Forskare mikrobiologer och deras prestationer. Abstrakt historia om utvecklingen av mikrobiologi. Framväxten av vetenskapen om mikrobiologi
Grundaren av rysk mikrobiologi är L. Tsenkovsky (1822-1887). Objekten för hans forskning var mikroskopiska protozoer, alger och svampar. Han upptäckte och beskrev ett stort antal protozoer, studerade deras morfologi och utvecklingscykler. Detta gjorde att han kunde dra slutsatsen att det inte fanns någon skarp gräns mellan växternas och djurens värld. Han organiserade också en av de första Pasteur-stationerna i Ryssland och föreslog ett vaccin mot mjältbrand ("levande Tsenkovsky-vaccin").
Namnet på I. Mechnikov (1845-1916) är förknippat med utvecklingen av en ny riktning inom mikrobiologi - immunologi. För första gången inom vetenskapen utvecklade Mechnikov och experimentellt bekräftade den biologiska teorin om immunitet, som gick till historien som Mechnikovs fagocytiska teori. Denna teori är baserad på idén om cellulära skyddsanordningar i kroppen. Mechnikov, i experiment på djur (daphnia, sjöstjärnelarver), visade att leukocyter och andra celler av mesodermalt ursprung har förmågan att fånga och smälta främmande partiklar (inklusive mikrober) som kommer in i kroppen. Detta fenomen, som kallas fagocytos, utgjorde grunden för den fagocytiska teorin om immunitet och har fått universellt erkännande. Mechnikov utvecklade de frågor som ställdes ytterligare och formulerade en allmän teori om inflammation som en skyddande reaktion av kroppen och skapade en ny riktning inom immunologi - läran om antigenspecificitet. För närvarande blir det allt viktigare i samband med utvecklingen av problemet med organ- och vävnadstransplantation och studiet av cancerimmunologi.
Mechnikovs viktigaste verk inom området medicinsk mikrobiologi inkluderar studier av koleras patogenes och biologin av koleraliknande vibrios, syfilis, tuberkulos och skovvis feber. Mechnikov är grundaren av doktrinen om mikrobiell antagonism, som fungerade som grunden för utvecklingen av vetenskapen om antibiotikaterapi. Idén om mikrobiell antagonism användes av Mechnikov för att utveckla problemet med livslängd. Genom att studera fenomenet åldrande av kroppen kom Mechnikov till slutsatsen. Att dess viktigaste orsak är kronisk förgiftning av kroppen med förruttnande produkter som produceras i tjocktarmen av förruttnande bakterier.
Av praktiskt intresse är Mechnikovs tidiga arbeten om användningen av svampen Isaria destructor för att bekämpa åkerskadegöraren - spannmålsbaggen. De ger anledning att betrakta Mechnikov som grundaren av den biologiska metoden för att bekämpa skadedjur av jordbruksväxter, en metod som får ökande tillämpning och popularitet i dessa dagar. Således har I.I. Mechnikov, en framstående rysk biolog som kombinerade egenskaperna hos en experimenterare, lärare och främjare av vetenskaplig kunskap, var en man med stor anda och arbete, vars högsta utmärkelse var tilldelningen av Nobelpriset till honom 1909 för sin forskning om fagocytos. Utvecklingen av mikrobiologi i vårt land är också nära förbunden med namnet på den största vetenskapsmannen, vännen och kollegan I. Mechnikov, N.F. Gamaleya (1859-1949). Gamaleya ägnade hela sitt liv åt studiet av infektionssjukdomar och utvecklingen av åtgärder för att bekämpa deras patogener. Gamaleya gjorde ett stort bidrag till studiet av tuberkulos, kolera och rabies, 1886 organiserade han tillsammans med I. Mechnikov den första Pasteur-stationen i Odessa och införde vaccination mot rabies i praktiken. Han upptäckte fågelvibrio - orsaken till koleraliknande sjukdom hos fåglar - och döpte den till Mechnikov vibrio för att hedra Ilya Ilyich. Sedan erhölls ett vaccin mot human kolera.
Gamaleya ägnade också stor uppmärksamhet åt epidemiologin av infektionssjukdomar. Han var en stor specialist inom området immunologi. Efter att ha utvecklat en originalmetod för att producera smittkoppsvaccin föreslog han först idén att isolera de mest kompletta antigenerna från bakterier och använda dem för att förbereda så kallade kemiska vacciner. Gamaleya var den första som observerade och beskrev fenomenet spontan lysis av bakterier under påverkan av ett då okänt medel - en bakteriofag. Därför anses Gamaleya inte bara vara en av grundarna av medicinsk mikrobiologi, utan också av immunologi och virologi.
S.N.Vinogradsky, V.L. Omelyansky, B.L. Isachenko, D.I. Ivanovsky, G.A. Nadson, som var grundarna till vissa riktningar inom allmän mikrobiologi. Skapandet av till exempel läran om markmikroorganismernas ekologi är oupplösligt kopplat till namnet på den ryske forskaren S. Vinogradsky (1856-1953). Winogradsky gjorde också ett stort bidrag till kunskapen om den fysiologiska mångfalden i den mikrobiella världen. Han utförde klassiska arbeten om svavelbakteriers och järnbakteriers fysiologi, vilket resulterade i upptäckten av kemosyntes i bakterier - 1800-talets största upptäckt.
S. Winogradsky bevisade att det finns bakterier som självständigt syntetiserar organiskt material, med hjälp av energin för oxidation av mineralföreningar (vätesulfid, ammoniak) och koldioxid, etc. Det vill säga, en ny typ av näring av mikroorganismer upptäcktes - autotrofism.
Winogradskys ständiga krav var att studera mikroorganismer i deras naturliga livsmiljö eller förhållanden så nära naturliga som möjligt. Efter denna princip utvecklade han enkla och originella metoder för att studera markmikroorganismer. Metoden med elektiva (selektiva) medier har blivit allmänt erkänd och allmänt använd, vilket gör det möjligt att isolera ett antal nya mikroorganismer från den naturliga miljön och bestämma deras roll i ämnens kretslopp.
S. Winogradsky publicerade över 300 vetenskapliga artiklar om ekologi och fysiologi hos markmikroorganismer. Han anses med rätta vara jordmikrobiologins fader. En av de framstående grundarna av rysk mikrobiologi bör också inkludera S. Vinogradskys student V.L. Omalyansky (1867-1928). Han var inte bara en anmärkningsvärd vetenskapsman, utan också en begåvad lärare och populariserare av mikrobiologins prestationer. V. Omelyansky hade, liksom Pasteur, djup kunskap inom kemiområdet, vilket låg till grund för hans och det ekologiska studiet av mikroorganismer. Omfånget av vetenskapliga intressen för V. Omelyansky är mycket brett, men huvudriktningen för hans forskning är relaterad till studiet av kretsloppet av ämnen i naturen, där han tilldelade mikroorganismer en betydande roll. Han studerade processerna för nedbrytning av organiskt material och var den första som isolerade cellulosanedbrytande bakterier, beskrev deras fysiologi och kemin i själva processen.
Omelyansky studerade djupt och omfattande mikroorganismer som är involverade i kvävets kretslopp, särskilt frilevande kvävefixerare och nitrifierare. Ett av Omelyanskys senaste verk, "The Role of Microorganisms in the Weathering of Rocks", tillhör ett nytt forskningsområde. Detta arbete låg till grund för geologisk mikrobiologi.
Omelyanskys stora förtjänst är skapandet av den första ryska läroboken "Fundamentals of Microbiology", som publicerades 1909 och gick igenom 9 upplagor. I den sammanfattade Omelyansky resultaten av mikrobiologisk forskning och gav allmänna diagram över kretsloppet i naturen för enskilda element, inklusive kväve, kol, svavel och järn. I decennier har denna lärobok varit en uppslagsbok för specialister.
Den ekologiska riktningen inom mikrobiologi utvecklades framgångsrikt av B.L. Isachenko (1871-1948). Verk inom området akvatisk mikrobiologi har blivit allmänt kända. Han var den första som studerade mikroorganismernas utbredning i Ishavet och påpekade deras viktiga roll i geologiska processer och i cirkulationen av ämnen i vattendrag.
D.I. gav ett stort bidrag till utvecklingen av inhemsk och världsmikrobiologi. Ivanovsky (1864-1920), som upptäckte växtvirus 1892 och därigenom lade grunden för en ny vetenskap - virologi. Den engelske virologen N. Pirie betonade vikten av Ivanovskys forskning och skrev: "Den enorma betydelsen av Ivanovskys upptäckt för teoretisk naturvetenskap ligger i det faktum att han upptäckte en ny form av existens av proteinkroppar." Ivanovskys idéer spelade en avgörande roll i de efterföljande lysande framgångarna för virologi, som ett resultat av vilka orsakerna till de flesta virussjukdomar hos människor, djur, växter och mikroorganismer upptäcktes. Enligt slutsatsen från den amerikanske virologen Stanley bör namnet på Ivanovsky i virologi betraktas i samma ljus som namnen på Pasteur och Koch i mikrobiologi.
10. Sfäriska former(kocker) - sfäriska bakterier 0,5 - 1,0 mikron i storlek; Enligt det ömsesidiga arrangemanget av celler särskiljs mikrokocker, diplokocker, streptokocker, tetrakocker, sarcina och stafylokocker.
Mikrokocker (lat. små) - separat placerade celler eller i form av "paket".
Diplococci (lat. dubbel) - är arrangerade i par, eftersom cellerna inte separeras efter delning.
Streptokocker (från grekiskans streptos - kedja) är runda eller avlånga celler som bildar en kedja på grund av celldelning i samma plan och upprätthåller kopplingen mellan dem vid delningsplatsen.
Sarciner (från latin sarcina - bunt, bal) - är arrangerade i form av paket med 8 eller fler kocker, eftersom de bildas genom celldelning i tre ömsesidigt vinkelräta plan.
Stafylokocker (från det grekiska stafylokocket - druvklasar) är kocker som ligger i form av ett druvklase som ett resultat av delning i olika plan.
Stavformade bakterier skiljer sig i storlek, form på celländar och relativa arrangemang av celler. Celllängden varierar från 1,0 till 8,0, tjocklek från 0,5 till 2,0 µm. Stavar kan vara regelbundna (Escherichia coli) eller oregelbundna (corynebakterier) i form, inklusive förgrenade sådana, såsom actinomycetes. Lätt krökta stavar kallas vibrios (Vibrio cholerae). De flesta stavformade bakterier är ordnade slumpmässigt eftersom cellerna flyttas isär efter att de har delats.
Elementära kroppar kommer in i epitelcellen genom endocytos med bildandet av en intracellulär vakuol. Inuti cellen förstoras de och omvandlas till delande retikulära kroppar och bildar kluster i vakuoler (inneslutningar). Elementära kroppar bildas av retikulära kroppar, som lämnar cellerna genom exocytos eller cellys.
Mykoplasma är små bakterier (0,15 - 1,0 µm), omgivna av ett cytoplasmatiskt membran och utan cellvägg. På grund av frånvaron av en cellvägg är mykoplasma osmotiskt känsliga. De har en mängd olika former: coccoid, filamentös, kolvformad. Dessa former är synliga under faskontrastmikroskopi av rena kulturer av mykoplasma. Patogena mykoplasmer orsakar kroniska infektioner - mykoplasmos.
Aktinomyceter är förgrenade, filamentösa eller stavformade grampositiva bakterier. De fick sitt namn (från det grekiska actis - ray, mykes - svamp) på grund av bildandet i de drabbade vävnaderna av drusen - granuler av tätt sammanflätade trådar i form av strålar som sträcker sig från mitten och slutar i kolvformade förtjockningar. Aktinomyceter kan dela sig genom fragmentering av mycelet till celler som liknar stavformade och coccoid-bakterier. På lufthyfer hos aktinomyceter kan det bildas sporer som tjänar till reproduktion. Aktinomycetesporer är vanligtvis inte värmebeständiga.
En vanlig fylogenetisk gren med aktinomyceter bildas av de så kallade nocardiform (nocardioform) actinomycetes - en samlad grupp av stavformade, oregelbundet formade bakterier. Deras individuella representanter bildar förgreningsformer. Dessa inkluderar bakterier av släktena Corynebacterium, bdycobacterium, Hocardia, etc.
Nocardi-liknande aktinomyceter kännetecknas av närvaron i cellväggen av sockerarterna arabinos, galaktos, samt mykolsyror och stora mängder fettsyror. Mykolsyror och cellväggslipider bestämmer bakteriers syraresistens, i synnerhet mykobakterier tuberkulos och spetälska (när de färgas enligt Ziehl-Neelsen är de röda, och icke syraresistenta bakterier och vävnadselement, sputum är blå).
Slingrade former är spiralformade bakterier, till exempel spirilla, som ser ut som korkskruvsformade slingrande celler. Patogen spirilla inkluderar orsaksmedlet sodoku (råttbettssjukdom). De invecklade inkluderar även Campylobacter, Helicobacter, som har kurvor som vingen på en flygande mås; bakterier som spiroketer är också nära dem.
Spirochetes är tunna, långa, invecklade (spiralformade) bakterier som skiljer sig från spirillum i sin rörlighet på grund av böjningsförändringar i celler. Spirochetes har en yttre membrancellvägg som omger en protoplasmatisk cylinder med ett cytoplasmatiskt membran. Under det yttre membranet av cellväggen (i periplasman) finns periplasmatiska fibriller (flagella), som, som om de vrider sig runt spiroketens protoplasmatiska cylinder, ger den en spiroketform (primära krullar av spiroketen). Fibrillerna är fästa i cellens ändar och riktade mot varandra. Den andra änden av fibrillerna är fri. Antalet och arrangemanget av fibriller varierar mellan arterna. Fibriller är involverade i rörelsen av spiroketer, vilket ger cellerna rotations-, böjnings- och translationsrörelser. I det här fallet bildar spiroketer slingor, lockar och böjar, som kallas sekundära lockar.
Spirochetes uppfattar inte färgämnen bra. De färgas med Romanovsky-Giemsa-metoden eller silverfärgning, och i sin levande form undersöks de med enkelkontrast- eller mörkfältsmikroskopi.
Leptospira (släktet Leptospira) har grunda och frekventa lockar - i form av ett vridet rep. Ändarna på dessa spiroketer är krökta som krokar med förtjockningar i ändarna. Bildar sekundära lockar, de tar formen av bokstäverna S eller C; har 2 axiella gängor. Den patogena representanten L. förhör orsakar leptospiros.
Den genomsnittliga storleken på bakterier är 0,5-5 mikron. Escherichia coli, till exempel, har dimensioner på 0,3-1 gånger 1-6 mikron, Staphylococcus aureus har en diameter på 0,5-1 mikron, Bacillus subtilis 0,75 gånger 2-3 mikron. Den största kända bakterien är Thiomargarita namibiensis, som når en storlek på 750 mikron (0,75 mm). Den andra är Epulopiscium fishelsoni, som har en diameter på 80 mikron och en längd på upp till 700 mikron och lever i matsmältningskanalen hos operationsfisken Acanthurus nigrofuscus. Achromatium oxaliferum når storlekar på 33 gånger 100 mikron, Beggiatoa alba - 10 gånger 50 mikron. Spirochetes kan bli upp till 250 µm långa med en tjocklek på 0,7 µm. Samtidigt inkluderar bakterier de minsta organismerna med en cellstruktur. Mycoplasma mycoides är 0,1-0,25 mikron i storlek, vilket liknar storleken på stora virus som tobaksmosaik, kokoppor eller influensa. Enligt teoretiska beräkningar blir en sfärisk cell med en diameter på mindre än 0,15-0,20 mikron oförmögen till oberoende reproduktion, eftersom den fysiskt inte innehåller alla nödvändiga biopolymerer och strukturer i tillräckliga mängder.
Det har dock beskrivits nanobakterier som är mindre än den "acceptabla" storleken och skiljer sig mycket från vanliga bakterier. De, till skillnad från virus, är kapabla till oberoende tillväxt och reproduktion (extremt långsamt). De har hittills varit lite studerade, deras levande natur ifrågasätts.
Med en linjär ökning av en cells radie ökar dess yta i proportion till kvadraten av radien och dess volym i proportion till kuben, därför är förhållandet mellan yta och volym högre hos små organismer än i större, vilket för de förstnämnda innebär ett mer aktivt utbyte av ämnen med miljön. Metabolisk aktivitet, mätt med olika indikatorer, per enhet biomassa är högre i små former än i stora. Därför ger små storlekar även för mikroorganismer bakterier och arkéer fördelar i tillväxt- och reproduktionshastighet jämfört med mer komplexa eukaryoter och bestämmer deras viktiga ekologiska roll.
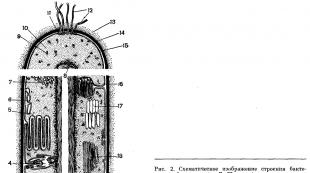
11. Det allmänna diagrammet över strukturen hos en bakteriecell visas i figuren 2. Den inre organisationen av en bakteriecell är komplex. Varje systematisk grupp av mikroorganismer har sina egna specifika strukturella egenskaper.
Cellvägg. Bakteriecellen är täckt med ett tätt membran. Detta ytskikt, som ligger utanför det cytoplasmatiska membranet, kallas cellväggen (Fig. 2, 14). Väggen utför skyddande och stödjande funktioner, och ger också cellen en permanent, karakteristisk form (till exempel formen av en stav eller coccus) och representerar cellens yttre skelett. Detta täta skal gör att bakterier liknar växtceller, vilket skiljer dem från djurceller, som har mjuka skal. Inuti bakteriecellen är det osmotiska trycket flera gånger, och ibland tiotals gånger högre än i den yttre miljön. Därför skulle cellen snabbt brista om den inte skyddades av en så tät, stel struktur som cellväggen.
Tjockleken på cellväggen är 0,01-0,04 mikron. Det utgör från 10 till 50 % av den torra massan av bakterier. Mängden material som utgör cellväggen förändras under bakterietillväxt och ökar vanligtvis med åldern.
Den huvudsakliga strukturella komponenten i väggarna, grunden för deras stela struktur i nästan alla bakterier som studerats hittills, är murein (glykopeptid, mukopeptid). Detta är en organisk förening med en komplex struktur, som inkluderar kvävebärande sockerarter - aminosocker och 4-5 aminosyror. Dessutom har cellväggsaminosyror en ovanlig form (D-stereoisomerer), som sällan finns i naturen.
Cellväggens beståndsdelar, dess komponenter, bildar en komplex, stark struktur.
Med hjälp av en färgningsmetod som först föreslogs 1884 av Christian Gram, kan bakterier delas in i två grupper: grampositiva och gramnegativa. Gram-positiva organismer kan binda vissa anilinfärgämnen, såsom kristallviolett, och efter behandling med jod och sedan alkohol (eller aceton) behåller jod-färgämneskomplexet. Samma bakterier där detta komplex förstörs under påverkan av etylalkohol (cellerna blir missfärgade) klassificeras som gramnegativa.
Den kemiska sammansättningen av cellväggarna hos grampositiva och gramnegativa bakterier är olika.
Hos grampositiva bakterier innefattar cellväggarnas sammansättning, förutom mukopeptider, polysackarider (komplexa, högmolekylära sockerarter), teichoic syror (komplexa föreningar i sammansättning och struktur, bestående av sockerarter, alkoholer, aminosyror och fosforsyra ). Polysackarider och teichoic syror är förknippade med väggramverket - murein. Vi vet ännu inte vilken struktur dessa komponenter i cellväggen hos grampositiva bakterier bildar. Med hjälp av elektroniska fotografier av tunna snitt (skiktning) upptäcktes inga grampositiva bakterier i väggarna. Förmodligen är alla dessa ämnen mycket tätt sammankopplade.
Väggarna hos gramnegativa bakterier är mer komplexa i kemisk sammansättning; de innehåller en betydande mängd lipider (fetter) associerade med proteiner och sockerarter till komplexa komplex - lipoproteiner och lipopolysackarider. Det finns i allmänhet mindre murein i cellväggarna hos gramnegativa bakterier än hos grampositiva bakterier. Väggstrukturen hos gramnegativa bakterier är också mer komplex. Med hjälp av ett elektronmikroskop fann man att väggarna hos dessa bakterier är flerskiktade (fig. 6).
Det inre lagret består av murein. Ovanför detta finns ett bredare lager av löst packade proteinmolekyler. Detta lager är i sin tur täckt med ett lager av lipopolysackarid. Det översta lagret består av lipoproteiner.
Cellväggen är permeabel: genom den passerar näringsämnen fritt in i cellen och metaboliska produkter kommer ut i miljön. Stora molekyler med hög molekylvikt passerar inte genom skalet.
Kapsel. Cellväggen hos många bakterier omges ovanpå av ett lager av slemhinnor - en kapsel (fig. 7). Kapselns tjocklek kan vara många gånger större än själva cellens diameter, och ibland är den så tunn att den bara kan ses genom ett elektronmikroskop – en mikrokapsel.
Kapseln är inte en väsentlig del av cellen, den bildas beroende på de förhållanden som bakterierna befinner sig i. Det fungerar som ett skyddande skydd för cellen och deltar i vattenmetabolismen, vilket skyddar cellen från att torka ut.
Den kemiska sammansättningen av kapslar är oftast polysackarider. Ibland består de av glykoproteiner (komplexa komplex av sockerarter och proteiner) och polypeptider (släktet Bacillus), i sällsynta fall - av fiber (släktet Acetobacter).
Slemämnen som utsöndras i substratet av vissa bakterier orsakar till exempel den slemtrådiga konsistensen av bortskämd mjölk och öl.
Cytoplasma. Hela innehållet i en cell, med undantag av kärnan och cellväggen, kallas cytoplasma. Den flytande, strukturlösa fasen av cytoplasman (matrisen) innehåller ribosomer, membransystem, mitokondrier, plastider och andra strukturer, samt reservnäringsämnen. Cytoplasman har en extremt komplex, fin struktur (skiktad, granulär). Med hjälp av ett elektronmikroskop har många intressanta detaljer om cellstrukturen avslöjats.
Det yttre lipoprotoida skiktet av bakterieprotoplasten, som har speciella fysikaliska och kemiska egenskaper, kallas det cytoplasmatiska membranet (fig. 2, 15).
Inuti cytoplasman finns alla vitala strukturer och organeller.
Det cytoplasmatiska membranet spelar en mycket viktig roll - det reglerar inträdet av ämnen i cellen och frisättningen av metaboliska produkter till utsidan.
Genom membranet kan näringsämnen komma in i cellen som ett resultat av en aktiv biokemisk process som involverar enzymer. Dessutom sker syntesen av vissa cellkomponenter i membranet, främst komponenter i cellväggen och kapseln. Slutligen innehåller det cytoplasmatiska membranet de viktigaste enzymerna (biologiska katalysatorer). Det ordnade arrangemanget av enzymer på membran gör det möjligt att reglera deras aktivitet och förhindra att vissa enzymer förstörs av andra. Förknippade med membranet är ribosomer - strukturella partiklar på vilka protein syntetiseras. Membranet består av lipoproteiner. Den är tillräckligt stark och kan säkerställa den tillfälliga existensen av en cell utan skal. Det cytoplasmatiska membranet utgör upp till 20 % av cellens torrmassa.
Mellan plasmamembranet och cellväggen finns en koppling i form av desmoser - broar. Det cytoplasmatiska membranet ger ofta upphov till invaginationer - invaginationer in i cellen. Dessa invaginationer bildar speciella membranstrukturer i cytoplasman som kallas mesosomer. Vissa typer av mesosomer är kroppar separerade från cytoplasman med sitt eget membran. Många vesikler och tubuli är packade inuti dessa membransäckar (fig. 2). Dessa strukturer utför en mängd olika funktioner i bakterier. Vissa av dessa strukturer är analoger till mitokondrier. Andra utför funktionerna hos det endoplasmatiska retikulumet eller Golgi-apparaten. Genom invaginering av det cytoplasmatiska membranet bildas också bakteriers fotosyntesapparat. Efter invagination av cytoplasman fortsätter membranet att växa och bildar staplar (tabell 30), som i analogi med växtkloroplastgranulat kallas tylakoidstaplar. I dessa membran, som ofta fyller det mesta av bakteriecellens cytoplasma, är pigment (bakterioklorofyll, karotenoider) och enzymer (cytokromer) som utför fotosyntesprocessen lokaliserade.
Bakteriecytoplasman innehåller ribosomer, proteinsyntetiserande partiklar med en diameter på 200A. Det finns mer än tusen av dem i en bur. Ribosomer består av RNA och protein. Hos bakterier är många ribosomer fritt placerade i cytoplasman, några av dem kan vara associerade med membran.
Ribosomer är centrum för proteinsyntesen i cellen. Samtidigt ansluter de ofta till varandra och bildar aggregat som kallas polyribosomer eller polysomer.
Bakteriecellers cytoplasma innehåller ofta granuler av olika former och storlekar. Däremot kan deras närvaro inte betraktas som något slags permanent tecken på en mikroorganism, det är vanligtvis till stor del relaterat till de fysiska och kemiska förhållandena i miljön. Många cytoplasmatiska inneslutningar är sammansatta av föreningar som fungerar som en källa till energi och kol. Dessa reservämnen bildas när kroppen tillförs tillräckligt med näringsämnen och, omvänt, används när kroppen befinner sig i förhållanden som är mindre gynnsamma när det gäller näring.
Hos många bakterier består granulat av stärkelse eller andra polysackarider - glykogen och granulosa. Vissa bakterier, när de odlas i ett sockerrikt medium, har droppar av fett inuti cellen. En annan utbredd typ av granulära inneslutningar är volutin (metakromatingranulat). Dessa granuler består av polymetafosfat (ett reservämne som innehåller fosforsyrarester). Polymetafosfat fungerar som en källa till fosfatgrupper och energi för kroppen. Bakterier är mer benägna att ackumulera volutin under ovanliga näringsförhållanden, såsom svavelfria medier. I cytoplasman hos vissa svavelbakterier finns droppar av svavel.
Förutom olika strukturella komponenter består cytoplasman av en flytande del - den lösliga fraktionen. Den innehåller proteiner, olika enzymer, t-RNA, vissa pigment och lågmolekylära föreningar - sockerarter, aminosyror.
Som ett resultat av förekomsten av föreningar med låg molekylvikt i cytoplasman uppstår en skillnad i det osmotiska trycket av cellinnehållet och den yttre miljön, och detta tryck kan vara olika för olika mikroorganismer. Det högsta osmotiska trycket observeras i grampositiva bakterier - 30 atm; i gramnegativa bakterier är det mycket lägre - 4-8 atm.
Kärnkraftsapparat. Den nukleära substansen, deoxiribonukleinsyra (DNA), är lokaliserad i den centrala delen av cellen.
Bakterier har inte en sådan kärna som högre organismer (eukaryoter), utan har sin analog - den "nukleära ekvivalenten" - nukleoiden (se fig. 2, 8), som är en evolutionärt mer primitiv form av organisering av kärnämne. Mikroorganismer som inte har en riktig kärna, men har en analog till den, klassificeras som prokaryoter. Alla bakterier är prokaryoter. I cellerna hos de flesta bakterier är huvuddelen av DNA koncentrerat på ett eller flera ställen. I eukaryota celler finns DNA i en specifik struktur - kärnan. Kärnan är omgiven av ett membran.
I bakterier packas DNA mindre tätt, till skillnad från riktiga kärnor; En nukleoid har inte ett membran, en nukleolus eller en uppsättning kromosomer. Bakteriellt DNA är inte associerat med huvudproteinerna - histoner - och finns i nukleoiden i form av ett knippe fibriller.
Flagella. Vissa bakterier har bihangsstrukturer på ytan; Den mest utbredda av dem är flageller - bakteriers rörelseorgan.
Flagellen är förankrad under det cytoplasmatiska membranet med hjälp av två par skivor. Bakterier kan ha en, två eller flera flageller. Deras placering är annorlunda: i ena änden av cellen, vid två, över hela ytan, etc. (Fig. 9). Bakteriella flageller har en diameter på 0,01-0,03 mikron, deras längd kan vara många gånger större än cellens längd. Bakteriella flageller består av ett protein - flagellin - och är tvinnade spiralformade filament.
 23 Enzymer eller enzymer (från lat.fermentum, Grekisk ζύμη, ἔνζυμον - surdeg) - vanligtvis proteinmolekyler eller RNA-molekyler (ribozymer) eller deras komplex som accelererar (katalyserar) kemiska reaktioner i levande system. Reaktanterna i en enzymkatalyserad reaktion kallas substrat, och de resulterande ämnena kallas produkter. Enzymer är substratspecifika (ATPas katalyserar nedbrytningen av endast ATP, och fosforylaskinas fosforylerar endast fosforylas). Enzymaktivitet kan regleras av aktivatorer och inhibitorer (aktivatorer ökar, hämmare minskar) Proteinenzymer syntetiseras på ribosomer och RNA syntetiseras i kärnan.
23 Enzymer eller enzymer (från lat.fermentum, Grekisk ζύμη, ἔνζυμον - surdeg) - vanligtvis proteinmolekyler eller RNA-molekyler (ribozymer) eller deras komplex som accelererar (katalyserar) kemiska reaktioner i levande system. Reaktanterna i en enzymkatalyserad reaktion kallas substrat, och de resulterande ämnena kallas produkter. Enzymer är substratspecifika (ATPas katalyserar nedbrytningen av endast ATP, och fosforylaskinas fosforylerar endast fosforylas). Enzymaktivitet kan regleras av aktivatorer och inhibitorer (aktivatorer ökar, hämmare minskar) Proteinenzymer syntetiseras på ribosomer och RNA syntetiseras i kärnan.
Enzymers funktioner
Enzymer finns i alla levande celler och hjälper till att omvandla vissa ämnen (substrat) till andra (produkter). Enzymer fungerar som katalysatorer i nästan alla biokemiska reaktioner som förekommer i levande organismer - de katalyserar mer än 4000 olika biokemiska reaktioner. Enzymer spelar en viktig roll i alla livsprocesser och styr och reglerar kroppens ämnesomsättning.
Liksom alla katalysatorer påskyndar enzymer både framåt- och bakåtreaktioner, vilket sänker processens aktiveringsenergi. I detta fall skiftar den kemiska jämvikten varken framåt eller bakåt. En utmärkande egenskap hos enzymer jämfört med icke-proteinkatalysatorer är deras höga specificitet - bindningskonstanten för vissa substrat till protein kan nå 10−10 mol/l eller mindre. Varje enzymmolekyl kan utföra från flera tusen till flera miljoner "operationer" per sekund. Till exempel, en molekyl av enzymet renin, som finns i magslemhinnan hos en kalv, kurar omkring 106 molekyler mjölkkaseinogen på 10 minuter vid en temperatur av 37 °C. Dessutom är effektiviteten hos enzymer mycket högre än effektiviteten hos icke-proteinkatalysatorer - enzymer påskyndar reaktioner med miljoner och miljarder gånger, icke-proteinkatalysatorer - hundratals och tusentals gånger. Se även Katalytiskt perfekt enzym
ENZYMERS GRUNDLÄGGANDE EGENSKAPER
Enzymer, som är proteiner, har ett antal egenskaper som är karakteristiska för denna klass av organiska föreningar som skiljer sig från egenskaperna hos oorganiska katalysatorer.
Termisk labilitet av enzymer. Hastigheten för kemiska reaktioner beror på temperaturen, så enzymkatalyserade reaktioner är också känsliga för temperaturförändringar. Det har fastställts att hastigheten för de flesta biokemiska reaktioner ökar med 2 gånger med en temperaturökning med 10°C och, omvänt, minskar med 2 gånger med en minskning av temperaturen med 10°C. Denna indikator kallas temperaturkoefficienten. På grund av enzymets proteinnatur kommer emellertid termisk denaturering med ökande temperatur att minska den effektiva koncentrationen av enzymet med en motsvarande minskning av reaktionshastigheten. Således, vid en temperatur som inte överstiger 45–50°C, ökar reaktionshastigheten enligt teorin om kemisk kinetik. Vid temperaturer över 50°C börjar den termiska denatureringen av enzymproteinet ha ett stort inflytande på reaktionshastigheten, vilket leder till att den enzymatiska processen helt upphör (Fig. 4.16).
Således är termolabilitet, eller känslighet för ökad temperatur, en av de karakteristiska egenskaperna hos enzymer som skarpt skiljer dem från oorganiska katalysatorer. I närvaro av den senare ökar reaktionshastigheten exponentiellt med ökande temperatur (se kurva "a" i fig. 4.16). Vid en temperatur på 100°C förlorar nästan alla enzymer sin aktivitet (det enda undantaget är uppenbarligen ett muskelvävnadsenzym - myokinas, som tål uppvärmning upp till 100°C). Den optimala temperaturen för verkan av de flesta enzymer hos varmblodiga djur är 40°C; under dessa förhållanden är reaktionshastigheten maximal på grund av en ökning av den kinetiska energin hos de reagerande molekylerna. Vid låga temperaturer (0°C och lägre) förstörs enzymer som regel inte, även om deras aktivitet sjunker nästan till noll. I alla fall är tidpunkten för exponering för lämplig temperatur viktig. För närvarande, för pepsin, trypsin och ett antal andra enzymer, har förekomsten av ett direkt samband bevisats

Ris. 4.17. Beroende av hastigheten för en enzymkatalyserad reaktion på pH (pilen indikerar pH-optimum).
mellan hastigheten för enzyminaktivering och graden av proteindenaturering. Det bör noteras att termolabiliteten hos enzymer definitivt påverkas av koncentrationen av substratet, pH i miljön och andra faktorer. Beroende av enzymaktivitet på pH i miljön. Enzymer är vanligtvis mest aktiva inom en smal zon med vätejonkoncentration, vilket för djurvävnader huvudsakligen motsvarar fysiologiska pH-värden på 6,0–8,0 utvecklade i evolutionsprocessen. När den ritas grafiskt har en klockformad kurva en specifik punkt vid vilken enzymet uppvisar maximal aktivitet; denna punkt kallas miljöns optimala pH för verkan av detta enzym (Fig. 4.17). Vid bestämning av enzymaktivitetens beroende av koncentrationen av vätejoner, utförs reaktionen vid olika pH-värden i mediet, vanligtvis vid en optimal temperatur och närvaron av tillräckligt höga (mättande) koncentrationer av substratet. I tabell Tabell 4.3 visar de optimala pH-värdena för ett antal enzymer. 
Från data i tabellen. 4.3 kan man se att pH-optimum för enzymverkan ligger inom fysiologiska värden. Undantaget är pepsin, vars pH-optimum är 2,0 (vid pH 6,0 är det inte aktivt och stabilt). Detta förklaras för det första av den strukturella organisationen av enzymmolekylen och för det andra av det faktum att pepsin är en komponent i magsaft som innehåller fri saltsyra, vilket skapar en optimal sur miljö för detta enzyms verkan. Å andra sidan ligger pH-optimumet för arginas i den starkt alkaliska zonen (ca 10,0); Det finns ingen sådan miljö i leverceller, därför fungerar arginas uppenbarligen inte i sin optimala pH-zon in vivo.
Enligt moderna koncept är effekten av förändringar i miljöns pH på enzymmolekylen att påverka tillståndet och graden av jonisering av sura och basiska grupper (särskilt COOH-gruppen av dikarboxylaminosyror, SH-gruppen av cystein imidazolkvävet av histidin, NH2-gruppens lysin, etc.). Med skarpa förändringar från miljöns optimala pH kan enzymer genomgå konformationsförändringar, vilket leder till förlust av aktivitet på grund av denaturering eller en förändring i laddningen av enzymmolekylen. Vid olika pH-värden av mediet kan det aktiva centret vara i en delvis joniserad eller icke-joniserad form, vilket påverkar den tertiära strukturen av proteinet och följaktligen bildandet av det aktiva enzym-substratkomplexet. Dessutom är joniseringstillståndet för substrat och kofaktorer viktigt.
Enzymspecificitet. Enzymer har hög verkningsspecificitet. Denna egenskap skiljer dem ofta avsevärt från oorganiska katalysatorer. Således kan finmald platina och palladium katalysera reduktionen (med deltagande av molekylärt väte) av tiotusentals kemiska föreningar med olika strukturer. Enzymets höga specificitet bestäms, som noterat, av den konformationella och elektrostatiska komplementariteten mellan molekylerna i enzymets substrat och den unika strukturella organisationen av det aktiva centret, vilket ger "igenkänning", hög affinitet och selektivitet för förekomsten av en reaktion av tusentals andra kemiska reaktioner som sker samtidigt i levande varelser.
Beroende på verkningsmekanismen särskiljs enzymer med relativ (eller grupp) och absolut specificitet. För verkan av vissa hydrolytiska enzymer är således typen av kemisk bindning i substratmolekylen av största betydelse. Till exempel bryter pepsin ner proteiner av animaliskt och vegetabiliskt ursprung i samma utsträckning, trots att dessa proteiner skiljer sig väsentligt från varandra både i kemisk struktur och aminosyrasammansättning, och i fysikalisk-kemiska egenskaper. Pepsin bryter dock inte ner varken kolhydrater eller fetter. Detta förklaras av det faktum att applikationspunkten, verkningsstället för pepsin, är peptid-CO-NH-bindningen. För verkan av lipas, som katalyserar hydrolysen av fetter till glycerol och fettsyror, är en liknande plats esterbindningen. Trypsin, kymotrypsin, peptidaser, enzymer som hydrolyserar α-glykosidbindningar (men inte β-glykosidbindningar som finns i cellulosa) i polysackarider etc. har liknande gruppspecificitet. Vanligtvis är dessa enzymer involverade i matsmältningsprocessen, och deras gruppspecificitet är mer sannolikt har allt en viss biologisk betydelse. Vissa intracellulära enzymer är också utrustade med relativ specificitet, till exempel hexokinas, som katalyserar fosforyleringen av nästan alla hexoser i närvaro av ATP, även om det samtidigt i celler finns enzymer specifika för varje hexos som utför samma fosforylering (se kapitel 10).
Absolut verkningsspecificitet är förmågan hos ett enzym att katalysera omvandlingen av endast ett enda substrat. Eventuella förändringar (modifieringar) i strukturen av substratet gör det otillgängligt för enzymets verkan. Exempel på sådana enzymer är arginas, som bryter ner arginin under naturliga förhållanden (i kroppen), ureas, som katalyserar nedbrytningen av urea, etc.
Det finns experimentella bevis på förekomsten av så kallad stereokemisk specificitet, på grund av förekomsten av optiskt isomera L- och D-former eller geometriska (cis- och trans-) isomerer av kemiska ämnen. Således är oxidaser av L- och D-aminosyror kända, även om endast L-aminosyror finns i naturliga proteiner. Varje typ av oxidaser verkar endast på sin specifika stereoisomer.
Ett tydligt exempel på stereokemisk specificitet är bakteriellt aspartatdekarboxylas, som katalyserar avlägsnandet av CO 2 endast från L-asparaginsyra och omvandlar det till L-alanin. Stereospecificitet uppvisas av enzymer som katalyserar och syntetiska reaktioner. Från ammoniak och α-ketoglutarat syntetiseras alltså L-isomerglutaminsyra, som är en del av naturliga proteiner, i alla levande organismer. Om en förening existerar i form av cis- och transisomerer med olika arrangemang av grupper av atomer runt dubbelbindningen, så kan i regel endast en av dessa geometriska isomerer tjäna som substrat för enzymets verkan. Till exempel katalyserar fumaras omvandlingen av endast fumarsyra (transisomer), men verkar inte på maleinsyra (cis-isomer):
På grund av deras höga verkningsspecificitet säkerställer enzymer att endast vissa kemiska reaktioner från en stor mängd möjliga omvandlingar sker med hög hastighet i cellernas mikroutrymme och hela organismen, och reglerar därigenom intensiteten av ämnesomsättningen.
Enzymers roll i ämnesomsättningen.
Mikrobiologi spelar en viktig roll i mänsklighetens historia. Ursprunget till denna vetenskap går tillbaka till 600- till 500-talen f.Kr. e. I dessa avlägsna tider började folk redan inse att sjukdomar inte bara uppstår så. Och detta händer på grund av mikroskopiska, osynliga för ögat, mikroorganismer. Hur uppstod och tog vetenskapen form?
Vad är mikrobiologi?
Mikrobiologi är en vetenskap som sysslar med forskning och studier av olika mikroorganismers livsprocesser som inte kan ses utan specialutrustning. De kan ha olika typer av ursprung: växt, djur. En av de grundläggande vetenskaperna är mikrobiologi. För dess fördjupade studie används många andra vetenskaper, nämligen:
- kemi;
- fysik;
- cytologi;
- biologi osv.
Det finns bara två typer av mikrobiologi: allmän, individuell. Allmän mikrobiologi handlar om studier av små mikroorganismers struktur och livsprocesser på olika nivåer. Och individuell mikrobiologi (eller privat) handlar om studiet av enskilda typer av mikrober.
Under 1800-talet bidrog framsteg inom medicinområdet, i synnerhet mikrobiologi, till bildandet av immunologi, som idag anses vara en allmän biologisk disciplin. I utvecklingen av mikrobiologi kan tre huvudstadier särskiljas:
- Att avslöja det faktum att det verkligen finns små mikroorganismer i naturen som inte går att upptäcka utan specialutrustning.
- Differentiering av arter.
- Studie av immunitet och sjukdomar (smittsamma).
Mikrobiologins huvuduppgift är den detaljerade studien av mikroorganismernas egenskaper. För detta ändamål används specialutrustning, till exempel mikroskop. Med deras hjälp kan du se små organismer och bestämma deras form och plats. Inom medicinen utövas ett experiment när små mikroorganismer medvetet implanteras i ett friskt djur. Detta hjälper till att rekonstruera och studera varje infektionsstadium.
Fransk upptäcktsresande Louis Pasteur
Den 27 december 1822 föddes den blivande store vetenskapsmannen Louis Pasteur i östra Frankrike. I tidig ålder var han intresserad av konstområdet. Men senare blev han intresserad av naturvetenskap. Han studerade i Paris vid Ecole Superieure. Efter avslutade studier var han förutbestämd att bli lärare i naturvetenskap.
På 48 av 1800-talet presenterade Louis resultaten av sin egen vetenskapliga forskning. Det var han som gav bevis för att vinsyra innehåller 2 typer av kristaller som polariserar ljus på helt olika sätt. Denna betydelsefulla händelse markerade början på hans lysande framgång inom vetenskapen.
Louis Pasteur är skaparen av mikrobiologi. Innan hans arbete började, antog forskarna bara att jäst bildade den kemiska processen. Och Louis Pasteur, efter att ha genomfört en serie studier, kunde bevisa detta faktum. Han upptäckte att det finns två typer av sådana mikroorganismer: vissa bildar alkohol, medan andra förstör den. Senare kunde han ta reda på att med långsam uppvärmning förstörs onödiga bakterier, vilket avsevärt ökade kvaliteten på alkoholhaltiga produkter.
Forskaren var också intresserad av bildandet av mögel på mat. Han bevisade senare att mögel orsakas av sporer i miljön. Ju färre av dem det finns i rymden, desto långsammare blir maten förstörd.
Hans forskning hjälpte till att rädda sidenindustrin i Frankrike. Och även många människoliv, eftersom det var han som uppfann rabiesvaccinet.
Den tyske vetenskapsmannen Robert Koch
Koch Robert anses vara en samtida med Pastser. Hans födelse inträffade i december 1843. Vid 23 års ålder tog han examen från medicinskt universitet och fick ett diplom, varefter han arbetade på flera medicinska institutioner.
Hans betydande karriär började med arbetet som bakteriolog. Han studerade mjältbrand på sjuka djur. Hans forskning visade att infekterade individer har många främmande mikroorganismer som friska djur inte har. Dessa bakterier var stavformade.
Koch blev senare intresserad av tuberkulos. De första studierna utfördes på liket av en arbetare som dog av konsumtion. En detaljerad studie av organen ledde inte till identifiering av patogena bakterier. Koch föreslog då att proverna skulle färgas. Och faktiskt, forskaren märkte några pinnar mellan lungvävnaderna. Efteråt utvecklade Robert Koch ett vaccin mot tuberkulos, men det kunde inte bota sjukdomen, utan det avgjorde till 100% om patienten var smittad eller inte. Detta vaccin används fortfarande idag.
Framväxten av vetenskapen om mikrobiologi
Människan mötte effekterna av den vitala aktiviteten hos mikroorganismer mycket tidigare än deras officiella upptäckt. Människor jäste medvetet mjölk, jäst deg och vin. Även i den antika grekiska vetenskapsmannens verk hittades rader om vad han antyder om förhållandet mellan sjukdomar och farliga patogena ångor.
Anthony van Leeuwenhoek bekräftade dessa gissningar med hjälp av ett förstoringsglas han uppfann. Med dess hjälp kunde Anthony undersöka de omgivande föremålen. Det visade sig att små organismer lever på dessa föremål som är osynliga för blotta ögat. Men han kunde aldrig bevisa deras deltagande i att smitta människor med farliga sjukdomar.
Förebyggande behandling av hemmet för att förebygga sjukdomar tillhandahölls av hinduerna. År 1771, i Moskva, använde en militärläkare först desinficering av tillhörigheter från personer som smittats av pesten och vaccinerade även de som hade kontakt med de smittade.
Den mest fascinerande historien handlar om upptäckten av smittkoppsvaccination. Det användes också av perser, turkar och kineser. Det hände så här: försvagade bakterier introducerades till en person, eftersom man trodde att sjukdomen skulle bli lättare på detta sätt. Den engelske läkaren Edward Jenner noterade att de flesta som inte har smittkoppor inte blev smittade genom nära kontakt med smittade personer. Detta faktum märktes hos mjölkpigor som var i kontakt med kor smittade med smittkoppor. Studiet av detta faktum varade cirka 10 år. Som ett resultat injicerade vetenskapsmannen det sjuka konblodet i en frisk pojke. Senare inokulerade Jenner den unge mannen med bakterier från en sjuk person. Så upptäcktes ett vaccin, tack vare vilket människor blev befriade från denna fruktansvärda sjukdom.
Forskning av inhemska forskare
De mest kända upptäckterna inom mikrobiologin, gjorda av vetenskapliga forskare från hela världen, gör det klart att nästan alla sjukdomar kan övervinnas. Inhemska forskare har gjort en enorm investering i bildandet av modern vetenskap. Peter I gjorde 1698 bekantskap med Leeuwenhoek, som i sin tur visade honom operationen av ett mikroskop.
L.S. Tsenkovsky publicerade sin vetenskapliga forskning, där mikroorganismer klassificerades som organismer av växtursprung. Han använde även Pasteurs metoder i kampen mot mjältbrand.
I.I. Mechnikov bildade teorin om immunitet. Han gjorde starka argument för att många celler i kroppen har alla möjligheter att undertrycka virusbakterier på egen hand. Hans studier blev grunden för studiet av inflammation. Mechnikov studerade människokroppen och försökte förstå varför den åldras. Professorn ville hitta en metod som skulle öka livslängden. Han trodde att giftiga element som uppstår under aktiviteten av förruttnande mikroorganismer förgiftar människokroppen. Enligt Mechnikov ska kroppen befolkas med fermenterade mjölkmikroorganismer som dämpar skadliga mikroorganismer. Professorn trodde att den förväntade livslängden på detta sätt kunde ökas avsevärt.
Mechnikov studerade ett stort antal allvarliga sjukdomar: tuberkulos, tyfus, kolera och många andra.
Teknisk mikrobiologi
Teknisk mikrobiologi studerar bakterier som används vid framställning av vitaminer och vissa ämnen. Huvudproblemet inom detta område anses vara tillväxten av vetenskapliga och tekniska tekniker inom tillverkning (särskilt inom livsmedelssektorn).
Att behärska industriell mikrobiologi riktar en specialist till behovet av noggrann överensstämmelse med absolut alla allmänt accepterade sanitära standarder i produktionen. Genom att studera denna vetenskap kan du förhindra att många produkter förstörs. Ämnet studeras mer av framtida experter inom livsmedelsindustrin.
Innovativa tekniker
Mikrobiologi är grunden för innovativ teknik. Mikroorganismer och deras värld har ännu inte studerats fullt ut. De flesta forskare är övertygade om att det med hjälp av mikroorganismer är möjligt att utveckla teknologier som inte kommer att ha några analoger. Det är bioteknik som kommer att bli grunden för de senaste tekniska upptäckterna.
När man utforskar olje- och kolfyndigheter används bakterier. Det är ingen hemlighet att bränslereserverna redan håller på att ta slut. Därför rekommenderar forskare redan användningen av mikrobiologiska metoder för att utvinna alkoholer från förnybara källor.
Mikrobiologiska tekniker kommer att hjälpa till att övervinna miljö- och energiproblem. Otroligt nog gör dock mikrobiologisk bearbetning av organiska rester det möjligt att rena miljön, samt få biogas som inte är sämre än naturgas. Denna typ av bränsleutvinningsmetod kräver inga stora kostnader. Idag finns det i naturen en stor mängd använt material för bearbetning runt omkring.
Många moderna forskare tror att det i framtiden är biologi som kommer att göra det möjligt att övervinna många energi- och miljösvårigheter som har alla möjligheter att dyka upp på kortast möjliga tid.
Mikrobiologi har gått igenom en lång utvecklingsväg, beräknad till många årtusenden. Redan under V-IV årtusendet f.Kr. människan njöt av frukterna av mikroorganismernas aktivitet utan att veta om deras existens. Vintillverkning, brödbakning, osttillverkning, lädergarvning är inget annat än processer som sker med deltagande av mikroorganismer. Sedan, i gamla tider, antog forskare och tänkare att många sjukdomar orsakades av några främmande osynliga orsaker av en levande natur.
Följaktligen uppstod mikrobiologi långt före vår tideräkning. I sin utveckling gick den igenom flera stadier, inte så mycket relaterade kronologiskt som bestämdes av de viktigaste prestationerna och upptäckterna.
Historien om utvecklingen av mikrobiologi kan delas in i fem stadier: heuristisk, morfologisk, fysiologisk, immunologisk och molekylärgenetisk.
Den heuristiska perioden (IV-IIV årtusende f.Kr. - 1500-talet e.Kr.) förknippas snarare med logiska och metodologiska metoder för att hitta sanningen, d.v.s. heuristik än med några experiment och bevis. Den tidens tänkare (Hippokrates, den romerske författaren Varro, etc.) gjorde antaganden om arten av infektionssjukdomar, miasmer och små osynliga djur. Dessa idéer formulerades till en sammanhängande hypotes många århundraden senare i skrifterna av den italienska läkaren D. Fracastoro (1478 - 1553), som uttryckte idén om att leva contagium vivum, som orsakar sjukdomar. Dessutom orsakas varje sjukdom av sin egen smitta. För att skydda sig mot sjukdomar rekommenderades de att isolera patienten, sätta i karantän, bära masker och behandla föremål med vinäger.
Således var D. Fracastoro en av grundarna av epidemiologi, det vill säga vetenskapen om orsaker, tillstånd och mekanismer för bildandet av sjukdomar och metoder för att förebygga dem.
Bevis på förekomsten av osynliga patogener blev dock möjligt efter uppfinningen av mikroskopet. Prioritet i upptäckten av mikroorganismer tillhör den holländska amatörnaturforskaren Antonio Leeuwenhoek (1b32 - 1723). Linnehandlaren A. Levenguk var förtjust i glasslipning och förde denna konst till perfektion, genom att konstruera ett mikroskop som gjorde det möjligt att förstora de aktuella föremålen med 300 gånger.
Genom att studera olika föremål under ett mikroskop (regnvatten, infusioner, tandplack, blod, avföring, spermier), observerade A. Leeuwenhoek de minsta djuren, som han kallade animalculi. A. Leeuwenhoek rapporterade regelbundet sina observationer till Royal Society of London, och 1695 sammanfattade han dem i boken "The Secrets of Nature Discovered by Anthony Leeuwenhoek."
Således, med uppfinningen av mikroskopet av A. Leeuwenhoek, börjar nästa steg i utvecklingen av mikrobiologi, kallad morfologisk.
A. Leeuwenhoeks upptäckt väckte enorm uppmärksamhet från specialister, och han hade många studenter och anhängare. Frågor om uppkomsten av mikroorganismer, deras levnadsförhållanden, syfte och deltagande i förekomsten av mänskliga sjukdomar förblev dock oklara. Dessa frågor fick sedan tydliga svar i studier av många forskare.
Även om uppkomsten av sjukdomar var förknippad med de nu upptäckta mikroorganismerna, behövdes direkta bevis. Och de erhölls av den ryska epidemiologen D. Samoilovich (1744 - 1805). För att bevisa att pesten orsakades av en speciell patogen, infekterade han sig själv med sekret från buboen från en person som led av pesten och blev sjuk av pesten. Lyckligtvis överlevde D. Samoilovich. Därefter utfördes heroiska experiment på självinfektion för att bevisa smittsamheten hos en viss mikroorganism av de ryska läkarna G. N. Minkh och O. O. Mochutkovsky, I. I. Mechnikov och andra.
Frågan om metoden för utseende och reproduktion av mikroorganismer löstes i en tvist med den då dominerande teorin om spontan generering. Trots att den italienske vetenskapsmannen L. Spallanzani i mitten av 1700-talet. observerade uppdelningen av bakterier under ett mikroskop, åsikten att de självgenererar (uppstår från röta, smuts, etc.) motbevisades inte.
Detta gjordes av den enastående franske vetenskapsmannen Louis Pasteur (1822 - 1895), som i ett kvickt experiment som var briljant i sin enkelhet visade att spontan generation inte existerar. L. Pasteur placerade en steril buljong i en kolv, som kommunicerade med atmosfärisk luft genom ett krökt S-format rör. I en sådan väsentligen öppen kolv förblev buljongen genomskinlig när den stod under lång tid, eftersom rörets krökning inte tillät mikroorganismer att penetrera med damm från luften in i kolven.
Mikrobiologins snabba utveckling på 1800-talet. ledde till upptäckten av orsakerna till många infektionssjukdomar (mjältbrand, pest, stelkramp, difteri, dysenteri, kolera, tuberkulos, etc.).
Slutligen, 1892, upptäckte den ryske botanikern D.I. Ivanovsky (1864. 1920) virus - representanter för vira-riket. Dessa levande varelser passerade genom filter som höll kvar bakterier och kallades därför filtrerbara virus. Först upptäcktes viruset som orsakar tobakssjukdomen känd som "tobaksmosaik", sedan mul- och klövsjukeviruset, gula febern och många andra virus. Det blev dock möjligt att se viruspartiklar först efter uppfinningen av elektronmikroskopet, eftersom virus inte är synliga i ljusmikroskop. Hittills omfattar virusriket (vira) upp till 1000 patogena arter av virus. Först nyligen har ett antal nya virus upptäckts, inklusive viruset som orsakar AIDS.
Det råder ingen tvekan om att perioden av upptäckter av nya virus och bakterier kommer att fortsätta. Upptäckten av nya mikroorganismer åtföljdes av studiet av inte bara deras struktur, utan också deras livsaktivitet. Därför brukar 1800-talet, särskilt dess andra hälft, kallas den fysiologiska perioden i mikrobiologins utveckling. Detta stadium är förknippat med namnet på L. Pasteur, som blev grundaren av medicinsk mikrobiologi, såväl som immunologi och bioteknik.
En mångsidig utbildad, briljant experimenterare, medlem av den franska medicinakademin, L. Pasteur gjorde ett antal enastående upptäckter. Under en kort period från 1857 till 1885 bevisade han att jäsning (mjölksyra, alkohol, ättiksyra) inte är en kemisk process, utan orsakas av mikroorganismer. Motbevisade teorin om spontan generering; upptäckte fenomenet anaerobios, d.v.s. möjligheten att mikroorganismer lever i frånvaro av syre. Lade grunden för desinfektion, asepsis och antiseptika; upptäckt ett sätt att skydda sig mot infektionssjukdomar genom vaccination.
Många av L. Pasteurs upptäckter medförde enorma praktiska fördelar för mänskligheten. Genom uppvärmning (pastörisering), sjukdomar i öl och vin besegrades mjölksyraprodukter orsakade av mikroorganismer; antiseptika introducerades för att förhindra purulenta komplikationer av sår; Baserat på L. Pasteurs principer har många vacciner utvecklats för att bekämpa infektionssjukdomar.
Men betydelsen av L. Pasteurs verk går långt utöver bara dessa praktiska landvinningar. L. Pasteur förde mikrobiologi och immunologi till i grunden nya positioner, visade mikroorganismernas roll i människors liv, ekonomi, industri, infektionspatologi, och lade fast de principer som mikrobiologi och immunologi utvecklas efter i vår tid.
L. Pasteur var dessutom en framstående lärare och organisatör av naturvetenskap. Pasteurinstitutet i Paris, som grundades 1888 med offentliga medel, är fortfarande en av de ledande vetenskapliga institutionerna i världen. Det är ingen slump att det humana immunbristviruset (HIV) upptäcktes av vetenskapsmannen vid detta institut, L. Montagnier (samtidigt som amerikanen R. Gallo).
Den fysiologiska perioden i utvecklingen av mikrobiologi är också förknippad med namnet på den tyske forskaren Robert Koch, som utvecklade metoder för att erhålla rena bakteriekulturer, färgning av bakterier under mikroskopi och mikrofotografering. Koch-triaden formulerad av R. Koch är också känd, som fortfarande används för att identifiera orsaken till sjukdomen.
L. Pasteurs arbete med vaccination öppnade ett nytt steg i utvecklingen av mikrobiologin, med rätta kallad "immunologisk".
Principen om försvagning (försvagning) av mikroorganismer genom passager genom ett känsligt djur eller genom att hålla mikroorganismer under ogynnsamma förhållanden (temperatur, torkning) gjorde det möjligt för L. Pasteur att få vaccin mot rabies, mjältbrand och kycklingkolera; denna princip används fortfarande vid framställning av vacciner. Följaktligen är L. Pasteur grundaren av den vetenskapliga immunologin, även om före honom var metoden att förebygga smittkoppor genom att infektera människor med kokoppor, utvecklad av den engelske läkaren E. Jenner, känd. Denna metod har dock inte utvidgats till att förebygga andra sjukdomar.
Efter L. Pasteurs arbete dök många studier upp som försökte förklara orsakerna och mekanismerna för bildandet av immunitet efter vaccination. Verken av I. I. Mechnikov och P. Ehrlich spelade en enastående roll i detta.
P. Ehrlich, en tysk kemist, lade fram en humoral (från latinets humor - flytande) teori om immunitet. Han trodde att immunitet uppstår som ett resultat av bildandet av antikroppar i blodet som neutraliserar giftet. Detta bekräftades av upptäckten av antitoxiner - antikroppar som neutraliserar toxiner hos djur som injicerades med difteri eller stelkrampstoxin (E. Bering, S. Kitazato). Studierna av I. I. Mechnikov (1845 - 1916) visade dock att speciella celler, makro- och mikrofager, spelar en stor roll i bildandet av immunitet. Dessa celler absorberar och smälter främmande partiklar, inklusive bakterier. I. I. Mechnikovs forskning om fagocytos visade övertygande att det, förutom humoral, finns cellulär immunitet. I. I. Mechnikov, närmaste assistent och anhängare till L. Pasteur, anses välförtjänt vara en av grundarna av immunologi. Hans arbete markerade början på studiet av immunkompetenta celler som den morfologiska grunden för immunsystemet, dess enhet och biologiska väsen. Den immunologiska perioden kännetecknas av upptäckten av immunsystemets huvudsakliga reaktioner på genetiskt främmande ämnen (antigener): antikroppsbildning och fagocytos/överkänslighet av fördröjd typ (DTH), överkänslighet av omedelbar typ (IHT), tolerans, immunologiskt minne. HRT och HNT är två reaktioner som ligger bakom allergier (från grekiskans allos - annan och ergon - verkan), det vill säga sjukdomar som kännetecknas av vissa kliniska symtom på grund av en atypisk, pervers reaktion på ett antigen. Allergiska reaktioner kan förekomma till exempel mot serumpreparat, antibiotika, animaliska och vegetabiliska proteiner, husdamm, ludd, ull m.m.
År 1915 observerade den ryske läkaren M. Raisky första gången fenomenet immunologiskt minne, d.v.s. snabb energiproduktion av antikroppar mot upprepad administrering av samma antigen.
Därefter associerade F. Vernet detta med bildandet av minnesceller i kroppen - T-lymfocyter - efter det initiala mötet med antigenet. 1953 upptäckte den engelske vetenskapsmannen P. Medawar och den tjeckiske vetenskapsmannen M. Hasek fenomenet tolerans, tolerans, resistens mot antigen, d.v.s. ett tillstånd där immunsystemet inte svarar på ett antigen. Tolerans mot självantigener utvecklas under embryonalperioden och kan skapas på konstgjord väg genom att införa antigen under embryonperioden eller omedelbart efter ett barns eller ett djurs födelse. Fenomenet immunologisk tolerans används inom kirurgi för att lösa problemet med organ- och vävnadstransplantation.
Det bör också noteras vikten av upptäckten under denna period av antigener från normala organ och vävnader hos människor och djur och individuella, antigena skillnader hos människor och djur. En vanlig indikation på dessa antigena skillnader är de individuella blodtyperna hos människor. Den inhemska forskaren L.A. Zilber (1957) upptäckte antigener från maligna tumörer, vilket var början på studiet av antitumörimmunitet.
Under den immunologiska perioden för utvecklingen av mikrobiologi skapades ett antal teorier om immunitet: den humorala teorin om P. Ehrlich, den fagocytiska teorin om I. I. Mechnikov, teorin om idiotypiska interaktioner av N. Erne, hypofysen-hypothalamus-binjuren. teorin om immunitetsreglering av P. F. Zdrodovsky, etc. Den klonala selektionsteorin skapad av den australiensiska immunologen F. Burnet (1899 - 1986) förblir acceptabel för att förklara många fenomen och immunitetsmekanismer. Den amerikanske vetenskapsmannen S. Tanegawa utvecklade de genetiska aspekterna av denna teori.
Mikrobiologi och immunologi fick en särskilt snabb utveckling under 50-60-talet av vårt sekel. Följande skäl bidrog till detta:
· de viktigaste upptäckterna inom området molekylärbiologi, genetik, bioorganisk kemi;
· framväxten av nya vetenskaper såsom genteknik, bioteknik, datavetenskap;
· skapande av nya metoder och vetenskaplig utrustning som gör att vi kan tränga djupare in i den levande naturens hemligheter.
Sedan 50-talet började alltså en molekylärgenetisk period i utvecklingen av mikrobiologi och immunologi, som kännetecknas av ett antal fundamentalt viktiga vetenskapliga landvinningar och upptäckter. Dessa inkluderar:
· dechiffrera den molekylära strukturen och den molekylärbiologiska organisationen för många virus och bakterier; upptäckten av de enklaste livsformerna, det "smittsamma proteinet" prion;
· dechiffrera den kemiska strukturen och kemiska syntesen av vissa antigener. Till exempel den kemiska syntesen av lysozym, AIDS-viruspeptider (R.V. Petrov, V.T. Ivanov, etc.);
· upptäckt av nya antigener, till exempel tumörantigener (L.A. Zilber och andra), histokompatibilitetsantigener (HLA-system);
· dechiffrera strukturen av antikroppar-immunoglobuliner;
· utveckling av en metod för att odla djur- och växtceller och odla dem i industriell skala för att erhålla virala antigener;
· produktion av rekombinanta bakterier och rekombinanta virus. Syntes av individuella gener av virus och bakterier. Erhålla rekombinanta stammar av bakterier och virus som kombinerar egenskaperna hos föräldraindivider eller förvärvar nya egenskaper;
· skapande av hybridom genom fusion av immuna B-lymfocyter, antikroppsproducenter och cancerceller för att producera monoklonala antikroppar
· upptäckt av immunmodulatorer, immuncytokiner (interleukiner, interferoner, myelopeptider, etc.), endogena naturliga regulatorer av immunsystemet och deras användning för att förebygga och behandla olika sjukdomar;
· Erhållande av vacciner (hepatit B-vaccin, malaria, HIV-antigener och andra antigener), biologiskt aktiva peptider (interferoner, interleukiner, tillväxtfaktorer, etc.) med hjälp av bioteknologiska metoder och genteknik;
· utveckling av syntetiska vacciner baserade på naturliga eller syntetiska antigener och deras fragment, samt en artificiell bärare. adjuvans (assistent). immunstimulerande medel;
· studie av medfödda och förvärvade immunbrister, deras roll i immunopatologi och utvecklingen av immunkorrigerande terapi. Upptäckt av virus som orsakar immunbrist;
· utveckling av fundamentalt nya metoder för att diagnostisera infektionssjukdomar och icke-infektionssjukdomar (enzymimmunanalyser, radioimmunanalyser, immunblotting, nukleinsyrahybridisering). Skapande av testsystem baserade på dessa metoder för indikation, identifiering av mikroorganismer, diagnos av infektionssjukdomar och icke-infektionssjukdomar (tumörer, kardiovaskulära, autoimmuna, endokrina, etc.), samt upptäckt av störningar under vissa tillstånd (graviditet, blodtransfusion) , organtransplantation och etc.)
Teorier om immunitet. Utveckling av teorier om immunitet. Fagocytisk teori om immunitet. I. I. Mechnikov
Identifieringen av patogena mikroorganismers roll i utvecklingen av infektionssjukdomar och möjligheten att på konstgjord väg skapa immunitet har föranlett studier av faktorer som skyddar kroppen från infektionsämnen.
Pasteur föreslog teorin om uttömd makt; Enligt denna teori representerar "immunitet" ett tillstånd där människokroppen (som ett näringsmedium) inte stödjer utvecklingen av mikrober.
Författaren insåg dock snabbt att hans teori inte kunde förklara ett antal observationer. I synnerhet visade Pasteur att om du infekterar en kyckling med mjältbrand och håller benen i kallt vatten, kommer den att utveckla sjukdomen (under normala förhållanden är kycklingar immuna mot mjältbrand). Utvecklingen av fenomenet orsakade en minskning av kroppstemperaturen med 1-2 °C, det vill säga att det inte kunde vara tal om någon utarmning av näringsmediet i kroppen.
Fagocytisk teori om immunitet. I. I. Mechnikov
1883 dök en teori om immunitet upp, baserad på Charles Darwins evolutionära läror och baserad på studiet av matsmältning hos djur i olika stadier av biologisk utveckling. Författaren till den nya teorin, I. I. Mechnikov, upptäckte likheten mellan intracellulär nedbrytning av ämnen i amöbor, endodermceller av coelenterater och vissa celler av mesenkymalt ursprung (blodmonocyter, vävnadsmakrofager). Mechnikov introducerade termen "fagocyter" från grekiskan. fager, äta, + kytos, cell, och föreslog senare att dela upp dem i mikrofager och makrofager. Denna uppdelning underlättades också av prestationerna av P. Ehrlich, som differentierade flera typer av leukocyter genom färgning. I klassiska verk om den jämförande patologin av inflammation bevisade I. I. Mechnikov rollen av fagocytiska celler i elimineringen av patogener. 1901 publicerades hans monumentala slutverk "Immunity in Infectious Diseases" i Paris.
Ett betydande bidrag till spridningen av den fagocytiska teorin gjordes av arbetet av E. Roux och elever av I. I. Mechnikov (A. M. Bezredka, I. G. Savchenko, L. A. Tarasevich, F. Ya. Chistovich, V. I. Isaev).
I. I. Mechnikov (1845–1916)
Enastående rysk vetenskapsman. 1882, vid kongressen för ryska naturforskare och läkare, höll Mechnikov sitt berömda tal "Om kroppens försvar", där han gav en djup vetenskaplig motivering för kroppens immunitet mot infektionssjukdomar. Denna teori kallas den fagocytiska teorin om immunitet. Bildandet av denna teori baserades på hans observationer av intracellulär matsmältning hos olika marina djur (maneter, svampar, blötdjur, etc.). Mechnikov upptäckte i kroppen av dessa djur speciella "vandrande" celler som möter, fångar och slukar alla främmande kroppar som kommer in i kroppen, inklusive olika mikrober. Han kallade de "vandrande" cellerna fagocyter, det vill säga äterceller.
Mechnikov betonade att fagocyter skyddar kroppen och, som ett resultat av deras aktivitet, utvecklas immunitet mot infektionssjukdomar. Den odödliga läran om kroppens immunitet och dess biologiska teori om immunitet öppnade ett nytt stadium i utvecklingen av medicin. För denna upptäckt 1908 tilldelades I. I. Mechnikov Nobelpriset.
Charles Niccol kallade bildligt Mechnikov "mikrobiologins poet", som krediteras med att utveckla läran om mikrobiell antagonism, som blev den teoretiska grunden för produktionen av antibiotika, läkemedel som används för att behandla ett antal infektionssjukdomar. Mechnikov äger också originalforskning om experimentell syfilis och kolera.
Mechnikov studerade orsakerna till ålderdom. Han trodde att "ålderdom är en sjukdom som måste behandlas", påpekade han att den utvecklas under påverkan av kronisk förgiftning av kroppen orsakad av avfallsprodukter från representanter för tarmmikrofloran. Han inkluderade indol, fenol och skatol bland sådana produkter. För att tränga undan förruttnelsebakterier föreslog Mechnikov en diet som i första hand består av grönsaker, frukt och surmjölk, som innehåller mjölksyrabakterier, som är antagonister till förruttnelsebakterier.
Namnet I. I. Mechnikov åtnjuter nationellt erkännande. Den sovjetiska regeringen inrättade en guldmedalj och Mechnikov-priset, som delas ut för det bästa arbetet inom biologiområdet
Det är omöjligt att inte notera verken av M. M. Terekhovsky (1740-1796). Från 1770 arbetade Terekhovsky utomlands i cirka fem år, vid universitetet i Strasbourg, som var känt för sin medicinska skola. Här disputerade Terekhovsky för sin doktorsavhandling – hans huvudsakliga vetenskapliga arbete. Avhandlingen hade titeln "Mörkrets rike av Linnés Ciliates". Det satte uppgiften att utforska naturen och sätten för uppkomsten av mikroskopiska varelser i olika infusioner. Baserat på ett stort antal experiment kom forskaren till slutsatsen att "djur" är levande varelser. De dör av höga temperaturer, gifter och elektrisk ström. Den viktigaste slutsatsen var omöjligheten av spontan generering av "djur", vilket stred mot åsikterna från många forskare på den tiden. Terekhovsky skrev att Harveys (1578-1651) välkända position - "allt levande kommer från ett ägg" - får kraften hos ett axiom.
När han återvände till Ryssland arbetade Terekhovsky på medicinska skolor. År 1782 bekräftades han med rang av professor vid St. Petersburg General Land Hospital och chef för Botaniska trädgården (nuvarande Botaniska institutet vid USSR Academy of Sciences).
År 1835, baserat på de fakta som samlats vid denna tid, publicerade K. Ehrenberg en bok med en mycket betydelsefull titel: "Ciliater som perfekta organismer." Han delade in lägre varelser i 22 klasser. Boken åtföljdes av en atlas över ciliater, av vilka många beskrevs i detalj och namngavs enligt binär nomenklatur. De tre klasserna inkluderade bakterier.
RYSKA FEDERATIONENS UTBILDNINGSMINISTERIE
TULA STATE UNIVERSITY
Institutionen för sanitära, hygieniska och förebyggande discipliner
CHESTNOVA T.V., SMOLYANINOVA O.L.
MEDICINSK MIKROBIOLOGI, VIRUSOLOGI
OCH IMMUNOLOGI
(Utbildnings- och praktisk manual för studenter vid medicinska universitet).
TULA – 2008
UDC 576,8
Recensenter: …………
Medicinsk mikrobiologi, virologi och immunologi: Pedagogisk och praktisk manual / Ed. M422 T.V. Chestnovoy, O.L. Smolyaninova, –….., 2008. -….s.
Den pedagogiska och praktiska manualen skrevs av anställda vid avdelningen för sanitetshygieniska och förebyggande discipliner vid Tula State University i enlighet med de officiellt godkända programmen för undervisning i mikrobiologi (bakteriologi, virologi, mykologi, protozoologi) och immunologi för studenter vid medicinska universitet i alla fakulteter.
Den pedagogiska och praktiska handboken beskriver det bakteriologiska laboratoriet, beskriver mikroskopiska forskningsmetoder, grunderna för beredning av näringsmedier och innehåller information om morfologi, systematik och fysiologi hos bakterier, svampar, protozoer och virus. Egenskaper för olika patogena mikroorganismer, virus och metoder för deras laboratorieforskning ges också.
ALLMÄN MIKROBIOLOGI
Introduktion…………………………………………………………………………………………………………
Kort historia om utvecklingen av mikrobiologi…………………………………………………………………
Ämne 1. Morfologi och klassificering av mikroorganismer…………………………………………..
1.1. Mikrobiologiska laboratorier, deras utrustning, grundläggande säkerhetsåtgärder och arbetsregler i dem…………………………………………………………………………………………………..
1.2. Struktur och klassificering av mikroorganismer…………………………………………………………………………………
1.3. Struktur och klassificering av bakterier (prokaryoter)………………………………………………………………….
1.4. Struktur och klassificering av svamp…………………………………………………………………………..
1.5. Struktur och klassificering av protozoer……………………………………………………………….
1.6. Struktur och klassificering av virus………………………………………………………………………
Testa på ämnet………………………………………………………………………………………………..
Ämne 2. Mikroskopi………………………………………………………………………………………..
2.1. Mikroskop, deras struktur, typer av mikroskopi, mikroskopitekniker för mikroorganismer, regler för hantering av mikroskop………………………………………………………………….
2.2. Metoder för framställning och färgning av mikroskopiska preparat…………………………..
Testa på ämnet……………………………………………………………………………………………………….
Ämne 3. Mikroorganismers fysiologi……………………………………………………………………….
3.1. Tillväxt och reproduktion av bakterier. Reproduktionsfaser……………………………………………………………………….
3.2 Näringsmedier, principer för deras klassificering, krav på näringsmedier, metoder för att odla mikroorganismer…………………………………………………..
3.3. Näring av bakterier………………………………………………………………………………………….
3.4. Metabolism av bakteriecellen……………………………………………………………………….
3.5. Typer av plastbyte…………………………………………………………………………………………………………
3.6. Principer och metoder för att isolera rena kulturer. Bakteriella enzymer, deras identifiering. Intraspecifik identifiering (epidemiologisk märkning)…………………………………..
3.7. Funktioner i fysiologin för svampar, protozoer, virus och deras odling………………
3.8. Bakteriofager, deras struktur, klassificering och tillämpning………………………………………………………………..
Testa på ämnet………………………………………………………………………………………………………
Ämne 4. Miljöförhållandenas inverkan på mikroorganismer…………………………………………..
4.1. Effekten av fysikaliska, kemiska och biologiska faktorer på mikroorganismer………….
4.2. Konceptet med sterilisering, desinfektion, asepsis och antiseptika. Steriliseringsmetoder, utrustning. Kvalitetskontroll av desinfektion…………………………………………………………………..
Ämne 5. Normal mikroflora i människokroppen……………………………………………………………………….
5.1. Normoflora, dess betydelse för mikroorganismer. Begreppet övergående flora, dysbiotiska tillstånd, deras bedömning, metoder för korrigering………………………………………………………………..
Ämne 6. Genetik hos mikrober. …………………………………………………………………………………………………………..
6.1. Struktur av bakteriegenomet. Fenotypisk och genotypisk variation. Mutationer. Ändringar …………………………………………………………………………………………………………..
Genetiska rekombinationer av mikroorganismer. Grunderna i genteknik, praktisk tillämpning………………………………………………………………………………………………………………………………….
Testa på ämnet………………………………………………………………………………………………..
Ämne 7. Antimikrobiella medel……………………………………………………………………………………………….
7.1. Antibiotika naturliga och syntetiska. Klassificering av antibiotika efter kemisk struktur, mekanism, spektrum och typ av verkan. Metoder för att erhålla ………………………………….
7.2. Läkemedelsresistens hos bakterier, sätt att övervinna det. Metoder för att bestämma känslighet för antibiotika………………………………………………………………………………………………………..
Ämne 8. Läran om infektion…………………………………………………………………………………………………..
8.1. Begreppet infektion. Infektionsformer och perioder av infektionssjukdomar. Patogenicitet och virulens. Patogenicitetsfaktorer. Bakteriegifter, deras natur, egenskaper, produktion………………………………………………………………………………………………………….
8.2. Begreppet epidemiologisk övervakning av den smittsamma processen. Begreppet reservoar, infektionskälla, smittvägar och överföringsfaktorer………………………………………………………………………
Testa på ämnet………………………………………………………………………………………………..
ALLMÄN IMMUNOLOGI……………………………………………………………………………………………………….
Ämne 9. Immunologi…………………………………………………………………………………………
9.1. Begreppet immunitet. Typer av immunitet. Ospecifika skyddsfaktorer………….
9.2. Centrala och perifera organ i immunsystemet. Immunsystemets celler. Former av immunsvar………………………………………………………………………………………
9.3. Komplement, dess struktur, funktioner, aktiveringsvägar. Roll i immunitet………………..
9.4. Antigener, deras egenskaper och typer. Antigener från mikroorganismer………………………………………..
9.5. Antikroppar och antikroppsbildning. Struktur av immunglobuliner. Klasser av immunglobuliner och deras egenskaper …………………………………………………………………………………………………………………………
96. Serologiska reaktioner och deras tillämpning……………………………………………………………….
9.7. Immunbrist tillstånd. Allergiska reaktioner. Immunologiskt minne. Immunologisk tolerans. Autoimmuna processer………………………………………………………………………
9.8. Immunprofylax, immunterapi………………………………………………………………..
PRIVAT MIKROBIOLOGI……………………………………………………………………………………….
Ämne 10. Patogener av tarminfektioner……………………………………………………………………….
10.1. Salmonella……………………………………………………………………………………..
10.2. Shigella……………………………………………………………………………………………….
10.3. Escherichia……………………………………………………………………………………………………………………….
10.4. Vibrio cholerae……………………………………………………………………………………….
10.5. Yersinia……………………………………………………………………………………………….
Ämne 11. Livsmedelsburna giftiga infektioner. Livsmedelstoxicitet…………………………………………………………………………
11.1. Allmänna egenskaper och patogener för PTI……………………………………………………………….
11.2. Botulism…………………………………………………………………………………………..
Ämne 12. Patogener av purulenta inflammatoriska sjukdomar…………………………………………………………………
12.1. Patogena kocker (streptokocker, stafylokocker)………………………………………………………………………..
12.2. Gramnegativa bakterier (Haemophilus influenzae, Pseudomonas aeruginosa, Klebsiella, Proteus)...
12.3. Sår anaeroba clostridial och icke-clostridial infektioner………………………
Ämne 13. Patogener från bakteriella luftburna infektioner………………………………….
13.1. Corynebacteria …………………………………………………………………………………………………
13.2. Bordetella………………………………………………………………………………………………………………………………
13.3. Meningokocker………………………………………………………………………………………………..
13.4. Mykobakterier………………………………………………………………………………………………..
13.5. Legionella………………………………………………………………………………………………..
Ämne 14. Patogener av sexuellt överförbara sjukdomar (STD)………………………
14.1. Klamydia………………………………………………………………………………………………………..
14.2. Orsaken till syfilis………………………………………………………………………………………………………….
14.3. Gonokocker……………………………………………………………………………………………….
Ämne 15. Patogener av rickettsialsjukdomar………………………………………………………………………………………………………..
Ämne 16. Orsakande agens till bakteriella zoonotiska infektioner……………………………….
16.1. Francisella………………………………………………………………………………………………………
16.2. Brucella……………………………………………………………………………………………….
16.3. Det orsakande ämnet för mjältbrand………………………………………………………………………………………………..
16.4. Orsaken till pesten …………………………………………………………………………………………
16.5. Leptospira………………………………………………………………………………………………..
Ämne 17. Patogena protozoer………………………………………………………………………………………………..
17.1. Plasmodium malaria……………………………………………………………………………………………….
17.2. Toxoplasma……………………………………………………………………………………………….
17.3. Leishmania………………………………………………………………………………………………..
17.4. Orsaken till amöbiasis………………………………………………………………………………………….
17.5. Giardia …………………………………………………………………………………………………………………………………
Ämne 18. Sjukdomar orsakade av patogena svampar………………………………………………………………..
PRIVAT VIRUSOLOGI………………………………………………………………………………………………………..
Ämne 19. Patogener för akuta luftvägsvirusinfektioner………………………………………………………………………………………………………………
19.1. Influensavirus……………………………………………………………………………………….
19.2. Parainfluensa. PC-virus………………………………………………………………………………………………………
19.3. Adenovirus………………………………………………………………………………………………………………………
19.4. Rhinovirus………………………………………………………………………………………………..
19.5. Reovirus……………………………………………………………………………………………….
Ämne 20. Patogener från virala luftburna infektioner…………………………………………..
20.1. Mässling och påssjuka virus…………………………………………………………………………………………………………..
20.2. Herpesvirus …………………………………………………………………………………………………...
20.3. Rubellavirus………………………………………………………………………………………………………
Ämne 21. Poxyvirus………………………………………………………………………………………….
21.1. Orsaken till smittkoppor………………………………………………………………………………………………………….
Ämne 22. Enterovirala infektioner………………………………………………………………………..
22.1. Poliovirus …………………………………………………………………………………………………
22.2. ECHO-virus. Coxsackie-virus………………………………………………………………………………………………
Ämne 23. Retrovirus……………………………………………………………………………………….......
23.1. Orsaken till HIV-infektion………………………………………………………………………………………………..
Ämne 24. Arbovirala infektioner……………………………………………………………………………………………….
24.1. Rabdovirus……………………………………………………………………………………………………….
24.2. Flavivirus …………………………………………………………………………………………………………………………
24.3. Hantavirus……………………………………………………………………………………………….
Ämne 25. Orsakande agens för viral hepatit…………………………………………………………………………………………………
25.1. Hepatit A-virus……………………………………………………………………………………………….
25.2. Hepatit B-virus………………………………………………………………………………………………..
25.3. Hepatit C-virus………………………………………………………………………………………………..
DEL ETT. ALLMÄN MIKROBIOLOGI
Introduktion.
Mikrobiologi är en vetenskap som studerar mikroskopiska varelser som kallas mikroorganismer, deras biologiska egenskaper, systematik, ekologi och relationer med andra organismer.
Mikroorganismer inkluderar bakterier, actinomycetes, svampar, inklusive filamentösa svampar, jäst, protozoer och icke-cellulära former - virus, fager.
Mikroorganismer spelar en extremt viktig roll i naturen - de utför cirkulationen av organiska och oorganiska (N, P, S, etc.) ämnen, mineraliserar växt- och djurrester. Men de kan orsaka stor skada - orsaka skador på råvaror, livsmedel och organiska material. Detta kan leda till bildning av giftiga ämnen.
Många typer av mikroorganismer orsakar sjukdomar hos människor, djur och växter.
Samtidigt används mikroorganismer för närvarande i stor utsträckning i samhällsekonomin: med hjälp av olika typer av bakterier och svampar erhålls organiska syror (ättiksyra, citronsyra, etc.), alkoholer, enzymer, antibiotika, vitaminer och foderjäst . Bakning, vinframställning, bryggning, produktion av mejeriprodukter, jäsning av frukt och grönsaker, liksom andra grenar av livsmedelsindustrin arbetar på basis av mikrobiologiska processer.
För närvarande är mikrobiologi indelad i följande sektioner:
Medicinsk mikrobiologi - studerar patogena mikroorganismer som orsakar mänskliga sjukdomar och utvecklar metoder för att diagnostisera, förebygga och behandla dessa sjukdomar. Studerar sätten och mekanismerna för deras spridning och metoder för att bekämpa dem. I anslutning till kursen medicinsk mikrobiologi finns en separat kurs - virologi.
Veterinär mikrobiologi studerar patogena mikroorganismer som orsakar sjukdomar hos djur.
Bioteknik undersöker egenskaperna och villkoren för utveckling av mikroorganismer som används för att erhålla föreningar och läkemedel som används i den nationella ekonomin och medicinen. Det utvecklar och förbättrar vetenskapliga metoder för biosyntes av enzymer, vitaminer, aminosyror, antibiotika och andra biologiskt aktiva substanser. Biotekniken står också inför uppgiften att utveckla åtgärder för att skydda råvaror, livsmedel och organiska material från att förstöras av mikroorganismer, och att studera de processer som sker under lagring och bearbetning av dem.
Markmikrobiologin studerar mikroorganismernas roll i bildningen och fertiliteten av mark och i växtnäring.
Akvatisk mikrobiologi studerar vattenkropparnas mikroflora, dess roll i näringskedjor, i ämnens kretslopp, i förorening och behandling av dricks- och avloppsvatten.
Genetik av mikroorganismer, som en av de yngsta disciplinerna, undersöker den molekylära grunden för ärftlighet och variabilitet hos mikroorganismer, mönster för mutagenesprocesser, utvecklar metoder och principer för att kontrollera den vitala aktiviteten hos mikroorganismer och erhåller nya stammar för användning inom industri, jordbruk och medicin .
En kort historia om utvecklingen av mikrobiologi.
Äran för upptäckten av mikroorganismer tillhör den holländska naturforskaren A. Leeuwenhoek (1632-1723), som skapade det första mikroskopet med en förstoring på 300 gånger. År 1695 han publicerade boken "Secrets of Nature" med teckningar av kocker, stavar och spirilla. Detta väckte stort intresse bland naturvetare. Vetenskapens tillstånd vid den tiden tillät endast att beskriva nya arter (morfologisk period).
Början av den fysiologiska perioden är förknippad med den store franske vetenskapsmannen Louis Pasteurs (1822-1895) aktiviteter. Namnet Pasteur är förknippat med de största upptäckterna inom mikrobiologi: han undersökte jäsningens natur, etablerade möjligheten till liv utan syre (anaerobios), förkastade teorin om spontan generering och undersökte orsakerna till förstörelse av vin och öl. Han föreslog effektiva sätt att bekämpa patogener av matförstöring (pastörisering), utvecklade principen om vaccination och metoder för att få vaccin.
R. Koch, en samtida med Pasteur, introducerade sådd på fasta näringsmedier, räknade mikroorganismer, isolerade rena kulturer och steriliserade material.
Den immunologiska perioden i utvecklingen av mikrobiologi är associerad med namnet på den ryska biologen I.I. Mechnikov, som upptäckte läran om kroppens immunitet mot infektionssjukdomar (immunitet), var grundaren av den fagocytiska teorin om immunitet och upptäckte antagonism i mikrober. Samtidigt med I.I. Mechnikov studerade mekanismerna för immunitet mot infektionssjukdomar av den stora tyska forskaren P. Ehrlich, som skapade teorin om humoral immunitet.
Gamaleya N.F. – grundare av immunologi och virologi, upptäckte bakteriofagi.
DI. Ivanovsky upptäckte först virus och blev virologins grundare. Medan han arbetade i Nikitskys botaniska trädgård med studien av tobaksmosaiksjukdom, som orsakade enorma skador på tobaksplantager, 1892. fann att denna sjukdom, utbredd på Krim, orsakas av ett virus.
N.G. Gabrichevsky organiserade det första bakteriologiska institutet i Moskva. Han äger verk för studier av scharlakansfeber, difteri, pest och andra infektioner. Han organiserade produktionen av antidifteriserum i Moskva och använde det framgångsrikt för att behandla barn.
P.F. Zdrodovsky är en immunolog och mikrobiolog, känd för sitt grundläggande arbete om immunitetens fysiologi, såväl som inom området rickettsiologi och brucellos.
V.M. Zhdanov är en stor virolog, en av organisatörerna av den globala utrotningen av smittkoppor på planeten, som stod för ursprunget till molekylär virologi och genteknik.
M.P. Chumakov är en immunobioteknolog och virolog, arrangör av Institutet för Poliomyelit och Viral Encefalit, författare till ett oralt poliovaccin.
Z.V. Ermolyeva - grundaren av inhemsk antibiotikaterapi
UTBILDNINGSMINISTERIET OCH VETENSKAP I REPUBLIKEN KAZAKHSTAN
EAST KAZAKHSTANS STATSUNIVERSITET UPPFYLLD EFTER S.AMANZHOLOVA
Biologiska institutionen
ABSTRAKT
Disciplin: "Biologi och utveckling av mikroorganismer och virus"
På ämnet: "Historia om utvecklingen av mikrobiologi"
Slutförd av: elever i grupp UBG-09 (A)
Grushkovskaya D., Fefelova N.
Kontrolleras av: Kalenova K.Sh.
Ust-Kamenogorsk, 2011
Planen:
Inledning……………………………………………………………………………………… 3
1. UPPTÄCKT AV MIKROORGANISMER………………………………………………………………… …4
2. BESKRIVANDE (MORFOLOGISK) PERIOD I UTVECKLINGEN AV MIKROBIOLOGI (SLUTET AV 1600-talet – MITTEN AV 1800-talet) …………………..5
2.1. Utveckling av idéer om karaktären hos jäsnings- och sönderfallsprocesserna...5
2.2. Utveckling av idéer om den mikrobiella naturen hos infektionssjukdomar……………………………………………………………………………… ………………………….7
3. FYSIOLOGISK PERIOD (PASTERIAN) (ANDRA HALVAN AV 1800-TALET)……………………………………………………………………………………….8
3.1. Louis Pasteurs vetenskapliga verksamhet…………………………………………………………8
3.2. Mikrobiologins utveckling under andra hälften av 1800-talet…………………………10
4. UTVECKLING AV MIKROBIOLOGI UNDER 1900-TALET………………………………………15
Slutsats.................... ............................. ........................................................................... ...... 18
Litteratur ........................................................... .......... ................................ .............. .......... 19
INTRODUKTION
Mikrobiologi är en vetenskap som studerar strukturen, systematik, fysiologi, biokemi, genetik och ekologi hos organismer som är små i storlek och osynliga för blotta ögat. Dessa organismer kallas mikroorganismer eller mikrober.
Under en lång tid levde en person omgiven av osynliga varelser, använde produkterna av sin livsviktiga aktivitet (till exempel när man bakade bröd av sur deg, förberedde vin och vinäger), led när dessa varelser orsakade sjukdomar eller förstörda matförråd, men gjorde inte misstänker deras närvaro. Jag misstänkte inte eftersom jag inte såg, och jag såg inte eftersom storleken på dessa mikrovarelser låg mycket under den synlighetsgräns som det mänskliga ögat kan. Det är känt att en person med normal syn på optimalt avstånd (25-30 cm) kan urskilja ett föremål som mäter 0,07-0,08 mm i form av en punkt. En person kan inte lägga märke till mindre föremål. Detta bestäms av de strukturella egenskaperna hos hans synorgan.
Försök att övervinna den skapade naturliga barriären och utöka det mänskliga ögats kapacitet gjordes för länge sedan. Sålunda, under arkeologiska utgrävningar i det antika Babylon, hittades bikonvexa linser - de enklaste optiska instrumenten. Linserna var gjorda av polerad bergskristall. Vi kan tänka oss att människan med deras uppfinning tog det första steget på vägen till mikrovärlden.
Ytterligare förbättringar av optisk teknik går tillbaka till 1500- och 1600-talen. och är förknippad med utvecklingen av astronomi. Vid den här tiden designade holländska glaskvarnar de första teleskopen. Det visade sig att om linserna är placerade annorlunda än i ett teleskop kan man förstora upp väldigt små föremål. Ett mikroskop av denna typ skapades 1610 av G. Galileo. Uppfinningen av mikroskopet öppnade nya möjligheter för att studera levande natur.
Ett av de första mikroskopen, bestående av två bikonvexa linser som gav en ökning på cirka 30 gånger, designades och användes för att studera växternas struktur av den engelske fysikern och uppfinnaren R. Hooke. Han undersökte sektioner av kork och upptäckte den vanliga cellstrukturen hos trävävnad. Dessa celler kallades därefter "celler" av honom och avbildades i boken "Micrography". Det var R. Hooke som introducerade termen "cell" för att beteckna de strukturella enheter från vilka en komplex levande organism är uppbyggd. Ytterligare penetration i mikrovärldens hemligheter är oupplösligt kopplad till förbättringen av optiska instrument.
1.UPPTÄCKT AV MIKROORGANISMER
Mikroorganismer upptäcktes i slutet av 1600-talet, men deras verksamhet och till och med praktiska tillämpningar var kända mycket tidigare. Till exempel framställdes och användes produkter av alkohol-, mjölksyra- och ättiksyrajäsningar i de äldsta tiderna. Användbarheten av dessa produkter förklarades av närvaron av en "levande ande" i dem. Men idén om förekomsten av osynliga varelser började dyka upp när man identifierade orsakerna till infektionssjukdomar. Således föreslog Hippokrates (6:e århundradet f.Kr.), och senare Varro (2:a århundradet) att infektionssjukdomar orsakas av osynliga varelser. Men först på 1500-talet kom den italienska forskaren Giralamo Fracastoro till slutsatsen att överföringen av sjukdomar från person till person utförs med hjälp av de minsta levande varelserna, som han gav namnet contagium vivum. Det fanns dock inga bevis för sådana antaganden.
Om vi antar att mikrobiologi uppstod i det ögonblick då människan såg de första mikroorganismerna, så kan vi absolut exakt indikera mikrobiologins "födelsedag" och upptäckarens namn. Den här mannen är holländaren Antonie van Leeuwenhoek (1632-1723), en tillverkare från Delft. Intresserad av linfiberns struktur polerade han flera grova linser åt sig själv. Senare blev Leeuwenhoek intresserad av detta känsliga och mödosamma arbete och uppnådde stor perfektion i tillverkningen av linser, som han kallade "mikroskopi". I yttre form var dessa enkla bikonvexa glas inramade i silver eller mässing, men i sina optiska egenskaper hade Leeuwenhoek-linser, som gav en förstoring på 200 - 270 gånger, ingen motsvarighet. För att uppskatta dem räcker det att komma ihåg att den teoretiska gränsen för förstoring av en bikonvex lins är 250 - 300 gånger.
Utan naturlig utbildning, men med naturlig nyfikenhet, tittade Leeuwenhoek med intresse på allt som kom till hands: dammvatten, tandplack, pepparinfusion, saliv, blod och mycket mer. Från 1673 började Leeuwenhoek skicka resultaten av sina observationer till Royal Society of London, som han därefter valdes till medlem av. Totalt skrev Leeuwenhoek över 170 brev till Royal Society of London och testamenterade senare till det 26 av sin berömda "mikroskopi". Här är ett utdrag ur ett brev: ”Den 24 april 1676 tittade jag på vattnet i mikroskop och såg med stor förvåning ett stort antal små levande varelser i det. Vissa av dem var 3-4 gånger längre än de var breda, även om de inte var tjockare än hårstråna som täckte lusens kropp. Andra hade en vanlig oval form. Det fanns också en tredje typ av organismer – de mest talrika – små varelser med svansar.” Genom att jämföra beskrivningen som ges i detta avsnitt och de optiska egenskaperna hos linserna till Leeuwenhoeks förfogande kan vi dra slutsatsen att Leeuwenhoek var den första som såg bakterier 1676.
Leeuwenhoek upptäckte mikroorganismer överallt och kom fram till att världen omkring honom var tätt befolkad av mikroskopiska invånare. Leeuwenhoek ansåg att alla mikroorganismer han såg, inklusive bakterier, var små djur, som han kallade "animalcules", och var övertygad om att de var strukturerade på samma sätt som stora organismer, dvs de hade matsmältningsorgan, ben, svansar, etc. ..d.
Leeuwenhoeks upptäckter var så oväntade och till och med fantastiska att de under nästan 50 efterföljande år orsakade universell häpnad. Medan han var i Holland 1698, besökte Peter I Leeuwenhoek och pratade med honom. Från denna resa tog Peter I med sig ett mikroskop till Ryssland, och senare, 1716, tillverkades de första inhemska mikroskopen i verkstäderna vid hans hov.
2. BESKRIVANDE (MORFOLOGISK) PERIOD I UTVECKLINGEN AV MIKROBIOLOGI (SLUTET AV 1600-talet – MITTEN AV 1800-talet)
2.1. Utveckling av idéer om jäsnings- och sönderfallsprocessernas karaktär
Många processer som utförs av mikroorganismer har varit kända för människan sedan urminnes tider. Först och främst är detta ruttnande och jäsning. I skrifter av antika grekiska och romerska författare kan man hitta recept för att göra vin, surmjölk och bröd, vilket indikerar den utbredda användningen av jäsning i vardagen. Under medeltiden ignorerade alkemisterna inte dessa processer och studerade dem tillsammans med andra rent kemiska omvandlingar. Det var under denna period som försök gjordes att ta reda på jäsningsprocessernas karaktär.
Termen "fermentation" ("fermentatio") för att beteckna processer som involverar frigöring av gas användes först av den holländska alkemisten J.B. van Helmont (1577-1644). J. van Helmont upptäckte likheter mellan gasen som bildas under jäsningen av druvjuice (koldioxid), gasen som frigörs när kol förbränns och gasen som uppstår "när vinäger hälls på kalkstenar", dvs. när en alkali reagerar med en syra. Baserat på detta kom J. van Helmont till slutsatsen att alla ovan beskrivna kemiska omvandlingar är av samma karaktär. Senare började jäsningar särskiljas från gruppen av kemiska processer åtföljda av frigörande av gaser. För att beteckna jäsningens materiella drivkraft, dess aktiva princip, användes termen "enzym". Synen på jäsning och förruttnelse som rent kemiska processer formulerades 1697 av den tyske läkaren och kemisten G.E. Stalem (1660-1734). Enligt G. Stahls idéer är jäsning och förruttnelse kemiska omvandlingar som sker under påverkan av "enzym"-molekyler, som överför sin inneboende inre aktiva rörelse till molekylerna i det jäsbara substratet, d.v.s. fungera som en slags katalysator för reaktionen. G. Stahls åsikter om arten av sönderfalls- och jäsningsprocesserna delades fullständigt och försvarades av en av sin tids största kemister, J. Liebig. Denna synpunkt accepterades dock inte av alla forskare.
En av de första gissningarna om sambandet mellan de av Leeuwenhoek beskrivna ”globerna” (jäst) och fenomenen jäsning och förruttnelse tillhör den franske naturforskaren J.L.L. Buffon (1707-1788). Den franske kemisten A. Lavoisier (1743-1794), som kvantitativt studerade sockers kemiska omvandlingar under alkoholjäsning, kom mycket nära att förstå jästens roll i jäsningsprocessen. 1793 skrev han: "Lite bryggjäst räcker för att ge den första impulsen till jäsningen: den fortsätter sedan av sig själv. Jag kommer att rapportera någon annanstans om enzymets verkan i allmänhet." Men han misslyckades med detta: A. Lavoisier blev ett offer för den franska borgerliga revolutionens terror.
Från 30-talet av 1800-talet började en period av intensiva mikroskopiska observationer. År 1827 beskrev den franske kemisten J. Demasier (1783-1862) strukturen hos jästen Mycoderma cerevisiae, som bildar en hinna på ölytan, och, eftersom han var övertygad om att dessa är de minsta djuren, klassificerade han dem som ciliater. I J. Demasiers arbete finns det dock ingen indikation på ett möjligt samband mellan jäsningsprocessen och en film som utvecklas på ytan av den jäsande vätskan. Tio år senare gjorde den franske botanikern C. Cagnard de Latour (1777-1859) en grundlig mikroskopisk undersökning av det sediment som bildades vid alkoholjäsning och kom fram till att det består av levande varelser, vars livsviktiga aktivitet är orsaken till jäsningen. Nästan samtidigt uppmärksammade den tyske naturforskaren F. Kützing (1807-1893), när han studerade bildandet av vinäger från alkohol, en slemmassa som såg ut som en hinna på ytan av en vätska som innehåller alkohol. Genom att studera slemmassan fann F. Kützing att den består av mikroskopiska levande organismer och är direkt relaterad till ansamlingen av vinäger i miljön. En annan tysk naturforskare T. Schwann (1810-1882) kom till liknande slutsatser.
Således kom C. Cagniard de Latour, F. Kützing och T. Schwann, oberoende av varandra och nästan samtidigt, till slutsatsen om sambandet mellan jäsningsprocesser och mikroskopiska levande varelsers vitala aktivitet. Huvudslutsatsen från dessa studier formulerades tydligt av F. Kützing: ”Vi måste nu betrakta varje jäsningsprocess annorlunda än vad kemin har ansett dem hittills. Hela processen med alkoholjäsning beror på närvaron av jäst, medan ättiksyrajäsning beror på närvaron av en ättiksyramamma."
Men idéerna om den biologiska naturen hos jäsnings-"enzymet" som de tre forskarna uttryckte accepterades inte. Dessutom utsattes de för allvarlig kritik från anhängare av teorin om jäsningens fysikalisk-kemiska natur, som anklagade sina vetenskapliga motståndare för "frånvända slutsatser" och frånvaron av bevis för att stödja denna "märkliga hypotes." Teorin om jäsningsprocessernas fysikalisk-kemiska natur förblev dominerande.
2.2 Utveckling av idéer om infektionssjukdomars mikrobiella natur
Även den antika grekiske läkaren Hippokrates (ca 460-377 f.Kr.) föreslog att infektionssjukdomar orsakas av osynliga levande varelser. Avicenna (ca 980-1037) skrev i "medicinens kanon" om de "osynliga" patogenerna av pest, smittkoppor och andra sjukdomar. Liknande tankar finns i verk av den italienske läkaren, astronomen och poeten G. Fracastros (1478-1553).
Den ryske epidemiologen D.S. var djupt övertygad om att infektionssjukdomar orsakas av levande mikroskopiska varelser. Samoilovich (1744-1805), som försökte upptäcka orsaken till pesten under ett mikroskop. Han misslyckades på grund av ofullkomligheten i mikroskop och mikroskopisk teknik. Åtgärderna för desinfektion och isolering av patienter som utvecklats av D.S. Samoilovich i enlighet med hans idé visade sig dock vara mycket effektiva i kampen mot epidemier och blev allmänt kända över hela världen.
Det är värt att nämna att D. Samoilovichs samtida M. Terekhovsky (1740-1796), den första ryska protistolog-experimentern, etablerade protozoernas levande natur och 1775, för första gången i världen, tillämpade en experimentell forskningsmetod på mikroorganismer , bestämma inverkan av temperatur, elektriska urladdningar, sublimat, opium, syror och alkalier på deras livsduglighet. Genom att studera rörelse, tillväxt och reproduktion av mikroorganismer under strikt kontrollerade förhållanden, var Terekhovsky den första som påpekade att division föregås av tillväxt och en ökning i storlek. Han bevisade också omöjligheten av spontan generering av protozoer i olika kokta vätskor (infusioner). Han beskrev sina iakttagelser i sitt arbete "On Linnaeus' Pouring Chaos."
1827 upptäckte den italienske naturforskaren A. Bassi (1773-1856), när han studerade silkesmaskens sjukdom, överföringen av sjukdomen när en mikroskopisk svamp överförs från en sjuk individ till en frisk. Således var A. Bassi den första som experimentellt bevisade denna sjukdoms mikrobiella natur. Idén om den mikrobiella naturen hos infektionssjukdomar har inte fått erkännande på länge. Den dominerande teorin var att orsakerna till sjukdomar ansågs vara olika störningar i flödet av kemiska processer i kroppen.
År 1846 definierade den tyske anatomen F. Henle (1809-1885) i sin bok "Manual of Rational Pathology" tydligt de grundläggande principerna för att känna igen infektionssjukdomar. Senare underbyggdes F. Henles idéer, formulerade i allmän form (F. Henle själv kunde inte se ett enda orsakande agens för mänskliga infektionssjukdomar), experimentellt underbyggda av R. Koch och kom in i vetenskapen under namnet "Henle-Koch triad”.
3. FYSIOLOGISK PERIOD (PASTERIAN) (ANDRA HALVAN AV 1800-TALET)
3.1. Louis Pasteurs vetenskapliga verksamhet
Början av den fysiologiska perioden går tillbaka till 60-talet av 1800-talet och är förknippad med aktiviteterna hos den framstående franska vetenskapsmannen, en kemist till yrket, Louis Pasteur (1822-1895). Mikrobiologi är skyldig Pasteur inte bara dess snabba utveckling, utan också dess bildande som vetenskap. Namnet Pasteur är förknippat med de största upptäckterna som gav honom världsberömmelse: jäsning (1857), spontan generation (1860), sjukdomar i vin och öl (1865), sjukdomar hos silkesmaskar (1868), infektion och vacciner (1881), rabies (1885) .
Pasteur började sin vetenskapliga karriär med arbete med kristallografi. Han upptäckte att vid omkristallisation av salter av optiskt inaktiv racemisk syra bildas två typer av kristaller. En lösning framställd av kristaller av en typ roterar planet av polariserat ljus till höger och från kristaller av en annan typ - till vänster. Pasteur upptäckte vidare att en mögel som odlats i en lösning av racemisk vinsyra förbrukar endast en av de isomera formerna (dextrorotatory). Denna observation gjorde det möjligt för Pasteur att dra en slutsats om de specifika effekterna av mikroorganismer på substrat och fungerade som en teoretisk grund för den efterföljande studien av mikroorganismers fysiologi. Pasteurs observationer av lägre mögelsvampar väckte hans uppmärksamhet till mikroorganismer i allmänhet.
1854 fick Pasteur tjänsten som heltidsprofessor vid universitetet i Lille. Det var här han började sin mikrobiologiska forskning, som lade grunden för mikrobiologi som en självständig vetenskaplig disciplin.
Anledningen till att man började studera jäsningsprocesser var en vädjan till Pasteur från en Lille-tillverkare med en begäran om att hjälpa till att ta reda på orsakerna till systematiska misslyckanden i jäsningen av betjuice för att producera alkohol. Forskningsresultaten, som publicerades i slutet av 1857, bevisade utan tvekan att processen med alkoholjäsning är resultatet av den vitala aktiviteten hos en viss grupp av mikroorganismer - jäst och sker under förhållanden utan tillgång till luft.
Nästan samtidigt med studiet av alkoholjäsning började Pasteur studera mjölksyrajäsning och visade också att denna typ av jäsning orsakas av mikroorganismer, som han kallade "mjölksyrajäst". Pasteur presenterade resultaten av sin forskning i sina publicerade verk "Memoir on Lactic Fermentation."
Resultaten av Pasteurs forskning är faktiskt inte bara nya vetenskapliga data, de är ett djärvt vederläggande av den då dominerande teorin om jäsningens fysiska och kemiska natur, stödd och försvarad av den tidens största vetenskapliga auktoriteter: I. Berzelius, E. Mitscherlich, J. Liebig. Mjölksyrajäsning är den enklaste "kemiska" processen för nedbrytning av en sockermolekyl i två trioser, och beviset på att denna nedbrytning är förknippad med den vitala aktiviteten hos mikroskopiska organismer var ett kraftfullt argument som stödde teorin om fermenteringarnas biologiska natur.
Det andra argumentet till stöd för jäsningens biologiska natur var Pasteurs experimentella bevis på möjligheten att genomföra alkoholjäsning i ett proteinfritt medium. Enligt den kemiska teorin om jäsning är den senare resultatet av den katalytiska aktiviteten hos ett "enzym", som är ett ämne av proteinnatur.
Studiet av smörsyrajäsning ledde Pasteur till slutsatsen att livet för vissa mikroorganismer inte bara kan fortsätta i frånvaro av fritt syre, utan det senare är skadligt för dem. Resultaten av dessa observationer publicerades 1861 i en rapport med titeln "Animalculi-ciliat som lever utan fritt syre och orsakar jäsning." Upptäckten av den negativa effekten av fritt syre på processen för smörsyrajäsning var kanske den sista punkten som helt vederlagde teorin om jäsningars kemiska natur, eftersom det var syre som tilldelades rollen som föreningen som gav första impulsen till den inre rörelsen av proteinpartiklarna i "enzymet". Genom en serie studier inom jäsningsområdet bevisade Pasteur på ett övertygande sätt inkonsekvensen i den kemiska teorin om jäsningar, vilket tvingade hans motståndare att erkänna sina fel. För sitt arbete med anaerobios 1861 fick Pasteur ett pris från den franska vetenskapsakademin och en medalj från Royal Society of London. Resultatet av tjugo års forskning inom jäsningsområdet sammanfattades av Pasteur i "Forskning om öl, dess sjukdomar, deras orsaker, sätt att göra det stabilt, med tillämpning av en ny teori om jäsning" (1876).
1865 vände sig den franska regeringen till Pasteur med en begäran om att hjälpa silkesodlare som led stora förluster på grund av silkessjukdomar. Pasteur ägnade ungefär fem år åt att studera denna fråga och kom till slutsatsen att silkesmasksjukdomar orsakas av vissa mikroorganismer. Pasteur studerade i detalj sjukdomsförloppet - pebrine silkesmaskar och utvecklade praktiska rekommendationer för att bekämpa sjukdomen: han föreslog att under ett mikroskop leta efter orsakerna till sjukdomen i fjärilar och puppor, separera sjuka individer och förstöra dem, etc. .
Efter att ha fastställt den mikrobiella naturen hos infektionssjukdomar hos silkesmaskar kom Pasteur till idén att djur- och människorsjukdomar också orsakas av mikroorganismers inverkan. Hans första arbete i denna riktning var att bevisa att barnsängsfeber, utbredd under den beskrivna perioden, orsakades av en viss mikroskopisk patogen. Pasteur identifierade det orsakande medlet för feber, visade att dess orsak var försummelsen av antiseptiska regler från medicinsk personal och utvecklade metoder för skydd mot patogenens penetration i kroppen.
Pasteurs fortsatta arbete inom området för att studera infektionssjukdomar ledde till att han upptäckte orsakerna till kycklingkolera, osteomyelit, purulenta bölder och ett av de orsakande medlen för gas kallbrand. På detta sätt visade och bevisade Pasteur att varje sjukdom orsakas av en specifik mikroorganism.
1879, medan han studerade kycklingkolera, utvecklade Pasteur en metod för att erhålla kulturer av mikrober som förlorar förmågan att vara orsaken till sjukdomen, det vill säga förlora virulens, och använde denna upptäckt för att skydda kroppen från efterföljande infektion. Den senare utgjorde grunden för skapandet av teorin om immunitet.
Pasteur kombinerade sin studie av infektionssjukdomar med utvecklingen av åtgärder för att aktivt bekämpa dem. Baserat på tekniken för att erhålla försvagade kulturer av virulenta mikroorganismer, kallade "vacciner", hittade Pasteur sätt att bekämpa mjältbrand och rabies. Pasteurs vacciner blev utbredda över hela världen. Institutioner där vaccinationer mot rabies genomförs benämns Pasteurstationer för att hedra Pasteur.
Pasteurs verk uppskattades av hans samtida och fick internationellt erkännande. År 1888 byggdes ett vetenskapligt forskningsinstitut i Paris för Pasteur, med medel som samlats in genom internationell prenumeration, som för närvarande bär hans namn. Pasteur var den första direktören för detta institut. L. Pasteurs upptäckter visade hur mångfaldig, ovanlig och aktiv mikrovärlden som är osynlig för blotta ögat är och vilket enormt verksamhetsområde dess studie representerar.
3.2. Mikrobiologins utveckling under andra hälften av 1800-talet
Den franske forskaren P. Tennery bedömde de framgångar som uppnåddes av mikrobiologin under andra hälften av 1800-talet i sitt arbete "Historisk skiss över utvecklingen av naturvetenskap i Europa": "Inför bakteriologiska upptäckter, historien om andra naturvetenskapen under 1800-talets sista decennier verkar något blekare.”
Mikrobiologins framgångar under denna period är direkt relaterade till nya idéer och metodologiska tillvägagångssätt som introducerats i mikrobiologisk forskning av L. Pasteur. Bland de första som insåg betydelsen av Pasteurs upptäckter var den engelske kirurgen J. Lister, han insåg att orsaken till en stor andel av dödsfallen efter operationer är, för det första, infektion av sår med bakterier på grund av okunnighet och, för det andra, underlåtenhet att följa efterlevnaden. med grundläggande antiseptiska regler.
En av grundarna av den medicinska mikrobiologin, tillsammans med Pasteur, var den tyske mikrobiologen R. Koch (1843-1910), som studerade patogener av infektionssjukdomar. Koch började sin forskning medan han fortfarande var läkare på landsbygden med studien av mjältbrand och 1877 publicerade han ett verk ägnat åt orsaken till denna sjukdom - Bacillus anthracis. Efter detta uppmärksammades Koch på en annan allvarlig och utbredd sjukdom vid den tiden - tuberkulos. 1882 tillkännagav Koch upptäckten av tuberkulosens orsaksämne, som fick namnet "Kochs bacill" till hans ära. (År 1905 tilldelades Koch Nobelpriset för sin forskning om tuberkulos.) Koch upptäckte också orsaken till kolera 1883.
Koch ägnade stor uppmärksamhet åt utvecklingen av mikrobiologiska forskningsmetoder. Han designade en belysningsapparat, föreslog en metod för mikrofotografering av bakterier, utvecklade tekniker för att färga bakterier med anilinfärgämnen och föreslog en metod för att odla mikroorganismer på fasta näringsmedia med gelatin. Att få fram bakterier i form av rena kulturer öppnade för nya tillvägagångssätt för en mer djupgående studie av deras egenskaper och fungerade som en drivkraft för den fortsatta snabba utvecklingen av mikrobiologin. Rena kulturer av orsakande agens av kolera, tuberkulos, difteri, pest, körtlar och lobar pneumoni isolerades.
Koch underbyggde experimentellt de bestämmelser som tidigare lagts fram av F. Henle om erkännande av infektionssjukdomar, vilka kom in i vetenskapen under namnet ”Henle-Koch-triaden” (senare visade det sig dock att den inte är tillämplig på alla smittämnen).
Grundaren av rysk mikrobiologi är L. Tsenkovsky (1822-1887). Objekten för hans forskning var mikroskopiska protozoer, alger och svampar. Han upptäckte och beskrev ett stort antal protozoer, studerade deras morfologi och utvecklingscykler. Detta gjorde att han kunde dra slutsatsen att det inte fanns någon skarp gräns mellan växternas och djurens värld. Han organiserade också en av de första Pasteur-stationerna i Ryssland och föreslog ett vaccin mot mjältbrand ("levande Tsenkovsky-vaccin").
Namnet på I. Mechnikov (1845-1916) är förknippat med utvecklingen av en ny riktning inom mikrobiologi - immunologi. För första gången inom vetenskapen utvecklade Mechnikov och experimentellt bekräftade den biologiska teorin om immunitet, som gick till historien som Mechnikovs fagocytiska teori. Denna teori är baserad på idén om cellulära skyddsanordningar i kroppen. Mechnikov, i experiment på djur (daphnia, sjöstjärnelarver), visade att leukocyter och andra celler av mesodermalt ursprung har förmågan att fånga och smälta främmande partiklar (inklusive mikrober) som kommer in i kroppen. Detta fenomen, som kallas fagocytos, utgjorde grunden för den fagocytiska teorin om immunitet och har fått universellt erkännande. Mechnikov utvecklade de frågor som ställdes ytterligare och formulerade en allmän teori om inflammation som en skyddande reaktion av kroppen och skapade en ny riktning inom immunologi - läran om antigenspecificitet. För närvarande blir det allt viktigare i samband med utvecklingen av problemet med organ- och vävnadstransplantation och studiet av cancerimmunologi.
Mechnikovs viktigaste verk inom området medicinsk mikrobiologi inkluderar studier av koleras patogenes och biologin av koleraliknande vibrios, syfilis, tuberkulos och skovvis feber. Mechnikov är grundaren av doktrinen om mikrobiell antagonism, som fungerade som grunden för utvecklingen av vetenskapen om antibiotikaterapi. Idén om mikrobiell antagonism användes av Mechnikov för att utveckla problemet med livslängd. Genom att studera fenomenet åldrande av kroppen kom Mechnikov till slutsatsen. Att dess viktigaste orsak är kronisk förgiftning av kroppen med förruttnande produkter som produceras i tjocktarmen av förruttnande bakterier.
Av praktiskt intresse är Mechnikovs tidiga arbeten om användningen av svampen Isaria destructor för att bekämpa fältskadegöraren – spannmålsbaggen. De ger anledning att betrakta Mechnikov som grundaren av den biologiska metoden för att bekämpa skadedjur av jordbruksväxter, en metod som får ökande tillämpning och popularitet i dessa dagar.
Således har I.I. Mechnikov, en framstående rysk biolog som kombinerade egenskaperna hos en experimenterare, lärare och främjare av vetenskaplig kunskap, var en man med stor anda och arbete, vars högsta utmärkelse var tilldelningen av Nobelpriset till honom 1909 för sin forskning om fagocytos.
En av de största forskarna inom mikrobiologi är I. Mechnikovs vän och kollega N.F. Gamaleya (1859-1949). Gamaleya ägnade hela sitt liv åt studiet av infektionssjukdomar och utvecklingen av åtgärder för att bekämpa deras patogener. Gamaleya gjorde ett stort bidrag till studiet av tuberkulos, kolera och rabies, 1886 organiserade han tillsammans med I. Mechnikov den första Pasteur-stationen i Odessa och införde vaccination mot rabies i praktiken. Han upptäckte fågelvibrio - orsaken till koleraliknande sjukdom hos fåglar - och döpte den till Mechnikov vibrio för att hedra Ilya Ilyich. Sedan erhölls ett vaccin mot human kolera.
etc.................