Glükolüüsi tähtsus organismile. Glükolüüs. Ma olen samm. Trioosfosfaadi isomeeride moodustumine
Glükolüüs on paljude elusorganismide jaoks erinevate süsivesikute katabolismi peamine protsess. Just tema võimaldab teil genereerida energiat ATP molekulide kujul nendes rakkudes, kus fotosünteesi ei toimu. Anaeroobne glükolüüs toimub hapniku juuresolekul või puudumisel.
Protsessi eripära
Selle protsessi keemilise olemuse arvukad uuringud on näidanud, et kääritamise ja hingamise algfaasis on sarnasusi. Tänu sellele avastusele selgitasid teadlased elusmaailma ühtsust. Anaeroobne glükolüüs toimub pärast trikarboksüülhapete tsükli möödumist, vabade elektronide ülekandmist. Püruvaat siseneb mitokondritesse ja oksüdeerub süsinikdioksiidiks. See viib heksoosist vaba energia vabanemiseni. Ensüümid, mis on glükolüütiliste reaktsioonide kiirendajad, on paljudes rakkudes tsütosoolis lahustuvas vormis. Heksoosi oksüdatsiooniprotsessi kiirendajad, mis toimuvad hapniku juuresolekul, on koondunud mitokondriaalsetesse membraanidesse.
Kuut süsinikuaatomit sisaldava süsivesiku molekuli jagamiseks kaheks molekuliks on vajalik kümne aktiivse ensüümi olemasolu. Teadlastel õnnestus need puhtal kujul isoleerida, et uurida nende füüsikalisi ja keemilisi omadusi.

Protsessi etapid
Glükolüüsi protsess toimub elusorganismide rakkudes. Sellega kaasneb püroviinamarihappe moodustumine ja see koosneb mitmest etapist. Hingamisteede häirete korral tuleb glükoos aktiveerida. Sarnane protsess toimub ka süsinikuaatomi fosforüülimisel ATP-ga suhtlemisel.
glükoos + ATP = glükoos-6-fosfaat + ADP
Selle keemilise interaktsiooni läbiviimiseks kasutatakse magneesiumi katioone ja heksokinaasi (ensüümi). Järgmisena isomeriseeritakse reaktsiooniprodukt fruktoos-6-fosfaadiks. Katalüsaatorina kasutatakse ensüümi fosfoglükoisomeraasi.
Glükoosi glükolüüsi iseloomustab veel üks etapp, milles osaleb ATP. Fosforhape on seotud fruktoosi esimese süsinikuaatomiga. Glükolüüsi järgnevad etapid on seotud saadud fruktoos-1,6-difosfaadi lahtiühendamisega trioosiks, PHA (3-fosfoglütseraldehüüdi) moodustumisega.
Glükolüüs on mitmeetapiline protsess, mis on seotud energia vabanemisega. Kui üks glükoosimolekul lõhustatakse, saadakse kaks PHA molekuli, seega neid korratakse.
Glükolüüs on protsess, mis on kokku võetud võrrandiga:
C6H12O6 + 2ATP + 2NAD + 2Pn + 4ADP → 2PVK 2NADH + 2H+ + 4ATP + 2ADP

Glükolüüsi reguleerimine
Glükolüüs on elusorganismi jaoks oluline protsess. Selle eesmärk on täita kahte funktsiooni:
- ATP teke heksoosi molekulide lagunemisel;
- ehitusplokkide transport järgnevaks sünteesiprotsessiks.
Selle protsessi reguleerimine on suunatud elusraku nende vajaduste täielikule rahuldamisele. Reaktsioonid, milles ensüümid toimivad katalüsaatoritena, on pöördumatud.
Reguleeriva funktsiooni rakendamisel on eriti oluline fosfofruktokinaas. See ensüüm täidab reguleerivat funktsiooni ja iseloomustab protsessi kiirust.

Füsioloogiline tähtsus
Glükolüüsi võib pidada glükoosi katabolismi universaalseks viisiks. Seda kasutavad aktiivselt pro- ja eukarüootsed organismid. Glükolüüsi katalüüsivad ensüümid lahustuvad vees ja kogunevad tsütosooli. Mõned loomarakud ja -kuded võivad heksoosi kataboliseerida ainult glükolüüsi teel. Näiteks neerukanalite rakkudel ja aju neuronitel on sarnased võimed.
Rasvkoes ja maksas on glükolüüsi füsioloogilises rollis mõningaid erinevusi. Seedimise ajal rasvkoes ja maksas on see protsess rasvade sünteesil kasutatavate substraatide allikaks.
Mõned taimekoed säilitavad tärklist mugulates. Veetaimed saavad energiat glükolüüsi teel.
Anaeroobsetes tingimustes muudetakse püruvaat etanooliks ja laktaadiks. Protsessiga kaasneb suure hulga energia vabanemine.
Glükolüüsil on adipotsüütides oluline füsioloogiline tähtsus. Selle abiga toimub oksüdatiivse protsessi asemel lipogenees, mis vähendab oksüdatiivset stressi.

Meditsiiniline tähtsus
Anaeroobsetes tingimustes tekkiva laktaadi kuhjumisega tekib veres laktatsidoos. See viib verekeskkonna reaktsiooni vähenemiseni, millega kaasnevad teravad häired raku ainevahetuses. Sarnane protsess esineb patoloogilistes protsessides, mis on seotud kudede hapnikuvarustuse häiretega. Näiteks müokardiinfarkti, verejooksu, kopsuembooliaga. See protsess on tingitud suhkurtõvest, mille puhul aeroobse glükolüüsi asemel toimub anaeroobne protsess.
Arvestades, et insuliin on glükolüüsi kiirendaja, on I tüüpi diabeedi korral glükolüüs aeglustunud. Sellepärast täidavad need ravimid, mis stimuleerivad selles protsessis kasutatavaid ensüüme, haiguse ravimise funktsiooni.

Järeldus
Glükolüüs on protsess, mis on vajalik organismide täisväärtuslikuks eluks. Vähi korral suureneb glükoosi tarbimine kümme korda, seega sõltub kasvajarakkude elujõulisus glükolüüsist.
Pärast selle protsessi voolu omaduste üksikasjalikku uurimist õnnestus teadlastel kasutada glükolüüsi mitte ainult rakkude toitmiseks, vaid ka teatud haiguste raviks.
IN anaeroobne protsess püroviinamarihape redutseeritakse piimhappeks (laktaadiks), seetõttu nimetatakse mikrobioloogias anaeroobset glükolüüsi piimhappekääritamiseks. laktaat on metaboolne ummiktee ja siis ei muutu millekski, ainus viis laktaadi kasutamiseks on oksüdeerida see tagasi püruvaadiks.
Paljud keharakud on võimelised glükoosi anaeroobseks oksüdeerimiseks. Sest erütrotsüüdid see on ainus energiaallikas. Rakud skeletilihased tänu glükoosi hapnikuvabale lagundamisele on nad võimelised tegema võimsat, kiiret, intensiivset tööd, nagu sprint, stress jõuspordialadel. Väljaspool füüsilist pingutust paraneb hapnikuvaba glükoosi oksüdatsioon rakkudes hüpoksia ajal – erinevat tüüpi aneemia, kell vereringehäired kude, olenemata põhjusest.
glükolüüs
Anaeroobne glükoosi muundamine toimub tsütosool ja sisaldab 11 ensümaatilise reaktsiooni kahte etappi.
Glükolüüsi esimene etapp
Glükolüüsi esimene samm on ettevalmistav, siin kulub ATP energiat, aktiveeritakse glükoos ja moodustub sellest glükoos trioosfosfaadid.
Esimene reaktsioon glükolüüs taandub tsüklisse mittekuuluva 6. süsinikuaatomi fosforüülimise tõttu glükoosi muundumiseks reaktiivseks ühendiks. See reaktsioon on esimene glükoosi muundamisel, mida katalüüsib heksokinaas.
Teine reaktsioon on vaja tsüklist eemaldada veel üks süsinikuaatom selle järgnevaks fosforüülimiseks (ensüüm glükoosfosfaadi isomeraas). Selle tulemusena moodustub fruktoos-6-fosfaat.
Kolmas reaktsioon- ensüüm fosfofruktokinaas fosforüleerib fruktoos-6-fosfaati, moodustades peaaegu sümmeetrilise fruktoos-1,6-difosfaadi molekuli. See reaktsioon on glükolüüsi kiiruse reguleerimisel kesksel kohal.
IN neljas reaktsioon pooleks lõigatud fruktoos 1,6-difosfaat fruktoos-1,6-difosfaat- aldolaas kahe fosforüülitud trioosi isomeeri - aldooside - moodustumisega glütseraldehüüd(GAF) ja ketoosid dihüdroksüatsetoon(DAF).
Viies reaktsioon ettevalmistav etapp - glütseraldehüüdfosfaadi ja dihüdroksüatsetoonfosfaadi üleminek üksteiseks osalusel trioosfosfaadi isomeraas. Reaktsiooni tasakaal on nihkunud dihüdroksüatsetoonfosfaadi kasuks, selle osakaal on 97%, glütseraldehüüdfosfaadi osakaal 3%. See reaktsioon määrab kogu oma lihtsuse juures glükoosi saatuse:
- raku energiapuuduse ja glükoosi oksüdatsiooni aktiveerimisega muutub dihüdroksüatsetoonfosfaat glütseraldehüüdfosfaadiks, mis oksüdeerub glükolüüsi teises etapis edasi,
- Piisava koguse ATP-ga, vastupidi, glütseraldehüüdfosfaat isomeriseerub dihüdroksüatsetoonfosfaadiks ja viimane suunatakse rasvade sünteesi.
Glükolüüsi teine etapp

Glükolüüsi teine etapp on energia vabanemine sisaldub glütseraldehüüdfosfaadis ja säilitab selle kujul ATP.
kuues reaktsioon glükolüüs (ensüüm glütseraldehüüdfosfaatdehüdrogenaas) - glütseraldehüüdfosfaadi oksüdeerimine ja sellele fosforhappe lisamine viib 1,3-difosfoglütseriinhappe ja NADH suure energiasisaldusega ühendi moodustumiseni.
IN seitsmes reaktsioon(ensüüm fosfoglütseraadi kinaas) 1,3-difosfoglütseraadis sisalduva fosfoestersideme energia kulub ATP moodustamiseks. Reaktsioon sai täiendava nimetuse - , mis määrab energiaallika makroergiliste sidemete saamiseks ATP-s (reaktsiooni substraadist) erinevalt oksüdatiivsest fosforüülimisest (vesinikioonide elektrokeemilisest gradiendist mitokondriaalsel membraanil).
Kaheksas reaktsioon- 3-fosfoglütseraat, mis sünteesiti eelmises reaktsioonis toimel fosfoglütseraadi mutaas isomeriseerub 2-fosfoglütseraadiks.
Üheksas reaktsioon- ensüüm enolaas eraldab 2-fosfoglütseriinhappest veemolekuli ja viib fosfoenoolpüruvaadi koostises makroergilise fosfoestersideme moodustumiseni.
Kümnes reaktsioon glükolüüs on teine substraadi fosforüülimise reaktsioon- seisneb kõrge energiasisaldusega fosfaadi ülekandmises fosfoenoolpüruvaadist ADP-le püruvaatkinaasi toimel ja püruviinhappe moodustumisest.
üldine ülevaade
Glükolüütiline rada koosneb 10 järjestikusest reaktsioonist, millest igaüht katalüüsib erinev ensüüm.
Glükolüüsi protsessi võib tinglikult jagada kaheks etapiks. Esimene etapp, mis jätkub 2 ATP molekuli energiatarbimisega, on glükoosimolekuli lagunemine kaheks glütseraldehüüd-3-fosfaadi molekuliks. Teises etapis toimub glütseraldehüüd-3-fosfaadi NAD-sõltuv oksüdatsioon, millega kaasneb ATP süntees. Glükolüüs ise on täiesti anaeroobne protsess, see tähendab, et reaktsioonide toimumiseks ei ole vaja hapniku olemasolu.
Glükolüüs on üks vanimaid ainevahetusprotsesse, mida tuntakse peaaegu kõigis elusorganismides. Arvatavasti ilmnes glükolüüs rohkem kui 3,5 miljardit aastat tagasi primaarsetes prokarüootides.
Lokaliseerimine
Eukarüootsete organismide rakkudes asuvad tsütosoolis kümme ensüümi, mis katalüüsivad glükoosi lagunemist PVC-ks, kõik muud energiavahetusega seotud ensüümid on mitokondrites ja kloroplastides. Glükoos siseneb rakku kahel viisil: naatriumist sõltuv sümport (peamiselt enterotsüütide ja neerutuubulite epiteeli jaoks) ja glükoosi hõlbustatud difusioon kandevalkude abil. Nende transportervalkude tööd kontrollivad hormoonid ja ennekõike insuliin. Kõige enam stimuleerib insuliin glükoosi transporti lihas- ja rasvkoes.
Tulemus
Glükolüüsi tulemuseks on ühe glükoosi molekuli muundamine kaheks püroviinamarihappe (PVA) molekuliks ja kahe redutseeriva ekvivalendi moodustumine koensüümi NAD∙H kujul.
Glükolüüsi täielik võrrand on järgmine:
Glükoos + 2NAD + + 2ADP + 2P n \u003d 2NAD ∙ H + 2PVC + 2ATP + 2H 2O + 2H+.Hapniku puudumisel või puudumisel rakus redutseeritakse püroviinamarihape piimhappeks, siis on glükolüüsi üldvõrrand järgmine:
Glükoos + 2ADP + 2F n \u003d 2 laktaat + 2ATP + 2H 2 O.Seega on ühe glükoosimolekuli anaeroobsel lagunemisel ATP kogusaagis kaks molekuli, mis saadakse ADP substraadi fosforüülimise reaktsioonides.
Aeroobsetes organismides läbivad glükolüüsi lõpp-produktid rakuhingamisega seotud biokeemilistes tsüklites täiendavaid transformatsioone. Selle tulemusena sünteesitakse pärast ühe glükoosimolekuli kõigi metaboliitide täielikku oksüdeerumist rakuhingamise viimases etapis - hapniku juuresolekul mitokondriaalses hingamisahelas toimuv oksüdatiivne fosforüülimine - iga glükoosi jaoks täiendavalt 34 või 36 ATP molekuli. molekul.
Tee
Esimene reaktsioon glükolüüs on fosforüülimine glükoosimolekulid, mis tekib koespetsiifilise heksokinaasi ensüümi osalusel 1 ATP molekuli energiatarbimisega; moodustub glükoosi aktiivne vorm - glükoos-6-fosfaat (G-6-F):
Reaktsiooni kulgemiseks on vajalik Mg 2+ ioonide olemasolu söötmes, millega seondub ATP molekuli kompleks. See reaktsioon on pöördumatu ja on esimene glükolüüsi võtmereaktsioon.
Glükoosi fosforüülimisel on kaks eesmärki: esiteks, kuna neutraalsele glükoosimolekulile läbilaskev plasmamembraan ei lase läbi negatiivselt laetud G-6-P molekule, on fosforüülitud glükoos raku sees lukustatud. Teiseks muundatakse glükoos fosforüülimise käigus aktiivseks vormiks, mis võib osaleda biokeemilistes reaktsioonides ja olla kaasatud ainevahetustsüklitesse.
Heksokinaasi maksa isoensüüm – glükokinaas – on oluline vere glükoositaseme reguleerimisel.
Järgmises reaktsioonis ( 2 ) muundatakse fosfoglükoisomeraasi G-6-P toimel fruktoos-6-fosfaat (F-6-F):

Selle reaktsiooni jaoks ei ole vaja energiat ja reaktsioon on täielikult pöörduv. Selles etapis võib fosforüülimise teel glükolüüsi protsessi kaasata ka fruktoosi.
Seejärel järgneb peaaegu kohe üksteise järel kaks reaktsiooni: fruktoos-6-fosfaadi pöördumatu fosforüülimine ( 3 ) ja sellest tulenev pöörduv aldoolilõhe fruktoos-1,6-bisfosfaat (F-1,6-bF) kaheks trioosiks ( 4 ).
F-6-F fosforüülimine toimub fosfofruktokinaasi poolt teise ATP molekuli energiakuluga; see on teine võtmereaktsioon glükolüüs, selle reguleerimine määrab glükolüüsi intensiivsuse tervikuna.

Aldooli dekoltee F-1,6-bF esineb fruktoos-1,6-bisfosfaat-aldolaasi toimel:

Neljanda reaktsiooni tulemusena dihüdroksüatsetoonfosfaat Ja glütseraldehüüd-3-fosfaat, ja esimene on peaaegu kohe tegevuse all fosfotrioosi isomeraas läheb teise juurde 5 ), mis osaleb edasistes transformatsioonides:

Iga glütseraldehüüdfosfaadi molekul oksüdeeritakse NAD+ juuresolekul glütseraldehüüdfosfaatdehüdrogenaas enne 1,3-difosfoglütseraat (6 ):

Pärit 1,3-difosfoglütseraat, mis sisaldab makroergilist sidet 1 asendis, kannab fosfoglütseraatkinaasi ensüüm ADP molekuli üle fosforhappe jäägi (reaktsioon 7 ) - moodustub ATP molekul:

See on substraadi fosforüülimise esimene reaktsioon. Sellest hetkest alates ei ole glükoosi lagundamise protsess energia mõttes kahjumlik, kuna esimese etapi energiakulud kompenseeritakse: sünteesitakse 2 ATP molekuli (üks iga 1,3-difosfoglütseraadi kohta) kahe energiakulu asemel. reaktsioonid 1 Ja 3 . Selle reaktsiooni toimumiseks on vajalik ADP olemasolu tsütosoolis, see tähendab, et kui rakus on ATP liig (ja ADP puudus), siis selle kiirus väheneb. Kuna ATP, mis ei metaboliseeru, ei ladestu rakus, vaid lihtsalt hävib, on see reaktsioon oluline glükolüüsi regulaator.
Siis järjestikku: moodustub fosfoglütseroolmutaas 2-fosfoglütseraat (8 ):

Enolaas moodustub fosfoenoolpüruvaat (9 ):

Ja lõpuks, ADP substraadi fosforüülimise teine reaktsioon toimub püruvaadi ja ATP enoolvormi moodustumisega ( 10 ):

Reaktsioon kulgeb püruvaatkinaasi toimel. See on glükolüüsi viimane võtmereaktsioon. Püruvaadi enoolvormi isomerisatsioon püruvaadiks toimub mitteensümaatiliselt.
Alates selle loomisest F-1,6-bF ainult reaktsioonid kulgevad energia vabanemisega 7 Ja 10 kus toimub ADP substraadi fosforüülimine.
Edasine areng
Glükolüüsi käigus moodustunud püruvaadi ja NAD∙H lõplik saatus sõltub organismist ja tingimustest rakus, eriti hapniku või muude elektronide aktseptorite olemasolust või puudumisest.
Anaeroobsetes organismides kääritatakse püruvaati ja NAD∙H edasi. Piimhappekäärimise käigus näiteks bakterites redutseeritakse püruvaat ensüümi laktaatdehüdrogenaasi toimel piimhappeks. Pärmis on sarnane protsess alkoholkäärimine, mille lõppsaaduseks on etanool ja süsinikdioksiid. Tuntud on ka või- ja tsitraadikääritamine.
Võikäärimine:
Glükoos → võihape + 2 CO 2 + 2 H 2 O.
Alkohoolne kääritamine:
Glükoos → 2 etanooli + 2 CO 2.
Sidrunhappe kääritamine:
Glükoos → sidrunhape + 2 H 2 O.
Kääritamine on toiduainetööstuses hädavajalik.
Aeroobides siseneb püruvaat tavaliselt trikarboksüülhappe tsüklisse (Krebsi tsükkel) ja NAD∙H oksüdeeritakse lõpuks oksüdatiivse fosforüülimise protsessi kaudu mitokondrites hingamisahelas hapniku toimel.
Vaatamata sellele, et inimese ainevahetus on valdavalt aeroobne, täheldatakse intensiivselt töötavates skeletilihastes anaeroobset oksüdatsiooni. Piiratud hapniku juurdepääsu tingimustes muundatakse püruvaat piimhappeks, nagu toimub piimhappe fermentatsiooni käigus paljudes mikroorganismides:
PVC + NAD∙H + H + → laktaat + NAD + .
Lihasvalu, mis tekib mõni aeg pärast ebatavalist intensiivset füüsilist aktiivsust, on seotud piimhappe kogunemisega neisse.
Piimhappe moodustumine on ainevahetuse tupikharu, kuid see ei ole ainevahetuse lõpp-produkt. Laktaatdehüdrogenaasi toimel oksüdeerub piimhape uuesti, moodustades püruvaati, mis osaleb edasistes transformatsioonides.
Glükolüüsi reguleerimine
Eristada kohalikku ja üldist regulatsiooni.
Kohalik regulatsioon toimub ensüümide aktiivsuse muutmisega rakusiseste erinevate metaboliitide mõjul.
Glükolüüsi reguleerimine tervikuna, koheselt kogu organismi jaoks, toimub hormoonide toimel, mis sekundaarsete sõnumitoojate molekulide kaudu mõjutavad rakusisest ainevahetust.
Insuliin mängib olulist rolli glükolüüsi stimuleerimisel. Glükagoon ja adrenaliin on glükolüüsi kõige olulisemad hormonaalsed inhibiitorid.
Insuliin stimuleerib glükolüüsi:
- heksokinaasi reaktsiooni aktiveerimine;
- fosfofruktokinaasi stimuleerimine;
- püruvaatkinaasi stimuleerimine.
Teised hormoonid mõjutavad ka glükolüüsi. Näiteks somatotropiin pärsib glükolüüsi ensüüme ja kilpnäärmehormoonid on stimulandid.
Glükolüüsi reguleeritakse mitme põhietapi kaudu. Heksokinaasi poolt katalüüsitud reaktsioonid ( 1 ), fosfofruktokinaas ( 3 ) ja püruvaatkinaas ( 10 ) mida iseloomustab vaba energia oluline vähenemine ja need on praktiliselt pöördumatud, mis võimaldab neil olla tõhusad glükolüüsi reguleerimise punktid.
Heksokinaasi reguleerimine
Heksokinaas inhibeerib reaktsiooniprodukt - glükoos-6-fosfaat, mis seostub allosteeriliselt ensüümiga, muutes selle aktiivsust.
Kuna suurem osa rakus olevast G-6-P-st toodetakse glükogeeni lagunemisel, ei ole heksokinaasi reaktsioon tegelikult glükolüüsi toimumiseks vajalik ja glükoosi fosforüülimine glükolüüsi reguleerimisel ei ole vajalik. erakordse tähtsusega. Heksokinaasi reaktsioon on oluline samm glükoosi kontsentratsiooni reguleerimisel veres ja rakus.
Fosforüülimise käigus kaotab glükoos võime transportida läbi membraani kandemolekulide abil, mis loob tingimused selle akumuleerumiseks rakus. Heksokinaasi G-6-P inhibeerimine piirab glükoosi sisenemist rakku, vältides selle liigset akumuleerumist.
Glükoos-6-fosfaat ei inhibeeri maksa glükokinaasi (heksokinaasi IV isotüüp) ja maksarakud jätkavad glükoosi akumuleerumist isegi suure G-6-P sisalduse korral, millest seejärel sünteesitakse glükogeen. Võrreldes teiste isotüüpidega on glükokinaasil kõrge Michaelise konstandi väärtus, see tähendab, et ensüüm töötab täisvõimsusel ainult kõrge glükoosikontsentratsiooni tingimustes, mis esineb peaaegu alati pärast sööki.
Glükoos-6-fosfaati saab glükoos-6-fosfataasi toimel tagasi glükoosiks muuta. Ensüümid glükokinaas ja glükoos-6-fosfataas osalevad normaalse veresuhkru taseme säilitamises.
Fosfofruktokinaasi reguleerimine
Fosfofruktokinaasi reaktsiooni intensiivsus mõjutab kogu glükolüüsi läbilaskevõimet ja fosfofruktokinaasi stimuleerimist peetakse regulatsiooni kõige olulisemaks sammuks.
Fosfofruktokinaas (PFK) on tetrameerne ensüüm, mis eksisteerib vaheldumisi kahes konformatsioonilises olekus (R ja T), mis on tasakaalus ja liiguvad vaheldumisi ühest teise. ATP on nii PFK substraat kui ka allosteeriline inhibiitor.
Igal FFK subühikul on kaks ATP sidumissaiti: substraadi sait ja inhibeerimissait. Substraadi sait on võrdselt võimeline kinnitama ATP-d mis tahes tetrameeri konformatsioonis. Kui inhibeerimiskoht seob ATP-d ainult siis, kui ensüüm on konformatsioonilises olekus T. Teine FPA substraat on fruktoos-6-fosfaat, mis kinnitub ensüümiga eelistatavalt R-olekus. ATP kõrge kontsentratsiooni korral on inhibeerimiskoht hõivatud, üleminekud ensüümide konformatsioonide vahel muutuvad võimatuks ja enamik ensüümi molekule stabiliseerub T-olekus, ei suuda P-6-P-d siduda. ATP fosfofruktokinaasi inhibeerimist aga pärsib AMP, mis kinnitub ensüümi R-konformatsioonidele, stabiliseerides seega ensüümi oleku F-6-P sidumiseks.
Glükolüüsi ja glükoneogeneesi kõige olulisem allosteeriline regulaator on fruktoos 2,6-bisfosfaat, mis ei ole nende tsüklite vahelüli. Fruktoos-2,6-bisfosfaat aktiveerib allosteeriliselt fosfofruktokinaasi.
Fruktoos-2,6-bifosfaadi sünteesi katalüüsib spetsiaalne bifunktsionaalne ensüüm - fosfofruktokinaas-2 / fruktoos-2,6-bifosfataas (FFK-2 / F-2,6-BPaas). Fosforüülimata kujul on valk tuntud kui fosfofruktokinaas-2 ja sellel on katalüütiline toime fruktoos-6-fosfaadile, mis toodab fruktoos-2-6-bisfosfaati. Selle tulemusena stimuleeritakse oluliselt FFK aktiivsust ja inhibeeritakse tugevalt fruktoos-1,6-bifosfataasi aktiivsust. See tähendab, et FFK-2 aktiivsuse tingimustes nihkub selle reaktsiooni tasakaal glükolüüsi ja glükoneogeneesi vahel esimese poole - sünteesitakse fruktoos-1,6-bisfosfaat.
Fosforüülitud vormis bifunktsionaalsel ensüümil ei ole kinaasi aktiivsust, vastupidi, selle molekulis aktiveerub sait, mis hüdrolüüsib P2,6BP P6P-ks ja anorgaaniliseks fosfaadiks. Bifunktsionaalse ensüümi fosforüülimise metaboolne toime seisneb selles, et PFK allosteeriline stimulatsioon peatub, F-1,6-BPaasi allosteeriline inhibeerimine elimineeritakse ja tasakaal nihkub glükoneogeneesi suunas. Tekib F6F ja seejärel glükoos.
Bifunktsionaalse ensüümi interkonversioonid teostab cAMP-sõltuv proteiinkinaas (PC), mida omakorda reguleerivad veres ringlevad peptiidhormoonid.
Kui glükoosi kontsentratsioon veres väheneb, pärsitakse ka insuliini moodustumist ja glükagooni vabanemist, vastupidi, stimuleeritakse ja selle kontsentratsioon veres tõuseb järsult. Glükagoon (ja teised kontrainsulaarhormoonid) seonduvad maksarakkude plasmamembraani retseptoritega, põhjustades membraani adenülaattsüklaasi aktivatsiooni. Adenülaattsüklaas katalüüsib ATP konversiooni tsükliliseks AMP-ks. cAMP seondub proteiinkinaasi regulatoorse alaühikuga, põhjustades selle katalüütiliste subühikute vabanemise ja aktiveerimise, mis fosforüülivad mitmeid ensüüme, sealhulgas bifunktsionaalset FFK-2/P-2,6-BPaasi. Samal ajal peatub glükoosi tarbimine maksas ning glükoneogenees ja glükogenolüüs aktiveeruvad, taastades normoglükeemia.
püruvaadi kinaas
Järgmine samm, kus glükolüüsi reguleerimine toimub, on viimane reaktsioon - püruvaatkinaasi toime etapp. Püruvaatkinaasi puhul on kirjeldatud ka mitmeid isoensüüme, millel on reguleerivad omadused.
Maksa püruvaadi kinaas(L-tüüpi) reguleerib fosforüülimine, allsteerilised efektorid ja geeniekspressiooni reguleerimine. Ensüümi inhibeerivad ATP ja atsetüül-CoA ning aktiveerib fruktoos-1,6-bisfosfaat. ATP püruvaatkinaasi inhibeerimine toimub sarnaselt ATP toimega PFK-le. ATP seondumine ensüümi inhibeerimise kohaga vähendab selle afiinsust fosfoenoolpüruvaadi suhtes. Maksa püruvaadi kinaas fosforüülitakse ja inhibeeritakse proteiinkinaasi poolt ning on seega ka hormonaalse kontrolli all. Lisaks reguleeritakse maksa püruvaadi kinaasi aktiivsust ka kvantitatiivselt, st muutes selle sünteesi taset. See on aeglane, pikaajaline regulatsioon. Süsivesikute sisalduse suurenemine toidus stimuleerib püruvaatkinaasi kodeerivate geenide ekspressiooni, mille tulemusena tõuseb ensüümi tase rakus.
M-tüüpi püruvaadi kinaas ajus, lihastes ja teistes glükoosinõudvates kudedes leiduvat ei reguleeri proteiinkinaas. See on põhiline, kuna nende kudede metabolismi määravad ainult sisemised vajadused ja see ei sõltu veresuhkru tasemest.
Lihaspüruvaatkinaas ei allu välismõjudele, nagu veresuhkru taseme langus või hormonaalne vabanemine. Ekstratsellulaarsed seisundid, mis põhjustavad fosforüülimist ja maksa isoensüümi inhibeerimist, ei muuda M-tüüpi püruvaatkinaasi aktiivsust. See tähendab, et glükolüüsi intensiivsus vöötlihastes on määratud ainult rakusiseste tingimustega ja ei sõltu üldisest regulatsioonist.
Tähendus
Glükolüüs on erakordse tähtsusega kataboolne rada. See annab energiat rakuliste reaktsioonide jaoks, sealhulgas valkude sünteesiks. Glükolüüsi vaheprodukte kasutatakse rasvade sünteesil. Püruvaati saab kasutada ka alaniini, aspartaadi ja muude ühendite sünteesimiseks. Tänu glükolüüsile ei piira mitokondrite jõudlus ja hapniku kättesaadavus lühiajaliste ekstreemsete koormuste korral lihasjõudu.
Vaata ka
Lingid
- glükolüüs (inglise keeles)
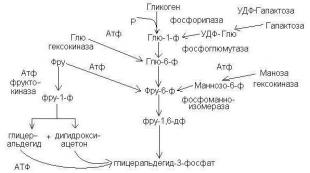
Anaeroobne glükolüüs on keeruline ensümaatiline protsess, mis koosneb glükoosi järjestikustest muundumistest, mis toimub inimese ja looma kudedes ilma hapnikutarbimiseta (joonis 28).
Püruviinhappe pöörduvat muundamist piimhappeks katalüüsib laktaatdehüdrogenaas:
Glükolüüsi kogutulemust väljendatakse järgmise võrrandiga: C 6 H 12 O 6 + 2H 3 RO 4 + 2ADP \u003d 2C 3 H 6 O 3 + 2ATP + 2H 2 O
Seega on ATP puhassaagis anaeroobse glükolüüsi ajal 2 mol ATP 1 mooli glükoosi kohta. Just tänu anaeroobsele glükolüüsile saab inimese ja looma organism hapnikupuuduse tingimustes teatud aja jooksul täita mitmeid füsioloogilisi funktsioone.
Seda protsessi bakterites nimetatakse piimhappekääritamiseks: see on fermenteeritud piimatoodete valmistamise aluseks. Anaeroobne glükolüüs toimub rakkude tsütosoolis, mis sisaldab kõiki selleks vajalikke ensüüme ja ei vaja mitokondriaalset hingamisahelat. ATP moodustub anaeroobse glükolüüsi protsessis substraadi fosforüülimise reaktsioonide tõttu.
Anaeroobsetes tingimustes pärmis toimub sarnane protsess - alkoholkäärimine, sel juhul püroviinamarihape dekarboksüülitakse, moodustades atseetaldehüüdi, mis seejärel redutseeritakse etüülalkoholiks:
CH3-CO-COOH → CH3-CHO + CO2;
CH 3 -CHO + OVERHAND + H + → CH 3 -CH 2 -OH + ÜLE +.

Joonis 28. Anaeroobse glükoosi glükolüüsi skeem
10.6. Glükoosi aeroobne lagunemine
Glükoosi aeroobne lagunemine koosneb kolmest etapist:
1) glükoosi muundamine püruviinhappeks (püruvaadiks) – aeroobne glükolüüs. See osa sarnaneb ülalpool käsitletud anaeroobse glükolüüsi protsessiga, välja arvatud selle viimane etapp (püruvaadi muundamine piimhappeks);
2) katabolismi üldine tee;
3) mitokondriaalne elektronide transpordiahel - kudede hingamise protsess.
Katabolismi üldine rada
Katabolismi üldine tee koosneb kahest etapist.
1. etapp - püroviinamarihappe oksüdatiivne dekarboksüülimine. See on keeruline mitmeetapiline protsess, mida katalüüsib mitmest ensüümist koosnev süsteem – püruvaatdehüdrogenaasi kompleks; lokaliseeritud mitokondrites (sisemembraan ja maatriks) ja seda saab väljendada üldise üldskeemiga:
CH 3 -CO-COOH + HS-KoA + ÜLE + → CH 3 -CO-SkoA + OVERHAND + H + + CO 2.
2. etapp – Krebsi tsükkel (tsitraaditsükkel ehk trikarboksüül- ja dikarboksüülhapete tsükkel) (joonis 29); lokaliseeritud mitokondrites (maatriksis). Selles tsüklis moodustab atsetüül-CoA-s sisalduv atsetüüljääk mitmeid primaarseid vesiniku doonoreid. Lisaks siseneb vesinik dehüdrogenaaside osalusel hingamisahelasse. Tsitraaditsükli ja hingamisahela konjugeeritud toime tulemusena oksüdeeritakse atsetüüljääk CO 2 -ks ja H 2 O-ks. Aeroobse lagunemise käigus toimuva glükoosi muundumiste kogu jada üldvõrrand on järgmine:
C6H12O2 + 6O2 → 6CO2 + 6H2O
Aeroobse lagunemise energiamõju on 38 ATP molekuli süntees 1 glükoosimolekuli lagunemise käigus. Seega energia osas on glükoosi täielik oksüdeerimine süsinikdioksiidiks ja veeks tõhusam protsess kui anaeroobne glükolüüs. Hapnik pärsib anaeroobset glükolüüsi, seetõttu täheldatakse liigse hapniku juuresolekul taimede ja loomade kudedes üleminekut anaeroobselt glükolüüsilt (käärimine) hingamisele (aeroobne glükolüüs), s.o. rakkude üleminek tõhusamale ja ökonoomsemale energia saamise viisile (Pasteuri efekt). Anaeroobse glükolüüsi roll organismi energiaga varustamisel on eriti suur lühiajalise intensiivse töö ajal, mil hapniku transportimise mehhanismi võimsusest mitokondritesse ei piisa aeroobse glükolüüsi läbiviimiseks. Niisiis tagab ~ 30 sekundi (200 m) jooksmise täielikult anaeroobne glükolüüs, samal ajal kui anaeroobse glükolüüsi kiirus väheneb suurenenud hingamisega ja aeroobse lagunemise kiirus suureneb. Pärast 4-5 min. jooksmine (1,5 km) - poole energiast annab anaeroobne, poole aeroobne protsess. Pärast 30 min. (10 km jooks) – energia tarnitakse peaaegu täielikult aeroobse protsessi abil.
Punastel verelibledel ei ole üldse mitokondreid ja nende vajadus ATP järele rahuldatakse täielikult anaeroobse glükolüüsi abil.
(kreeka keelest glykys - magus ja lüüs - lagunemine, lagunemine) - üks kolmest peamisest (glükolüüs, Krebsi tsükkel ja Entner-Doudorovi tee) elusorganismides energia tootmise viisist. See on loomsetes kudedes süsivesikute (peamiselt glükoosi ja glükogeeni) anaeroobne (see tähendab, et see ei nõua vaba O 2 osalemist) ensümaatilise mittehüdrolüütilise lagunemise protsess, millega kaasneb adenosiintrifosforhappe (ATP) süntees ja mis lõpeb piimhappe moodustumine. Glükolüüs on oluline lihasrakkudele, spermatosoididele, kasvavatele kudedele (ka kasvajatele), sest tagab energia salvestamise hapniku puudumisel. Kuid glükolüüsi tuntakse ka O 2 juuresolekul (aeroobne glükolüüs) - erütrotsüütides, võrkkestas, loote kudedes vahetult pärast sündi ja soole limaskestas. G. ja K. Corey, aga ka sellised biokeemia pioneerid nagu O. Meyerhof ja G. Embden andsid suure panuse glükolüüsi uurimisse. Glükolüüs oli esimene täielikult dešifreeritud biokeemiliste reaktsioonide jada (19. sajandi lõpust 1940. aastateni). Heksomonofosfaadi šundi või pentoosfosfaadi rada mõnes rakus (erütrotsüüdid, rasvkude) võib samuti täita energiatarnija rolli.
Lisaks glükoosile võivad glükolüüsi protsessis osaleda glütserool, mõned aminohapped ja muud substraadid. Lihaskoes, kus glükolüüsi põhisubstraadiks on glükogeen, algab protsess reaktsioonidega 2 ja 3 ( cm. skeem) ja seda nimetatakse glükogenolüüsiks. Tavaline glükogenolüüsi ja glükolüüsi vaheühend on glükoos-6-fosfaat. Glükogeeni moodustumise vastupidist rada nimetatakse glükogeneesiks.
Glükolüüsi käigus tekkivad saadused on substraadid järgnevateks oksüdatiivseteks muundumisteks ( cm. Trikarboksüülhappe tsükkel või Krebsi tsükkel). Glükolüüsiga sarnased protsessid on piimhappe, võihappe, alkoholi, glütserooli fermentatsioon, mis toimub taime-, pärmi- ja bakterirakkudes. Glükolüüsi üksikute etappide intensiivsus sõltub happesusest - pH - pH (optimaalne pH 7–8), temperatuurist ja söötme ioonsest koostisest. Glükolüüsi reaktsioonide järjestus ( cm. skeem) on hästi uuritud ja vaheproduktid on kindlaks tehtud. Rakumahlas sisalduvad lahustuvad glükolüüsi ensüümid on eraldatud kristalsel või puhastatud kujul.
Ensüümid, mis viivad läbi glükolüüsi üksikuid etappe:
1. Heksokinaas CF2.7.1.1 (või glükokinaas CF2.7.1.2)
2. Glükogeenfosforülaas CP2.4.1.1
3. Fosfoglukomutaas CP2.7.5.1
4. Glükoosfosfaat isomeraas CP5.3.1.9
5. Fosfofruktokinaas KF2.7.1.11
6. Fruktoosbisfosfaat-aldolaas EC4.1.2.13
7. Trioosfosfaadi isomeraas CP5.3.1.1
8, 9. Glütseraldehüüdfosfaatdehüdrogenaas CP1.2.1.12
10. Fosfoglütseraatkinaas CP2.7.2.3
11. Fosfoglütsermutaas EC2.7.5.3
12. Enolase KF4.2.1.11
13. Püruvaatkinaas KF2.7.1.40
14. Laktaatdehüdrogenaas CP1.1.1.27
Glükolüüs algab suhkrute fosfori derivaatide moodustumisega, mis aitab kaasa substraadi tsüklilise vormi muutumisele atsükliliseks, reaktiivsemaks. Üks glükolüüsi kiirust reguleerivatest reaktsioonidest on reaktsioon 2, mida katalüüsib ensüüm fosforülaas. Keskne reguleeriv roll glükolüüsis kuulub ensüümile fosfofruktokinaasile (reaktsioon 5), mille aktiivsust inhibeerivad ATP ja tsitraat, kuid stimuleerivad selle lagunemissaadused. Glükolüüsi keskseks lüliks on glükolüütiline oksidoreduktsioon (reaktsioonid 8–10), mis on redoksprotsess, mis kulgeb 3-fosfoglütseraldehüüdi oksüdeerumisega 3-fosfoglütseriinhappeks ja koensüümi n(NAD) redutseerimisega. Need transformatsioonid viiakse läbi 3-fosfoglütseradehüdrogenaasi (DPHA) abil fosfoglütseraatkinaasi osalusel. See on glükolüüsi ainus oksüdatiivne etapp, kuid see ei vaja vaba hapnikku, vajalik on ainult NAD + olemasolu, mis seejärel redutseeritakse NAD-H2-ks.
Oksüdoreduktsiooni (redoksprotsessi) tulemusena vabaneb energia, mis akumuleerub (energiarikka ATP ühendi kujul) substraadi fosforüülimise protsessis. Teine reaktsioon, mis tagab ATP moodustumise, on reaktsioon 13 – püroviinamarihappe moodustumine. Anaeroobsetes tingimustes lõpeb glükolüüs piimhappe moodustumisega (reaktsioon 14) laktaatdehüdrogenaasi toimel ja redutseeritud NAD osalusel, mis oksüdeeritakse NAD-ks (NAD-H 2) ja mida saab taas kasutada oksüdatiivses etapis. . Aeroobsetes tingimustes oksüdeerub püroviinamarihape mitokondrites Krebsi tsükli ajal.
Seega, kui laguneb 1 molekul glükoosist, moodustub 2 molekuli piimhapet ja 4 molekuli ATP-d. Samal ajal kulub glükolüüsi esimestes etappides (vt reaktsioonid 1, 5) 2 ATP molekuli 1 glükoosimolekuli kohta. Glükogenolüüsi käigus moodustub 3 ATP molekuli, kuna. glükoos-6-fosfaadi saamiseks pole vaja kulutada ATP-d. Glükolüüsi esimesed üheksa reaktsiooni on selle endergooniline (energia neeldumisega) faas ja viimased reaktsioonid on selle eksergooniline (energia vabanemisega) faas. Glükolüüsi käigus vabaneb ainult umbes 7% teoreetilisest energiast, mida saab glükoosi täielikul oksüdatsioonil (CO 2 ja H 2 O-ks). Üldine energia salvestamise efektiivsus ATP kujul on aga 35–40% ja praktilistes tingimustes võivad rakud olla isegi kõrgemad.
Glütseraldehüüdfosfaatdehüdrogenaas ja laktaatdehüdrogenaas on sisemiselt seotud (üks vajab NAD +, teine moodustab NAD +), mis tagab selle koensüümi ringluse. Võib-olla on see terminaalse dehüdrogenaasi peamine biokeemiline tähtsus.
Kõik glükolüüsireaktsioonid on pöörduvad, välja arvatud 1, 5 ja 13. Glükoosi (reaktsioon 1) või fruktoosmonofosfaati (reaktsioon 5) saab aga saada nende fosfori derivaatidest fosforhappe hüdrolüütilise lõhustamise teel sobivate ensüümide juuresolekul; reaktsioon 13 on praktiliselt pöördumatu, ilmselt fosforrühma suure hüdrolüüsienergia tõttu (umbes 13 kcal/mol). Seetõttu toimub glükoosi moodustumine glükolüüsi saadustest erineval viisil.
O 2 juuresolekul glükolüüsi kiirus väheneb (Pasteuri efekt). Mõnedes intensiivselt glükolüüsivates kudedes on näiteid kudede hingamise pärssimisest glükolüüsiga (Crabtree efekt). Anaeroobsete ja aeroobsete oksüdatiivsete protsesside vaheliste seoste mehhanisme ei mõisteta täielikult. Glükolüüsi ja glükogeneesi protsesside samaaegne reguleerimine määrab ainulaadselt süsiniku voolu mõlemal teel, sõltuvalt keha vajadustest. Kontroll viiakse läbi kahel tasandil - hormonaalne (kõrgematel loomadel regulatoorsete kaskaadide kaudu teise sõnumitoojate osalusel) ja metaboolne (kõigis organismides).
Igor Rapanovitš