroq, bu ferment tionik bakteriyalar tomonidan tiosulfat metabolizmida ishtirok etadimi yoki yo'qmi, aniq aniqlanmagan.
Sulfit oksidlanish yo'li ancha o'rganilgan. Pek laboratoriyasida T. thioparus bilan olib borilgan ishlar shuni ko'rsatdiki, sulfit adenozin monofosfat (AMP) bilan reaksiyaga kirishadi, natijada adenil sulfat yoki adenozin fosfosulfat (APS) deb ataladigan birikma hosil bo'ladi. Keyingi bosqichda API va noorganik fosfat (Pn) o'rtasida adenozin difosfat (ADP) hosil bo'lishi va erkin sulfatning chiqishi bilan reaktsiya sodir bo'ladi:

Adenilatkinaz fermenti ta'siri natijasida ikkita ADP molekulasi ATP ga aylanishi va yana AMP hosil qilishi mumkin:
Shunday qilib, sulfit oksidlanishining bu yo'li substrat fosforlanishi deb ataladigan natijada energiya (ATP) ishlab chiqarish bilan bog'liq. Shu bilan birga, sulfit oksidlanish jarayoni elektronlarning nafas olish zanjiriga o'tkazilishi bilan birga bo'lishi mumkin, ularning ishlashi ATP sintezi bilan bog'liq.
Bir qator tionli bakteriyalar, shu jumladan T. thioparus uchun elektronlarning nafas olish zanjiriga oʻtishini taʼminlovchi sulfit oksidlovchi ferment taʼsiri natijasida APS hosil boʻlmasdan sulfit oksidlanishi mumkinligi koʻrsatilgan. sitoxromlar darajasida:
![]()
Ehtimol, bir xil organizmda sulfit va boshqa oltingugurt birikmalarining oksidlanish yo'llari turlicha bo'lishi mumkin va u yoki boshqasining ahamiyati atrof-muhit sharoitlariga va boshqa omillarga bog'liq. Hozirgi mavjud ma'lumotlarga asoslanib, turli xil oltingugurt birikmalarining tionik bakteriyalar tomonidan oksidlanishini quyidagi umumlashtirilgan sxema bilan ifodalash mumkin.

Tionik bakteriyalarning elektron transport tizimlarining tarkibiy qismlariga kelsak, barcha ma'lumotlarga ko'ra, ularning turli xil turlarida tarkibi doimo c tipidagi sitoxromlarni o'z ichiga oladi. b tipidagi sitoxromlar va, aftidan, uning sitoxrom o deb ataladigan varianti, ba'zi vakillarida esa a va d sitoxromlari ham topilgan. Bundan tashqari, elektron tashish tizimiga flavoproteinlar va ubikinonlar kiradi. Ammo nitrifikator bakteriyalar kabi elektronlarning kislorodga o'tishini ta'minlaydigan tion bakteriyalarining nafas olish zanjiri nisbatan qisqa, chunki oksidlangan substratlar ancha yuqori oksidlanish-qaytarilish potentsialiga ega. Shuning uchun, bu mikroorganizmlar uchun termodinamik gradientga (teskari elektron uzatish) qarshi energiyaga bog'liq bo'lgan elektron uzatish muhim ahamiyatga ega. U karbonat angidrid va boshqa konstruktiv jarayonlarni assimilyatsiya qilish uchun zarur bo'lgan kamaytirilgan NAD hosil bo'lishini ta'minlaydi. Shuni ham ta'kidlash kerakki, elektron transport tizimlari va elektron uzatish yo'llarining tarkibiy qismlari nafaqat tiop bakteriyalarining turli vakillarida aniq farqlarga ega bo'lishi mumkin, balki oksidlangan substratning tabiatiga ham bog'liq. Shuning uchun umumlashtirilgan diagramma (140-rasm) juda shartli.

Turli mualliflarning hisob-kitoblariga ko'ra, tionik bakteriyalar tomonidan erkin energiyadan foydalanish samaradorligi 2 dan 37% gacha. Boshqa kimyoavtotroflar singari, tionli bakteriyalar karbonat angidridni asosan Kalvin sikli orqali o'zlashtiradi. Ammo ular ma'lum metabolitlarning shakllanishi uchun muhim bo'lgan boshqa karboksillanish reaktsiyalarini ham amalga oshirish qobiliyatiga ega.
Oltingugurtga haddan tashqari termoatsidofil bog'liq bo'lgan arxeyalar, shuningdek, elementar oltingugurt ishtirokida litotrofik o'sishga qodir va 40-100 ° C va pH 1-6 da rivojlanishi mumkin; bu gidrotermlar va vulqon kalderalarining aholisi.
Geterotrof oltingugurt oksidlovchi mikroorganizmlar (oziqlanish uchun organik moddalardan foydalanadigan organizmlar).
Ba'zi majburiy geterotrof bakteriyalar, xamirturushlar va mikromitsetlarning turli noorganik oltingugurt birikmalarini oksidlash qobiliyati ancha vaqtdan beri ma'lum bo'lib kelgan, garchi bu reaktsiyalarning geterotroflarning metabolizmidagi ahamiyati hali ham to'liq tushunilmagan. Geterotrof oltingugurt oksidlovchi bakteriyalar orasida tiosulfatning tetrationatgacha to'liq bo'lmagan oksidlanishini amalga oshiradigan psevdomonadlar guruhining turlari ustunlik qiladi.
Shuningdek, S.N. Winogradskiy (1887-1889) rangsiz oltingugurt bakteriyalari juda oz miqdorda organik moddalar bo'lgan suvda o'sishi mumkinligini ta'kidladi va shuning uchun ular karbonat angidridni o'zlashtira oladi deb taxmin qildilar. Beggiatoaning turli shtammlari fiziologiyasini o'rganishga asoslanib, Pringsheim ular orasida vodorod sulfidini oksidlovchi va CO 2 ni mahkamlaydigan avtotroflar va organik birikmalarni talab qiladigan vakillar borligiga ishonadi. Ammo bir qator geterotrofik Beggiatoa shtammlari ham organik birikmalar ishtirokida vodorod sulfidini oksidlaydi, ehtimol energiya ishlab chiqaradi, ya'ni. ular xemolitoheterotroflardir. Rangsiz oltingugurt bakteriyalari bilan bir qatorda vodorod sulfidi, molekulyar oltingugurt va tiosulfat oksidlanishida ishtirok etadigan tipik geterotrof mikroorganizmlar ham ma'lum. Bularga Bacillus, Pseudomonas, Achromobacter, Sphaerotilus vakillari, shuningdek aktinomitsetalar, mog'or (Penicillium luteum, Aspergillus niger), xamirturush va Alternaria kiradi. Ulardan ba'zilari, xususan, filamentli ko'p hujayrali bakteriya Sphaerotilus natans vodorod sulfid ishtirokida oltingugurtni hujayralarga joylashtiradi. Boshqalar (Pseudomonas aeruginosa, Ps. fluorescens, Achromobacter stuzeri) tiosulfatni tetrationat (Na2S4O6) ga oksidlanishga qodir. Geterotrof mikroorganizmlarning aralash kulturalari elementar oltingugurt bilan reaksiyaga kirishganda politiyonatlar va sulfat hosil bo'lishi ham qayd etilgan. Oksidlanish mexanizmi va bu jarayonning geterotroflar uchun biologik ahamiyati noaniqligicha qolmoqda. Ba'zi geterotrof rangsiz oltingugurt bakteriyalari oltingugurt birikmalarini vodorod periks bilan oksidlash qobiliyatiga ega, organik moddalarning oksidlanishi paytida hujayralarda hosil bo'lgan superoksid radikali: H 2 O 2 + H 2 S → S 0 + 2H 2 O. Bu jarayonning fiziologik ma'nosi. kislorodni to'liq bo'lmagan kamaytirishning toksik mahsulotlarini zararsizlantirishdir (H 2 O 2, O 2-).
Oltingugurt birikmalarining sulfatgacha toʻliq oksidlanishi elementar oltingugurt bilan boyitilgan tuproqlarda yashovchi mikromitsetlar uchun koʻproq xosdir. Aspergillus, Penicillium, Trichoderma, Fusarium, Mucor va Auerobasidium mikroskopik zamburug'larining ba'zi turlari uchun elementar oltingugurt, tiosulfat va hatto metall sulfidlarni sulfatga oksidlash qobiliyati ko'rsatilgan, ammo bunday oksidlanish tezligi bir yoki ikkita tartibni tashkil qiladi. litotrof bakteriyalarnikidan pastroqdir.
oltingugurtning siklik transformatsiyasi mikroorganizmi
Xulosa
So'nggi o'n yilliklarda oltingugurtning tabiiy aylanishi ortib borayotgan antropogen ta'sirga duchor bo'lib, toksik oltingugurt birikmalarining to'planishiga va tabiiy oltingugurt aylanishining muvozanatiga olib keldi. Xususan, oltingugurt birikmalarining katta hajmdagi chiqindilari natijasida issiqlik elektr stansiyalari tomonidan qazib olinadigan yoqilg‘i, vodorod sulfidi va sellyuloza-qog‘oz va metallurgiya korxonalari tomonidan ajralib chiqadigan uchuvchan organik sulfidlar, shuningdek, kommunal xomashyoning parchalanishi paytida ajralib chiqadigan oltingugurt dioksidi hosil bo‘ladi. va qishloq xo'jaligi oqava suvlari. Bu birikmalar mikrogram konsentrasiyalarda ham zaharli hisoblanadi. Ular havoni zaharlashi, atmosfera kimyosiga ta'sir qilishi va suvda erigan kislorod tanqisligiga olib kelishi mumkin.
Shuning uchun oltingugurt birikmalarini o'zgartirish va toksik oltingugurt birikmalarini zararsizlantirishda ishtirok etadigan prokaryotlardan foydalanish juda dolzarb ko'rinadi. Bundan tashqari, mikroorganizmlarning juda yuqori katalitik faolligi ularning Yer yuzasida sodir bo'ladigan kimyoviy o'zgarishlarda katta rol o'ynashiga sabab bo'ladi. Mikroorganizmlar kichik o'lchamlari tufayli hayvonlar va yuqori o'simliklar bilan solishtirganda sirt-hajm nisbati yuqori bo'lib, hujayra va atrof-muhit o'rtasida substratlar va ajralib chiqadigan mahsulotlarning tez almashinuviga olib keladi.
Ularning qulay sharoitlarda ko'payishi va biosferada keng tarqalganligi ham muhim omillardir. Shu bilan birga, prokariotlarning ozuqa moddalarining o'zgarishidagi alohida ahamiyatiga qaramay, moddalar aylanishidagi, xususan, oltingugurt aylanishidagi prokariotik faollik ko'lami hali to'liq baholanmagan.
Adabiyot
1. Grabovich M.Yu. Prokaryotlarning oltingugurt aylanishida ishtiroki // Soros Educational Journal, 1999, No 12, p. 16–20.
2. Gromov B.V., Pavlenko G.V. Bakteriyalar ekologiyasi. L.: Leningrad davlat universiteti nashriyoti, 1989. 248 b.
3. Gromov B.V. Arxeyaning ajoyib dunyosi // Soros ta'lim jurnali. 1997 yil. 4-son. 23–26-betlar.
4. Kaldera mikroorganizmlari / Ed. G.A. Zavarzina. M.: Nauka, 1989. 120 b.
5. Kondratyeva E.N. Xemolitotroflar va metilotroflar. M.: Moskva davlat universiteti nashriyoti, 1983. 172 b.
6. Malaxov V.V. Vestimentifera avtotrof hayvonlardir // Soros Educational Journal. 1997 yil. 9-son. 18–26-betlar.
Sayyora biosferasi yagona megaorganizm bo'lib, uning qismlari bir-biri bilan uyg'un bog'langan. Hayotning butun xilma-xilligi ikkita super shohlikka bo'lingan - prokaryotlar (yadrodan oldingi organizmlar) va eukaryotlar (yadroga ega). Haddan tashqari qirolliklar tirik shohliklarga bo'lingan:
- viruslar;
- bakteriyalar;
- qo'ziqorinlar;
- o'simliklar;
- hayvonlar.
Bakteriyalar shohligi viruslar shohligi bilan birga prokariotlarning super qirolligiga - yadrosiz organizmlarga birlashtirilgan. Tarixiy jihatdan, bu sayyoradagi tirik organizmlarning birinchi eshelonidir.
Bir hujayrali tirik organizmlar taxminan 3,8 milliard yil oldin paydo bo'lgan. Deyarli bir milliard yil davomida ular sayyoramizning yagona tirik aholisi bo'lgan - ular muvaffaqiyatli ko'paygan, rivojlangan va moslashgan. Ularning hayotiy faoliyati natijasi sayyora atmosferasida erkin kislorodning paydo bo'lishi edi, bu ko'p hujayrali organizmlar - zamburug'lar, o'simliklar, gullar va hayvonlarning paydo bo'lishiga imkon berdi.
Hozirgi vaqtda tirik bakteriyalar hamma joyda yashaydi: siyrak atmosfera qatlamlaridan tortib eng chuqur okean tublarigacha, ular Arktika muzlarida va termal geyzerlarda yashaydi. Bakteriyalar nafaqat bo'sh joyni mustamlaka qildilar - ular qo'ziqorinlar, o'simliklar yoki hayvonlar bo'lsin, boshqa organizmlarning ichida ko'payadi.

Tabiatda barcha hayvonlar va odamlar bundan mustasno emas, ular yashaydigan mikroblar uchun yashash joylari hisoblanadi:
- teri;
- og'iz bo'shlig'i;
- ichaklar.
Olimlar inson organizmida yashovchi mikroorganizm hujayralari soni o‘z hujayralari sonidan 10 barobar ko‘p ekanligini aniqladilar. Bunday yuqori miqdoriy ko'rsatkichlarga qaramay, organizmda yashovchi bakteriyalarning og'irligi 2 kg dan oshmaydi - bu hujayra hajmidagi sezilarli farq bilan bog'liq.
Bakteriyalar qirolligining tirik vakillari son-sanoqsiz turlarga ega, ammo ularning barchasida umumiy narsa bor:
- aniq yadroning yo'qligi;
- juda kichik (o'simlik va hayvon hujayralariga nisbatan) hujayra o'lchamlari;
- Biologik birlik - bu hujayraning o'zi; ular birlashganda, biz bakteriyalar koloniyasi haqida gapiramiz.
Bu zamburug'lar, o'simliklar va hayvonlarning paydo bo'lishiga imkon yaratgan bakteriyalar shohligining vakillari edi. Sayyorada paydo bo'lgan mikroorganizmlar nafaqat mavjud sharoitlarga moslashdi - ular yashash joylarini faol ravishda o'zgartirib, sifat jihatidan yangi xususiyatlarni yaratdilar.

Tabiatdagi azot va uglerodning aylanishi faqat mikroorganizmlar tufayli sodir bo'ladi. Olimlar biosferadan mikroblar olib tashlansa, sayyoradagi hayot omon qolmasligini aniqladi.
Prokariotlarning biosfera siklidagi roli
Sayyoradagi hayotning paydo bo'lishida bakteriyalar qirolligi vakillari biosferaning shakllanishida faol ishtirok etdilar. Zamonaviy biosfera faoliyat darajasini saqlab turish uchun mikroorganizmlarga muhtoj - tabiatdagi energiya va moddalarning aylanishi mikroblar tomonidan ta'minlanadi.
Tirik mikroblarning biosfera jarayonlaridagi dominant roliga misol qilib unumdor tuproq qatlamini yaratish va saqlashni keltirish mumkin.
Gaz va oksidlanish funksiyalaridan tashqari mikroorganizmlarning geokimyoviy funksiyalari ham muhim ahamiyat kasb etmoqda. Fermentlarning faolligi va konsentratsiya funktsiyalari sayyoramizning geokimyosiga sezilarli ta'sir ko'rsatadi.
Mikroorganizmlarning tur xilma-xilligi
Bakteriyalar qirolligi vakillari biosferaning barcha darajalarida joylashdilar va mikroblar mavjudligi bilan ular sayyora biosferasining yuqori va quyi chegaralarini aniqlaydilar. Bunday turli xil jismoniy parametrlarga ega bo'lgan muhitda yashaydigan mikroblar bir qator xususiyatlar bilan farqlanadi.
- Tirik bakteriya hujayrasi shakliga ko'ra:
- sharsimon kokklar;
- novda shaklidagi;
- qiyshiq, vibrion va spiroketalarga bo'linadi.
- Tananing kosmosda harakat qilish usuliga ko'ra:
- flagellasiz (Brownianga o'xshash xaotik harakat);
- flagella yordamida (raqam butun perimetr bo'ylab birdan ko'pgacha o'zgaradi).
- Bakteriyalar qirolligi vakillarining metabolik xususiyatlariga ko'ra:
- noorganiklardan - avtotroflardan zarur moddalarni sintez qilish;
- organik moddalarni qayta ishlash - geterotroflar.
- Energiya olish usuliga ko'ra:
- nafas olish (aerob va anaerob mikroorganizmlar);
- fermentatsiya;
- fotosintez (kislorodsiz va kislorodsiz).

Mikroblar va viruslar o'rtasidagi munosabatlarning xususiyatlari - bir xil nomdagi shohliklarning vakillari
Prokariotlarning super shohligi ikkita shohlikni - bakteriyalar va viruslarni birlashtiradi, ular o'xshashlikdan ko'ra ko'proq farqlarga ega. Masalan, bakteriyalar hayotni ta'minlash uchun zarur bo'lgan barcha moddalarni sintez qilsa, viruslar umuman oqsil sinteziga qodir emas. Ular hatto o'z turlarini ham o'z-o'zidan ko'paytira olmaydilar, faqat birovning hujayrasiga kirib borish orqali.
Viruslar mezbon hujayraning DNKsini to'sib qo'yadi va uni o'zlari bilan almashtiradi - buning natijasida ushlangan hujayra bosqinchi virusning nusxalarini ishlab chiqaradi, bu odatda uning o'limiga olib keladi.