Bakteriyalar səltənətinin canlı orqanizmlərinin biosferdəki rolu. Xülasə: Prokariotların kükürd dövranında iştirakı Dərsin istinad nöqtələri
Sual 1. Prokariotların biosferdəki rolunu təsvir edin.
Prokaryotlar fotosintez həyata keçirir və bununla da atmosferə oksigen çıxarırlar. Prokaryotlar arasında kemosintez geniş yayılmışdır. Bundan əlavə, bakterial orqanizmlər arasında azot fiksasiya edən formalar var: bu, planetimizdə azotu birbaşa atmosfer havasından udmaq qabiliyyətinə malik olan və bununla da molekulyar azotu bioloji dövrəyə cəlb edən yeganə canlı orqanizmlər qrupudur.
Prokariotların başqa mühüm funksiyası da var: üzvi birləşmələrin məhv edilməsi (minerallaşması) yolu ilə qeyri-üzvi maddələrin ətraf mühitə qaytarılması.
Bütövlükdə biosfer səviyyəsində prokaryotlar, ilk növbədə bakteriyalar, başqa bir çox vacib funksiyaya malikdir - konsentrasiya. Onlar hətta son dərəcə aşağı konsentrasiyalarda da ətraf mühitdən müəyyən elementləri aktiv şəkildə çıxara bilirlər.
Prokaryotların xassələri və funksiyaları o qədər müxtəlifdir ki, prinsipcə onlar davamlı fəaliyyət göstərən ekosistemlər yaratmağa qadirdirlər (yəni yalnız onların iştirakı ilə).
Sual 2. Çirkləndiricilərlə mübarizə aparmaq üçün prokaryotik dərmanların təbii mühitə daxil edilməsinin təhlükəsinin nə olduğunu izah edin.
Prokaryotları ətraf mühitə gətirərkən ortaya çıxan problemli məsələlərdən biri yerli mikroflora ilə təqdim olunan arasında əlaqənin xarakterini müəyyən etməkdir. Mikrobiosenozlarda mövcud balansın dəyişməsi nəticəsində introduksiya edilmiş mikroorqanizmlərin təbii biogeosenozların vəziyyətinə mənfi təsir göstərməsi ehtimalı istisna oluna bilməz. Bu vəziyyət böyük ehtiyatlılıq tələb edir və yalnız diqqətli tədqiqatlardan sonra prokaryotlardan insan ehtiyacları üçün və biosferi çirkləndiricilərlə mübarizədə istifadə etmək lazımdır.
Sual 3. Məlumdur ki, bütün ən müxtəlif azot fiksasiya edən bakteriyalar molekulyar azotun bağlanmasını kataliz edən eyni fermentə, nitrogenazaya malikdir. Bu faktın canlı təbiətin təkamülündə hansı əhəmiyyəti var?
Nitrogenaza iki zülal kompleksindən ibarət çoxölçülü bir fermentdir. Müxtəlif azot fiksatorlarından olan nitrogenazlar molekulyar çəkilərinə və metal tərkibinə görə bir qədər fərqlənir. Bu fakt azot fiksasiya edən bakteriyaların mənşəyinin birliyini göstərir. Nitrogenaza bakteriyaların ata-baba formasında meydana çıxdı, sonra divergensiya baş verdi, bu da azot fiksasiya edən bakteriyaların müxtəlifliyinə səbəb oldu, hər birində nitrogenazı kodlayan bir gen var idi.
Azotun fiksasiyası prosesi normal atmosfer təzyiqində və normal temperaturda baş verir, lakin çox enerji tələb edir.
Nitrogenazanın aktiv işləməsi üçün mikroaerofil şərait tələb olunur. Molekulyar oksigen hər iki nitrogenaz zülalına zərərli təsir göstərir. Düyündəki mikroaerofil şərait oksigen üçün diffuz maneə ilə təmin edilir. Havadan ammonyak istehsal edən nitrogenaz kompleksi çox qənaətcil fəaliyyət göstərir. Yaşayış yerində kifayət qədər ammonium və ya nitrat ionları varsa, o, fəaliyyətini dayandırır.
Azot bütün canlı orqanizmlər üçün tamamilə vacib elementdir. Azotun əsas anbarı yerin atmosferidir. Eukaryotik orqanizmlər azotu birbaşa atmosferdən qəbul edə bilmirlər. Azot fiksasiya edən bakteriyalarda molekulyar azotun bağlanmasını kataliz edən nitrogenaz fermentinin olması səbəbindən onlarla simbiozda yaşayan bitkilər asanlıqla əldə etdikləri azotu əldə edə bilirlər. Bir çox bitki, heyvan və göbələk azot fiksasiya edən prokaryotlarla simbioza girmək qabiliyyətinə malikdir.
Sual 4. 70-ci illərin əvvəllərində amerikalı bioloq və ekoloq Barri Kommoner. XX əsr tezisini ekologiya qanunu kimi irəli sürdü: “Ən yaxşısını təbiət bilir”. Nə demək istədiyinizi izah edin. Bu məsələ ilə bağlı fikrinizi bildirin. Cavabınızı misallarla izah edin.
Barry Commoner deyir ki, təbiətin mexanizmləri və funksiyaları haqqında tamamilə etibarlı məlumatlar olana qədər, biz, saatın quruluşu ilə tanış olmayan, lakin onu düzəltmək istəyən bir insan kimi, təbii sistemləri təkmilləşdirməyə çalışarkən asanlıqla zərər veririk. O, həddindən artıq ehtiyatlı olmağa çağırır. Təbiətin çevrilməsi iqtisadi cəhətdən fəlakətli və ekoloji cəhətdən təhlükəlidir. Nəhayət, həyat üçün yararsız şərait yarana bilər. Təkmilləşdirmə üçün ekoloji meyarlar göstərilmədən təbiətin abadlaşdırılması ilə bağlı mövcud rəy mənasızdır.
Mən bu fikirlə razıyam. Çox vaxt təbiətin bir təzahürünün biosenozlar, biogeosenozlar və ya bütövlükdə biosfer üçün daha dağıdıcı və zərərli hadisənin nəticəsi olduğunu düşünmürük. "Əsas odur ki, özümüzü yaxşı hiss edirik" və təbiət gözləyəcək. Bunu belə edə bilməzsiniz!!!
Canavarların "meşə nizamlıları" olduğu ortaya çıxan güllələnməsinə misallar verə bilərsiniz. Çində sərçələr məhv edilirdi, hansı ki, guya əkinləri məhv edir, lakin heç kim quşsuz əkinlərin zərərli həşəratlar tərəfindən məhv ediləcəyini düşünmürdü.
Bioloji təkamüldə ən mühüm hadisələrdən biri olan eukaryotik hüceyrənin görünüşüdür. Eukaryotik və prokaryotik orqanizmlər arasındakı fərq genomun tənzimlənməsinin daha inkişaf etmiş bir sistemidir. Və bunun sayəsində təkhüceyrəli orqanizmlərin uyğunlaşma qabiliyyəti, genomda irsi dəyişikliklərə məruz qalmadan dəyişən ətraf mühit şəraitinə uyğunlaşma qabiliyyəti artmışdır. Uyğunlaşma qabiliyyəti sayəsində eukariotlar çoxhüceyrəli ola bildi - çoxhüceyrəli orqanizmdə eyni genomlu hüceyrələr şəraitdən asılı olaraq morfologiyası və funksiyası ilə tamamilə fərqli toxumalar əmələ gətirir.
Bu aromorfoz, yalnız eukaryotik hüceyrələrə xas olan steroid birləşmələrinin qalıqları - biomarkerlər tərəfindən müəyyən edilən arxey və proterozoyun (2,6 - 2,7 milyard il əvvəl) növbəsində meydana gəldi. Eukariotların görünüşü oksigen inqilabı ilə üst-üstə düşür.
Ümumiyyətlə qəbul edilir ki, eukariotlar bir neçə növ prokariotun simbiozu nəticəsində yaranmışdır. Göründüyü kimi, mitoxondriya alfa-proteobakteriyalardan (aerob eubakteriyalar), plastidlər siyanobakteriyalardan, sitoplazma isə naməlum arxebakteriyalardan əmələ gəlmişdir. Nüvənin, sitoskeletin və flagellanın mənşəyi haqqında ümumi qəbul edilmiş bir nəzəriyyə hələ mövcud deyil. Yer üzündə həyatın mənşəyi haqqında fərziyyələr hüceyrənin mənşəyi məsələsinə aydınlıq gətirmədi. Prokaryotların mənşəyi ilə bağlı onların meydana gəlməsini inandırıcı şəkildə təsvir edən heç bir fərziyyə yoxdursa, eukaryotik hüceyrələrin mənşəyinə gəldikdə, bir neçə fikir var.
Eukariotların mənşəyinin əsas fərziyyələri:
1. Simbiotik hipotez iki anlayışa əsaslanır. Bu anlayışlardan birincisinə görə, canlı təbiətdəki ən əsas fərq bakteriya ilə həqiqi nüvəli hüceyrələrdən ibarət olan orqanizmlər - protistlər, heyvanlar, göbələklər və bitkilər arasındakı fərqdir. İkinci konsepsiya ondan ibarətdir ki, eukaryotik hüceyrələrin bəzi hissələrinin mənşəyi simbiozların təkamülü - müxtəlif növ orqanizmlər arasında daimi birləşmələrin formalaşması idi. Güman edilir ki, üç sinif orqanellər - mitoxondriyalar, kirpiklər və fotosintetik plastidlər, simbioz nəticəsində prokaryotik ev sahibi hüceyrələrə daxil edilmiş sərbəst yaşayan bakteriyalardan yaranmışdır. Bu nəzəriyyə daha çox Mendel genetikasını Darvinin təbii seçmə ideyası ilə əlaqələndirən genetiklər, ekoloqlar və sitoloqlar tərəfindən hazırlanmış neo-Darvinist fikirlərə əsaslanır. O, həmçinin molekulyar biologiyadan, xüsusən də zülalların və amin turşusu ardıcıllığının strukturu haqqında məlumatlardan, Yerdəki həyatın ən erkən izlərini öyrənən mikropaleontologiyadan və bu elmlər bioloji mənşəli qazlarla məşğul olduğu üçün atmosfer fizikası və kimyasından istifadə edir.
2.İnvaginasiya hipotezi eukaryotik hüceyrənin əcdad formasının aerob prokaryot olduğunu deyir. İçəridə hüceyrə membranına bağlı bir neçə genom var idi. Korpuskulyar orqanoidlər və nüvə qabığın hissələrinin invaginasiyası və bağlanması, ardınca nüvəyə, mitoxondriyaya və xloroplastlara funksional ixtisaslaşma ilə yarandı. Sonra təkamül prosesində nüvə genomu mürəkkəbləşdi və sitoplazmatik membranlar sistemi meydana çıxdı. Bu fərziyyə qabıqlarda nüvənin, mitoxondrilərin, xloroplastların və iki membranın olmasını izah edir. Lakin o, korpuskulyar orqanoidlərdə və eukaryotik hüceyrənin sitoplazmasında zülal biosintezi prosesinin təfərrüatları arasındakı fərqləri izah etməkdə çətinliklərlə qarşılaşır. Mitoxondriya və xloroplastlarda bu proses müasir prokaryotik hüceyrələrdəki prosesə tam uyğun gəlir.
Simbiotik (I) və invaginasiya (II) hipotezlərinə görə eukaryotik hüceyrənin mənşəyi:
1 - anaerob prokaryot (ev sahibi hüceyrə), 2 - mitoxondriyalı prokaryotlar, 3 - mavi-yaşıl yosunlar (ehtimal xloroplast), 4 - siirochaetebənzər bakteriya (ehtimal flagellum), 5 - bayraq hüceyrəli primitiv eukariot, 6 - bitki 7 - bayraqcıqlı heyvan hüceyrəsi, 8 - aerob prokaryot (ehtimal mitoxondriya), 9 - aerob prokaryot (II fərziyyəyə görə əcdad hüceyrə), 10 - nüvə və mitoxondriyanın yaranmasına səbəb olan hüceyrə membranının invaginasiyası, 11 - primitiv eukariot 12 - xloroplastı verən hüceyrə membranının invaginasiyası, 13 - bitki hüceyrəsi; a - prokaryotik hüceyrənin DNT-si, b - mitoxondri, c - eukaryotik hüceyrənin nüvəsi, d - flagellum, e - xloroplast.
Mövcud məlumatlar fərziyyələrdən birinə üstünlük vermək və ya əksər elm adamlarına uyğun gələn yenisini hazırlamaq üçün hələ kifayət deyil, lakin son illərdə eukaryotik hüceyrənin mənşəyinin simbiogenetik nəzəriyyəsini inandırıcı şəkildə sübut etmək mümkün olmuşdur.
Eukaryotik hüceyrələrin təkamül qabiliyyəti prokaryotik hüceyrələrdən daha yüksəkdir. Burada aparıcı rol prokariotların genomundan daha böyük olan eukariotların nüvə genomuna aiddir. Əhəmiyyətli fərqlər nüvələrdə iki gen dəstinin olması səbəbindən eukaryotik hüceyrələrin diploidliyi, həmçinin bəzi genlərin dəfələrlə təkrarlanmasıdır.
Hüceyrə fəaliyyətinin tənzimlənməsi mexanizmi getdikcə mürəkkəbləşir ki, bu da tənzimləyici genlərin nisbi sayının artmasında və prokaryotların dairəvi “çılpaq” DNT molekullarının DNT-nin zülallarla əlaqəli olduğu xromosomlarla əvəz edilməsində özünü göstərir.
Aerob tənəffüs də çoxhüceyrəli formaların inkişafı üçün ilkin şərt kimi xidmət edirdi. Atmosferdə O 2 konsentrasiyası 1%-ə (Paster nöqtəsi) çatdıqdan sonra eukaryotik hüceyrələrin özləri Yerdə peyda oldular. Və bu konsentrasiya aerob tənəffüs üçün zəruri şərtdir.
Məlumdur ki, hər bir eukaryotik hüceyrədə müxtəlif mənşəli genomlar var: heyvan və göbələk hüceyrələrində bunlar nüvənin və mitoxondriyanın genomlarıdır, bitki hüceyrələrində isə bunlar da plastid genomlardır. Kiçik dairəvi DNT də eukaryotik hüceyrələrin flagellasının bazal gövdəsində olur.
Molekulyar saat metoduna görə eukariotlar prokariotlarla eyni vaxtda yaranmışdır. Ancaq aydındır ki, Yer tarixinin əhəmiyyətli bir hissəsində prokaryotlar üstünlük təşkil edirdi. Eukaryotik ölçülərə uyğun gələn ilk hüceyrələrin (akritarxların) 3 milyard yaşı var, lakin onların təbiəti hələ də aydın deyil. Eukariotların demək olar ki, müəyyən qalıqlarının təxminən 2 milyard il yaşı var. Və yalnız oksigen inqilabından sonra planetin səthində eukariotlar üçün əlverişli şərait yarandı (təxminən 1 milyard il əvvəl).
Çox güman ki, eukaryotik hüceyrələrin əsas əcdadı qida hissəciklərini qəbul edərək qidalanmaya keçən arxebakteriyalardır. Belə qəbul üçün lazım olan hüceyrə formasının dəyişməsi aktin və miyozindən ibarət olan sitoskeleton tərəfindən təmin edilmişdir. Belə bir hüceyrənin irsi aparatı membranla əlaqəsini qoruyub saxlayaraq, dəyişən səthindən daha da dərinləşmişdir. Və bu, artıq nüvə məsamələri olan bir nüvə qabığının görünüşünə səbəb oldu.
Ev sahibi hüceyrə tərəfindən udulmuş bakteriyalar onun içərisində mövcud olmağa davam edə bilər. Beləliklə, mitoxondrilərin əcdadları bir qrup fotosintetik bakteriyalara - bənövşəyi alfaproteobakteriyalara çevrildi. Ev sahibi hüceyrənin içərisində onlar fotosintez qabiliyyətini itirmiş və üzvi maddələrin oksidləşməsini öz üzərinə götürmüşlər. Onların sayəsində eukaryotik hüceyrələr aerob oldu. Digər fotosintetik hüceyrələrlə simbiozlar bitki hüceyrələrinin plastidlər almasına səbəb oldu. Mümkündür ki, eukaryotik hüceyrələrin flagellası ev sahibi hüceyrələr və qıvrılan hərəkətlərə qadir olan bakteriyalar arasındakı simbioz nəticəsində yaranmışdır.
Eukaryotik hüceyrələrin irsi aparatı prokariotlarla təxminən eyni quruluşda idi. Lakin daha böyük və daha mürəkkəb bir hüceyrəni idarə etmək zərurətindən sonra xromosomların təşkili sonradan dəyişdi və DNT histon zülalları ilə əlaqələndirildi. Prokaryotik təşkilat hüceyrədaxili simbionların genomlarında qorunub saxlanılır.
Müxtəlif simbiogenez aktları nəticəsində eukaryotik orqanizmlərin müxtəlif qrupları yaranmışdır: eukaryotik hüceyrə + siyanobakteriyalar = qırmızı yosunlar; eukaryotik hüceyrə + proklorofit bakteriya = yaşıl yosunlar. Hətta qızılı, diatom, qəhvəyi və kriptomonad yosunlarının xloroplastları iki ardıcıl simbioz nəticəsində yaranıb, bunu 4 membranın olması sübut edir.
Eukariotların görünüşü, biosfer tarixində şəraitin xüsusilə qeyri-sabit və gözlənilməz olduğu, prokaryotların adaptiv strategiyasının (sürətli mutasiya, üfüqi gen mübadiləsi və davamlı klonların seçilməsi) çox israfçı olduğu bir dövrlə məhdudlaşdı. kifayət qədər təsirli. Belə bir vəziyyətdə məqsədəuyğun modifikasiya dəyişkənliyinin inkişafına əsaslanan prinsipial olaraq daha universal və iqtisadi uyğunlaşma strategiyası böyük üstünlük əldə edə bilər.
Ola bilsin ki, eukariotların əmələ gəlməsi və onların cinsi prosesinin inkişafı dəyişkənliyin və biomüxtəlifliyin strukturunu daha diskret və “idarə olunan” etdi - bu, biomüxtəlifliyin sürətlə artmasına və növlərin təkamül plastikliyinin və ekoloji dözümlülüyünün artmasına səbəb olmalı idi. , icmalar və bütövlükdə biota.
Eukaryotların görünüşünü haqlı olaraq "standart" aromorfoz adlandırmaq olar. Bu hadisə bioloji təkamülün ümumi mütərəqqi istiqamətini açıq şəkildə nümayiş etdirdi. Tərəqqi təkcə təşkilatın mürəkkəbləşməsi, həyatın ümumi adaptiv zonasının genişlənməsi, biokütlənin və sayların artması, orqanizmlərin muxtariyyətinin artması ilə yanaşı, həm də canlı sistemlərin dayanıqlığının artırılmasında özünü göstərirdi.
Eukariotların nümunəsindən istifadə etməklə aydın şəkildə göstərilir ki, yeni həyat formalarının yaranması bəzi ayrı-ayrı filetik xətlərin və ya təbəqələrin təkamülü nəticəsində deyil, daha yüksək nizamlı sistemlərin inkişafının təbii və qaçılmaz təsiri kimi qəbul edilməlidir. - icmalar, biosfer və bəlkə də bütövlükdə bütün planet.
İstifadə olunan mənbələr:
A. V. Markov, A. M. Kulikov. Mikrob icmasında inteqrasiya prosesləri nəticəsində eukariotların mənşəyi
A.V.Markov. Eukariotların mənşəyi problemi
M. V. Larina. Eukaryotik hüceyrələrin mənşəyi haqqında fərziyyələr. Çoxhüceyrəliliyin yaranması
Dərsin növü - birləşdirilmiş
Metodlar: qismən axtarış, problemin təqdimatı, izahlı və illüstrativ.
Hədəf:
Şagirdlərdə canlı təbiət, onun sistemli təşkili və təkamülü haqqında vahid biliklər sisteminin formalaşdırılması;
Bioloji məsələlər üzrə yeni məlumatlara əsaslandırılmış qiymət vermək bacarığı;
Vətəndaş məsuliyyətinin, müstəqilliyin, təşəbbüskarlığın tərbiyəsi
Tapşırıqlar:
Təhsil: bioloji sistemlər haqqında (hüceyrə, orqanizm, növ, ekosistem); canlı təbiət haqqında müasir təsəvvürlərin inkişaf tarixi; biologiya elmində görkəmli kəşflər; dünyanın müasir təbiətşünaslıq mənzərəsinin formalaşmasında biologiya elminin rolunu; elmi biliyin metodları;
İnkişafümumbəşəri insan mədəniyyətinə daxil olan biologiyanın görkəmli nailiyyətlərinin öyrənilməsi prosesində yaradıcılıq qabiliyyətləri; müxtəlif informasiya mənbələri ilə iş prosesində müasir elmi baxışların, ideyaların, nəzəriyyələrin, konsepsiyaların, müxtəlif fərziyyələrin (həyatın, insanın mahiyyəti və mənşəyi haqqında) inkişafının mürəkkəb və ziddiyyətli yollarını;
tərbiyə canlı təbiəti bilmək imkanına, təbii mühitə və öz sağlamlığına qayğı göstərmək ehtiyacına inam; bioloji problemləri müzakirə edərkən rəqibin fikrinə hörmət
Biologiyanın öyrənilməsinin şəxsi nəticələri:
1. rus vətəndaş şəxsiyyətinin tərbiyəsi: vətənpərvərlik, Vətənə sevgi və hörmət, Vətənə qürur hissi; öz etnik mənsubiyyətini bilmək; çoxmillətli rus cəmiyyətinin humanist və ənənəvi dəyərlərinin mənimsənilməsi; Vətən qarşısında məsuliyyət və borc hissini tərbiyə etmək;
2. öyrənməyə məsuliyyətli münasibətin formalaşdırılması, öyrənmə və bilik motivasiyası, şüurlu seçim və dünyadakı oriyentasiyaya əsaslanan gələcək fərdi təhsil trayektoriyasının qurulması əsasında tələbələrin özünü inkişaf və özünü təhsilə hazırlığı və bacarığı. davamlı idrak maraqları nəzərə alınmaqla peşələr və peşəkar üstünlüklər;
Biologiyanın tədrisinin metafənni nəticələri:
1. öyrənmə məqsədlərini müstəqil müəyyənləşdirmək, öyrənmə və idrak fəaliyyətində özü üçün yeni məqsədlər qoymaq və formalaşdırmaq, idrak fəaliyyətinin motivlərini və maraqlarını inkişaf etdirmək bacarığı;
2. tədqiqat və layihə fəaliyyətinin komponentlərini mənimsəmək, o cümlədən problemi görmək, sual vermək, fərziyyələr irəli sürmək bacarığı;
3. müxtəlif bioloji məlumat mənbələri ilə işləmək bacarığı: müxtəlif mənbələrdə (dərslik mətni, elmi-kütləvi ədəbiyyat, bioloji lüğətlər və məlumat kitabçaları) bioloji məlumatları tapmaq, təhlil etmək və
məlumatı qiymətləndirmək;
Koqnitiv: bioloji obyektlərin və proseslərin əsas xüsusiyyətlərinin müəyyən edilməsi; insanlar və məməlilər arasındakı əlaqənin sübutunu (arqumentasiyasını) təmin etmək; insanlar və ətraf mühit arasında əlaqələr; insan sağlamlığının ətraf mühitin vəziyyətindən asılılığı; ətraf mühitin qorunması ehtiyacı; biologiya elminin metodlarına yiyələnmək: bioloji obyektlərin və proseslərin müşahidəsi və təsviri; bioloji təcrübələrin qurulması və onların nəticələrinin izahı.
Tənzimləyici: məqsədə çatmaq yollarını, o cümlədən alternativləri müstəqil şəkildə planlaşdırmaq, təhsil və idrak problemlərinin həllinin ən təsirli yollarını şüurlu şəkildə seçmək bacarığı; müəllim və həmyaşıdları ilə təhsil əməkdaşlığı və birgə fəaliyyəti təşkil etmək bacarığı; fərdi və qrupda işləmək: mövqelərin əlaqələndirilməsi və maraqların nəzərə alınması əsasında ümumi həll yolu tapmaq və münaqişələri həll etmək; informasiya-kommunikasiya texnologiyalarından istifadə sahəsində səriştələrin (bundan sonra İKT səlahiyyətləri) formalaşdırılması və inkişafı.
Ünsiyyətcil: həmyaşıdları ilə ünsiyyət və əməkdaşlıqda kommunikativ səriştənin formalaşması, yeniyetməlik dövründə gender sosiallaşmasının xüsusiyyətlərini dərk etmək, sosial faydalı, təhsil və tədqiqat, yaradıcılıq və digər fəaliyyət növləri.
Texnologiyalar : Sağlamlığın qorunması, problem əsaslı, inkişaf etdirici təhsil, qrup fəaliyyətləri
Texnikalar: təhlil, sintez, nəticə çıxarma, məlumatın bir növdən digərinə tərcüməsi, ümumiləşdirmə.
Təqdimat hostinqi
"Prokaryotların kükürd dövranında iştirakı"
Nijni Novqorod 2010
Giriş
Prokaryotlar (bakteriyalar və arxeya) Yerdəki həyat üçün müstəsna əhəmiyyətə malikdir - onlar həyat üçün zəruri olan əsas elementlərin (karbon, oksigen, azot, kükürd, fosfor) tsiklik çevrilməsində əsas rol oynayırlar. Canlı orqanizmlərin toplu şəkildə qurulduğu elementlərin tsiklik çevrilmələri maddələrin dövranını təmsil edir. İndi mübahisəsiz şəkildə sübut edilmişdir ki, dövrlərin müəyyən mərhələləri yalnız prokaryotlar tərəfindən həyata keçirilir, biosferdəki əsas biogen elementlərin dövrlərinin bağlanmasını təmin edir. V.İ. Vernadski (biosfer doktrinasının banilərindən biri), "biosfer həyatın qabığını - canlı maddənin mövcudluğu bölgəsini təmsil edir".
1. Kükürd dövrü
Kükürd biogen elementdir, canlı maddənin zəruri komponentidir. Zülallarda amin turşularının bir hissəsi kimi olur, zülallarda kükürdün miqdarı 0,8-2,4% təşkil edir. Kükürd həmçinin vitaminlərin, qlikozidlərin, koenzimlərin tərkibinə daxildir və bitki efir yağlarında olur. Kükürd yer qabığında, kömürlərdə, şistlərdə, neftlərdə və təbii qazlarda çox olur.
Kükürd dəyişən valentliyə malik bir elementdir. Bu, onun hərəkətliliyini təmin edir. Qeyri-üzvi birləşmələr şəklində kükürd oksidləşmiş formada (sulfatlar, politiyonatlar), azaldılmış formada (sulfidlər) və molekulyar formada baş verir, aktiv redoks dövrü keçirir. Təbiətdə kükürd müxtəlif kimyəvi və bioloji çevrilmələrə məruz qalır, qeyri-üzvi birləşmələrdən üzvi birləşmələrə və geriyə doğru hərəkət edərək, valentliyini – 2 ilə +6 arasında dəyişir.
Kükürd birləşmələrinin siklik çevrilmələrinə kükürd dövrü deyilir.
Kükürdün çevrilmə dövrünə oksidləşdirici və reduksiya vahidləri, həmçinin kükürdün valentliyini dəyişmədən çevrilməsi daxildir. Kükürd dövrünün oksidləşdirici hissəsi şəraitdən asılı olaraq həm sırf kimyəvi, həm də orqanizmlərin, əsasən mikroorqanizmlərin iştirakı ilə baş verə bilən mərhələləri əhatə edir (bunlar çılpaq gözlə görünməyən orqanizmlər, o cümlədən mikroskopik eukaryotlar: göbələklər, yosunlar, protozoa və bütün prokaryotlar). Kükürd dövrünün azaldıcı hissəsi bu prosesdə prokaryotların dominant rolu ilə əsasən bioloji olaraq həyata keçirilir, kükürd atomu isə maksimum oksidləşmə vəziyyətindən (+ 6) maksimum reduksiyaya (-2) bərpa olunur. Bununla belə, bu proses həmişə başa çatmır və ətraf mühitdə çox vaxt natamam oksidləşmiş məhsullara rast gəlinir: elementar kükürd, politiyonatlar, sulfit.
Beləliklə, kükürd dövranı, eləcə də maddələrin dövranı dövrün bağlanmasını təmin edən prokariotların iştirakı olmadan mümkün deyil.
2. Bərpaedici filial
Sulfat assimilyasiyası.
Sulfat demək olar ki, bütün bitkilər və mikroorqanizmlər tərəfindən kükürd mənbəyi kimi istifadə olunur. Assimilyasiya zamanı sulfat azalır ki, kükürd üzvi birləşmələrə daxil ola bilsin, çünki canlı orqanizmlərdə kükürd demək olar ki, yalnız sulfhidril (-SH) və ya disulfid (-S-S-) qrupları şəklində azaldılmış formada olur. Hər iki halda, yalnız orqanizmin inkişafı üçün lazım olan qədər kükürd tərkibli qida maddələri mənimsənilir, buna görə də kükürd mübadiləsinin azaldılmış məhsulları ətraf mühitə buraxılmır. Biosintez nəticəsində kükürd əsasən kükürdlü amin turşularının tərkibinə daxildir: sistin, sistein, metionin. Kükürd tərkibli üzvi maddələrin tərkibində sulfatların iştirakına assimilyativ sulfat reduksiyası deyilir.
H 2 S əmələ gəlməsi ilə üzvi kükürd birləşmələrinin çevrilməsi.
Kükürd canlı orqanizmlər üçün əsasən həll olunan sulfatlar və ya azaldılmış üzvi kükürd birləşmələri şəklində mövcuddur.
Üzvi kükürd tərkibli birləşmələrin minerallaşması zamanı kükürd qeyri-üzvi reduksiya edilmiş formada H 2 S şəklində ayrılır. Üzvi kükürd tərkibli birləşmələrdən (canlıların metabolik məhsulları) kükürdün ayrılmasında ammonifikasiya qabiliyyətinə malik saprofit mikroorqanizmlər iştirak edir. , ölü bitki və heyvan qalıqları). Ammonifikasiya zamanı kükürd tərkibli zülallar və nuklein turşuları parçalanaraq CO 2, karbamid, üzvi turşular, aminlər və kükürd dövranı üçün vacib olan H 2 S və merkaptanlar (tioalkollar) əmələ gətirir. Merkaptanlar da aerob şəraitdə oksidləşərək H2S-i buraxırlar.
Zülalların mikroorqanizmlər tərəfindən məhv edilməsi hüceyrədənkənar bir proses kimi başlayır. Bu zaman zülallar protolitik ekzofermentlər vasitəsilə hüceyrəyə nüfuz edə bilən daha kiçik molekullara hidrolizə olunur və hüceyrədaxili proteazlar tərəfindən daha da parçalana bilən amin turşularına parçalanır.
Sülfat və elementar kükürddən H 2 S-nin birbaşa formalaşması.
Biosferdə hidrogen sulfidin əmələ gəlməsi əsasən qlobal kükürd dövranı üçün böyük əhəmiyyət kəsb edən sulfat reduksiya edən bakteriyaların fəaliyyəti ilə bağlıdır. Sulfat reduksiya edən bakteriyalar dissimilyasiya edən sulfat reduksiyasını həyata keçirir, bu anaerob tənəffüsdür, burada sulfat üzvi maddələrin və ya molekulyar hidrogenin oksidləşməsi zamanı son elektron qəbuledicisi (oksigen əvəzinə) kimi xidmət edir. Buna görə də, sulfat azaldan bakteriyalarda metabolizmin enerji növünə çox vaxt sulfat tənəffüsü deyilir. Sxematik olaraq dissimilyasiya edən sulfat reduksiyası zamanı sulfat reduksiya prosesini aşağıdakı kimi göstərmək olar: SO 4 2- →SO 3 2- →S 3 O 6 2- →S 2 O 3 2- →S 2-.
Sulfat reduksiyasında iştirak edən enzimatik sistem iki hissədən ibarətdir: birincisi, ATP-dən asılı prosesdə sulfatı sulfitə, ikincisi isə altı elektron ötürülməsi ilə sulfiti sulfidə azaldır. Hüceyrəni enerji ilə təmin edən, dissimilyasiya edən sonuncu reaksiyadır.
Sulfat reduksiya edən bakteriyalar əsasən məcburi anaerob bakteriyalardır. Sulfat reduksiya edən bakteriyaların geokimyəvi rolu olduqca böyükdür, çünki onların fəaliyyəti sayəsində anaerob zonada inert birləşmə sulfat bioloji kükürd dövrəsində geniş miqyasda iştirak edir.
Sulfatı azaldan bakteriyaların fəaliyyəti gölməçələrin və çayların dibində, bataqlıqlarda və dəniz sahillərində lildə xüsusilə nəzərə çarpır. Dəniz suyunda sulfat konsentrasiyası nisbətən yüksək olduğundan, sulfat reduksiyası dənizin dayazlıqlarında üzvi maddələrin minerallaşmasında mühüm amildir. Belə minerallaşmanın əlamətləri H 2 S qoxusu və bu prosesin baş verdiyi qara lildir. Şlamın qara rəngi onun tərkibində çoxlu miqdarda qara dəmir sulfidinin olması ilə əlaqədardır. Üzvi maddələrin toplanması xüsusilə intensiv sulfat azalmasına səbəb olan bəzi sahilyanı ərazilər H 2 S-in zəhərli təsirləri səbəbindən praktiki olaraq cansızdır.
Sulfat reduksiya edən bakteriyalar sistematik deyil, fizioloji qrupdur, çünki onlara bir fizioloji prosesi - sulfatların iştirakı ilə anaerob tənəffüsü, məsələn, Desulfovibrio (vibrio) cinsinin bakteriyalarını həyata keçirməyə qadir olan müxtəlif taksonomik qruplardan olan bakteriyalar daxildir. Desulfotomaculum (spor əmələ gətirən çubuqlar). Arxeylərə sulfat reduksiya edən orqanizmlər arasında da rast gəlinmişdir. Sulfat reduksiya edən bakteriyalar təkcə sulfatları deyil, həm də tiosulfat, sulfit, elementar kükürd və digər kükürd birləşmələrini azaldaraq inkişaf edə bilər.
Bəzi sulfat azaldan bakteriyalar üçün kükürd mübadiləsinin əsasən yeni növü göstərilmişdir. Bu bakteriyalar üzvi substratlarda böyüdükdə təkcə sulfat reduksiyası hesabına deyil, həm də tiosulfat, sulfit, ditionitin sulfat və sulfid əmələ gəlməsi ilə qeyri-mütənasibliyi nəticəsində enerji əldə edə bilirlər.
Ümumi tiosulfat disproporsiya reaksiyası:
S 2 O 3 2- + H 2 0 → SO 4 2- + HS - + H +
Get = – 21,9 kJ/mol S 2 O 3 2-
Ümumi sulfit disproporsiya reaksiyası:
4SO 3 2- + H + → 3SO 4 2- + HS -
Get = – 58,9 kJ/molSO 3 2-
Müəyyən edilmişdir ki, dəniz çöküntülərində qeyri-mütənasib tiosulfat olan bakteriyalar geniş yayılmışdır.
Elementar kükürdün reduksiyası zamanı hidrogen sulfid də əmələ gələ bilər. Bu gün molekulyar kükürddən hidrogen sulfidin əmələ gəlməsinin iki mexanizmi məlumdur. Birinci halda, bakteriya və arxeylər molekulyar kükürddən anaerob tənəffüs zamanı (dissimilyasiya kükürdün azaldılması) elektron qəbuledicisi kimi istifadə edirlər, bu zaman ATP sintez olunur. Dissimilyativ kükürd reduksiyası həm mezofil, həm də termofilik prokaryotlar tərəfindən həyata keçirilən enzimatik prosesdir. İkinci halda, mikroorqanizmlər (maya və prokaryotlar) kükürddən yalnız fermentasiya zamanı buraxılan elektronları atmaq üçün istifadə edirlər (asanlaşdırılmış fermentasiya). Bu, ATP sintezi ilə müşayiət olunmayan elektronların boş (boş) boşalmasıdır.
Molekulyar kükürdün hidrogen sulfid halına salınması xüsusilə elementar kükürdün kükürdün ən əhəmiyyətli formalarından biri olduğu və kükürd birləşmələrinin ilkin olaraq vulkanik mənşəli olduğu hidrotermlərdə yaşayan termofilik mikroorqanizmlər üçün böyük əhəmiyyət kəsb edir. Hidrotermal biosenozlar canlıların unikal icmalarıdır. Yüksək temperaturda (45-50-dən 100 ° C-ə qədər) inkişaf edərək, əsasən prokaryotlar - bakteriya və arxeylər tərəfindən əmələ gəlir. Hidrotermal mayelərin mikrob icmalarını təşkil edən mikroorqanizmlərin böyük əksəriyyəti başqa yerlərdə tapılmır. Hidrotermlərin mikrob icmaları Yer kürəsinin ən qədim biosenozları hesab olunur.
3. Oksidləşdirici budaq
Kükürd dövrünün bu hissəsi tamamilə qeyri-üzvi kükürd birləşmələrinin reaksiyalarından ibarət ola bilər: S 2- → nS 2- → S 0 → S 2 O 3 2- → SO 3 2- → SO 4 2-, həmçinin üzvi reaksiyalar daxildir. formaları. Üzvi sulfidlərin kükürd atomu ümumiyyətlə S2 şəklində ayrıldıqdan sonra oksidləşir - qeyri-üzvi yolla, baxmayaraq ki, kükürd atomu üzvi birləşmələrin bir hissəsi olaraq oksidləşdikdə, məsələn, sistein kimi sırf üzvi oksidləşmə yolu da mümkündür. → sistin. Kükürd birləşmələrinin oksidləşmə reaksiyalarının əksəriyyəti güclü oksidləşdirici maddələrin (H 2 O 2, O 3, oksigen radikalları) iştirakı ilə mikroorqanizmlərin iştirakı olmadan baş verə bilər, lakin mikrob oksidləşməsi xüsusilə reagentin aşağı konsentrasiyalarında daha təsirli olur.
Kükürd birləşmələrini oksidləşdirə bilən mikroorqanizmlər arasında üç əsas fizioloji qrup fərqləndirilir: anaerob fototrof bakteriyalar, aerob və fakultativ anaerob litotrof bakteriyalar, arxeya və müxtəlif heterotrof mikroorqanizmlər.
Anaerob fototrof bakteriyalar.
Bu, bitkilərdə və siyanobakteriyalarda aerob (oksigen) fotosintezdə olduğu kimi, H 2 O əvəzinə elektron donorları kimi müxtəlif azaldılmış kükürd birləşmələrindən istifadə edərək, anaerob (anoksigen) fotosintez həyata keçirən xüsusi bir bakteriya qrupudur. Azaldılmış kükürd birləşmələrinin ən çox yayılmış oksidləşmə məhsulları SO 4 2- və S 0-dır, sonuncu bəzi fototrof bakteriyalarda hüceyrədaxili toplana bilər.
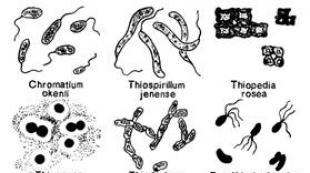
Sistematik olaraq fototrof bakteriyalar bir neçə qrupa bölünür: kükürdlü və kükürdsüz bənövşəyi və yaşıl bakteriyalar, heliobakteriyalar, eritrobakteriyalar (şək. 123). Fototrof bakteriyalar piqmentlidir və qəhvəyi, yaşıl və ya bənövşəyi rəngli ola bilər. Anoksigen fotosintezi həyata keçirən fototrof bakteriyaların tərkibində onlara uyğun rəng verən bakterioxlorofil və karotenoidlər var. Bu bakteriyalar kifayət qədər işığın nüfuz etdiyi göllər, gölməçələr, laqunlar və mineral bulaqlar kimi dayaz su hövzələrinin anaerob su sütununda sulfid oksidləşməsində üstünlük təşkil edir. İstisna, məcburi aeroblar olan və aerob şəraitdə anoksigen fotosintez aparan eritrobakteriyalardır.

Litotrofik kükürd oksidləşdirici bakteriyalar və arxeylər. Kükürd bakteriyaları və tionik bakteriyalar.
Bəzi prokaryotlar azalmış kükürd birləşmələrinin iştirakı ilə litotrofik böyüməyə qadirdir. Litotrofik böyümə zamanı azalmış kükürd birləşmələri və ya digər reduksiya edilmiş qeyri-üzvi birləşmələr (NH 3, H 2, CO, Fe 2+) enerji mübadiləsində elektron donordur, yəni elektron daşıma zəncirində oksidləşdikdə ATP sintez olunur. Yalnız prokaryotlar litotrofik böyüməyə qadirdir. Litotrofik kükürddən asılı prokaryotlarda azalmış kükürd birləşmələrinin oksidləşmə reaksiyaları enerji mənbəyi kimi xidmət edir. Oksidləşmə reaksiyaları xüsusi ferment sistemlərinin iştirakı ilə həyata keçirilir. Müxtəlif kükürd birləşmələrinin və kükürdün özünün litotrof prokaryotlar tərəfindən oksidləşməsi adətən sulfatların əmələ gəlməsinə səbəb olur. Lakin bu proses heç də həmişə başa çatmır və mühitdə çox vaxt natamam oksidləşmiş məhsullara (S 0, SO 3 2-, S 2 O 3 2-, S 4 O 6 2-) rast gəlinir. Litotrof kükürddən asılı prokaryotlar qrupuna əsasən tion bakteriyalarının nümayəndələri, həddindən artıq termoasidofil kükürddən asılı olan arxeylər, hidrogen bakteriyaları, rəngsiz kükürd bakteriyaları, həmçinin kükürd birləşmələrinin oksidləşməsi nəticəsində litotrofik böyüməyə qadir olan bənövşəyi bakteriyaların bəzi nümayəndələri daxildir. qaranlıq.
Çoxdan məlumdur ki, hidrogen sulfid bulaqlarında və tərkibində hidrogen sulfid olan digər su hövzələrində, bir qayda olaraq, hüceyrələrində kükürd damcıları olan çoxlu sayda rəngsiz mikroorqanizmlərə rast gəlinir. Hidrogen sulfidin konsentrasiyasının nisbətən aşağı olduğu yerlərdə (50 mq/l-dən az) rəngsiz kükürd bakteriyaları adlanan belə mikroorqanizmlər çox vaxt plyonkalar, ağ çöküntülər və digər çirklənmələr şəklində kütləvi yığılmalar əmələ gətirirlər. S.N. Winogradsky (1887) sübut etdi ki, kükürd bakteriyalarının tipik nümayəndələrindən birinin, yəni Beggiatoa-nın hüceyrələrində çökən kükürd hidrogen sulfiddən əmələ gəlir və bu mikroorqanizm tərəfindən sulfat turşusuna qədər oksidləşə bilər. Sistematik olaraq, rəngsiz kükürd bakteriyaları heterojen bir qrupdur və təsvir olunan növlərin və hətta nəsillərin hamısını möhkəm qurulmuş hesab etmək olmaz. Morfologiyasına, hərəkətinin təbiətinə, çoxalma üsuluna və hüceyrə quruluşuna görə rəngsiz kükürdlü bakteriyaların həm çoxhüceyrəli, həm də birhüceyrəli (Beggiatoa, Thiothrix, Thiospirillopsis, Thioploca, Achromatium) bir sıra nümayəndələri mavi-yaşıl yosunlarla böyük oxşarlıq göstərirlər. Bəzi tədqiqatçılar, xüsusən də Pringsheim (1963) bu mikroorqanizmləri onların rəngsiz variantları hesab edirlər. Mavi-yaşıl yosunlar Oscillatoria, Thiothrix - Rivularia, Thiospirillopsis - Spirulina Beggiatoa-nın analoqları hesab olunur və Achromatium Synechococcus-a bənzəyir. Mavi-yaşıl yosunlar indi bakteriya kimi təsnif olunduğundan, onların rəngsiz kükürd bakteriyaları ilə əlaqəsi getdikcə daha çox əsaslandırılır. Onu da qeyd etmək lazımdır ki, bəzi mavi-yaşıl yosunların hüceyrələrində kükürd çökdürdüyü sübut edilmişdir, baxmayaraq ki, bu xüsusiyyət tək başına mikroorqanizmlərin taksonomiyasını çox təmin etmir. Rəngsiz kükürd bakteriyalarına aid olan mikroorqanizmlərə həm şirin, həm də duzlu su hövzələrində rast gəlinir. Onların bəziləri aşağı temperaturda yaxşı inkişaf edir, digərləri (Thiospirillum pistiense) 50°C-dən yuxarı temperaturda termal kükürdlü bulaqlarda inkişaf edir. Mobil formalarda kemotaksis var və optimal oksigen və hidrogen sulfid tərkibi olan yerlərə keçə bilir.
Tionik bakteriyaların əksəriyyəti tipik kemoautotroflardır, yəni. azaldılmış kükürd birləşmələrini təkcə H-donorları kimi deyil, həm də enerji mənbələri kimi istifadə edirlər və karbon qazını assimilyasiya edərək sırf mineral mühitlərdə inkişaf edə bilirlər. Belə bakteriyalar ilk dəfə Neapol körfəzinin suyundan təcrid olunmuş (Nathanson, 1902) və Thiobacillus (Beijerinck, 1904) adını almışdır. Bu günə qədər müxtəlif su obyektlərindən, torpaqdan, həmçinin kükürd və müxtəlif metal yataqlarından təcrid olunmuş avtotrof tiobasillərin bir çox növləri təsvir edilmişdir.
Bəzi, əsasən fizioloji xüsusiyyətlərə əsasən (müxtəlif kükürd birləşmələrini və digər qeyri-üzvi substratları oksidləşdirmək, üzvi birləşmələrdən istifadə etmək, ətraf mühitin pH dəyərlərindən, oksigenə münasibətdən və s. asılı olaraq böyümək qabiliyyəti), əhəmiyyətli sayda tiobasil növləri fərqləndirilir. Bunlardan aşağıdakılar ən möhkəm qurulmuş hesab olunur: Thiobacillus thioparus, T. thiooxidans, T. denitrificans, T. thiocyanoxidans, T. neapolitanus, T. intermedius, T. novellus, T. ferrooxidanas. Bütün bu mikroorqanizmlər bikarbonatın tez-tez əlavə olunduğu sadə mineral mühitlərdə böyüyür. Azotun mənbəyi adətən ammonium duzlarıdır. Bəzi növlər və suşlar nitratları azot mənbəyi kimi mənimsəməyə qadirdir. Ayrı-ayrı nümayəndələrin karbamid və amin turşularından istifadə etmək qabiliyyəti də göstərilmişdir. Duzlu su obyektlərindən təcrid olunmuş suşlar böyümə üçün patrium xlorid tələb edir və doymuş NaCl məhlulunda böyüyə bilən halofillər məlumdur. Thiobacillus cinsinin əksər təcrid olunmuş mikroorqanizmlərinin inkişafı üçün optimal temperatur təxminən 30 ° C-dir. Lakin təbii şəraitdə onlar 55°C-ə qədər olan temperaturda termal bulaqlarda rast gəlinir. Bu yaxınlarda Thiobacillus sp.-nin təmiz kulturası təcrid edilmişdir. 50 ° C-də böyümək üçün optimaldır.
Ətraf mühitin turşuluğuna gəldikdə, tiobacilli neytral və ya hətta qələvi reaksiyada yaxşı inkişaf edən orqanizmlərə və asidofil olan növlərə bölünür, yəni. asidik mühitdə inkişaf edir və çox aşağı pH dəyərlərinə tab gətirir.
Birinci qrupa T. thioparus, T. denitrificans, T. novellus, T. thiocyanoxidans, T. neapolitanus kimi növlər daxildir. Bu mikroorqanizmlər üçün optimal pH dəyəri 6.0-9.0 bölgəsindədir və onların böyüməsinin mümkün olduğu pH dəyərləri zonası 3.0-6.0 ilə 10.0-11.0, müxtəlif növlər və suşlar üçün optimal pH dəyərləri və. onların böyüməsinin müşahidə olunduğu aktiv turşuluq bölgəsi əhəmiyyətli dərəcədə fərqlənə bilər.
İkinci qrupa T. thiooxidans, T. ferroxidans, T. intermedius daxildir. Bu mikroorqanizmlər üçün optimal pH dəyəri 2,0-4,0-dır və böyümə 0,5-2,0 ilə 5,0-7,0 arasında olan pH-da mümkündür. İlk iki növ ən asidofilik orqanizmlərdir. Bu bakteriyaların hər ikisi 5.0-dən çox olmayan pH dəyərlərində böyüyür. Eyni zamanda göstərilmişdir ki, T. thiooxidans 0-a yaxın pH dəyərində həyat qabiliyyətini saxlayır, bu da 1,0 n-ə uyğundur. sulfat turşusu məhlulu. Bu, bəlkə də tədqiqatçılara məlum olan ən asidofil mikroorqanizmdir.
Tionik bakteriyaların əksəriyyəti yalnız oksigenin iştirakı ilə inkişaf edir, baxmayaraq ki, bəzi nümayəndələrin böyüməsi aşağı oksigen səviyyəsində mümkündür. Lakin fakultativ anaeroblar da məlumdur. Bunlara T. denitrificans daxildir. Aerob şəraitdə bu bakteriyalar molekulyar oksigenin iştirakı ilə oksidləşmə proseslərini həyata keçirir, anaerob şəraitdə isə denitrifikasiyaya keçir və nitratları molekulyar azota qədər azaldır. Maraqlıdır ki, T. denitrificans, Paracoccus denitrificans kimi, nitratları azot mənbəyi kimi mənimsəyə bilmir və böyümə üçün azot mənbəyi kimi ammoniuma ehtiyac duyur.
Tion bakteriyaları hidrogen sulfid, sulfidlər, sulfit, tiosulfat, tetrationat, tiosiyanat (rodanid), ditionit kimi kükürd birləşmələrini, həmçinin molekulyar kükürdləri oksidləşdirərək tam oksidləşmə zamanı sulfatlar əmələ gətirir. Ancaq fərdi növlərin qabiliyyətləri tamamilə eyni deyil. Bundan əlavə, hansı kükürd birləşmələrinin bioloji oksidləşdiyini müəyyən etmək həmişə asan deyil, çünki onların çoxu aşağı pH-da qeyri-sabitdir və atmosfer oksigeni ilə də oksidləşə bilər.
Neytral və qələvi mühitdə böyüyən bütün növlər, bir qayda olaraq, hidrogen sulfid, kükürd və tiosulfat oksidləşdirir. T. thiocyanoxidans üçün xarakterik xüsusiyyət tiosiyanidi oksidləşdirmək qabiliyyətidir. Bu əsasda, ayrı bir növ kimi fərqləndirilir, çünki digər xüsusiyyətlərinə görə bu mikroorqanizm T. thioparusa bənzəyir. T. thiooxidans kimi asidofil bakteriya adətən tərkibində molekulyar kükürd olan mühitdə becərilir. Bu növün hidrogen sulfid və digər kükürd birləşmələrini oksidləşdirmək qabiliyyəti məsələsi tam həll edilməmişdir, çünki bu birləşmələr asidik şəraitdə qeyri-sabitdir. T. ferroksidanlara gəldikdə, bu bakteriyaların həm molekulyar kükürdü, həm də onun müxtəlif birləşmələrini oksidləşdirə biləcəyinə dair sübutlar var, yəni: hidrogen sulfid, tiosulfat, ditionit, tetrasulfat, sulfit. Bundan əlavə, T. ferroksidans suda həll olunmayan ağır metal sulfidlərinin oksidləşməsində fəal iştirak edir. Bunlara pirit (FeS2), xalkopirit (CuFeS2), stibnit (SbS2), xalkosit (Cu2S), kavellit (CuS), pirrotit (FeS), realqar (AsS), violarit (Ni2FeS4) və s. kimi minerallar daxildir. Qalanları. bakteriyalarda ya bu qabiliyyət yoxdur, ya da zəif ifadə olunur.
T. ferroksidanların fərqli xüsusiyyəti həm də dəmir dəmirini oksid dəmirə oksidləşdirmək qabiliyyətidir. Bu əsasda bu bakteriyalar Thiobacillus cinsinə aid olsalar da, dəmir bakteriyaları hesab edilə bilər.
Yuxarıda qeyd edildiyi kimi, molekulyar kükürdün və onun müxtəlif birləşmələrinin tion bakteriyaları tərəfindən oksidləşməsinin son məhsulu sulfatdır. Əgər proses bu şəkildə gedirsə, yəni. orijinal substratın tam oksidləşməsi baş verir, sonra onun nəticələri aşağıdakı tənliklərdə əks olunur. Hidrogen sulfidin oksidləşməsi zamanı:
Aerob şəraitdə kükürdün və tiosulfatın oksidləşməsi zamanı:

Nitratların istifadəsi ilə anaerob şəraitdə T. denitrificans tərəfindən kükürdün və tiosulfatın oksidləşməsi zamanı:

T. thiocyanoxidans tiosiyanidin oksidləşməsi zamanı:

Bununla belə, oksidləşmə çox vaxt sona çatmır və mühitdə müxtəlif natamam oksidləşmiş məhsullar tapılır. Beləliklə, hidrogen sulfidin oksidləşməsi zamanı bəzən molekulyar kükürd meydana çıxır, tiosulfat və politionatlar da tapılır. Molekulyar kükürdün bakteriyalar tərəfindən oksidləşməsi zamanı tiosulfat və politiyonatların görünüşü müşahidə edilmişdir. Tiosulfat oksidləşməsi də tez-tez politiyonatların (tritionat, tetrationat, peitatiopatat) və elementar kükürdün əmələ gəlməsi ilə müşayiət olunur. Sülfit də tam oksidləşməmiş məhsullardan biridir. Lakin bu birləşmələrin hamısı fermentativ proseslərin nəticəsi ola bilməz və əslində orijinal substratın bakterial oksidləşməsinin aralıq məhsullarına aiddir. Onların bəziləri sırf kimyəvi və ya yan bioloji reaksiyalar nəticəsində əmələ gəlir. Buna görə də, müxtəlif kükürd birləşmələrinin mikroorqanizmlər tərəfindən oksidləşmə yollarını aydınlaşdırmaq çox çətindir və bütün reaksiyalar hələ də aydınlaşdırılmamışdır.
Sulfid çevrilməsinin birinci mərhələsinin təbiəti haqqında hələ də çox az məlumat var və kükürdün onun oksidləşməsinin birbaşa və ya əlavə məhsulu olması tam aydın deyil.
Bakteriyalar tərəfindən elementar kükürdün istifadəsi məsələsi çox mürəkkəbdir və onun iki aspekti var: mikroorqanizmlərin bu suda həll olunmayan maddə üzərində necə hərəkət etməsi və oksidləşmə prosesinin özü necə baş verir.
Tiobacillusun kükürd üzərində təsir mexanizmi ilə bağlı iki fikir var.
1. Kükürdün bakteriyalar tərəfindən oksidləşməsi üçün hüceyrələrlə birbaşa təmasda olmalıdır.
2. Kükürd bakteriyalar tərəfindən ətraf mühitə buraxdıqları lipid xarakterli maddələrdə ilkin həll edildikdən sonra istifadə olunur.
T. thiooxidans molekulyar kükürdlü mühitdə böyüdükdə, mühitdə fosfolipidlər tapılır. Bəzi məlumatlara görə, bu, fosfatidilinositol, digərlərinə görə, bir qədər fərqli birləşmələrdir (fosfatidil-N-metiletanol, fosfatidilqliserin, difosfatidilqliserin) və onların yığılması bitkilərin aktiv böyüməsi mərhələsinə təsadüf edir. Lakin ikinci fərziyyəni sübuta yetirilmiş hesab etmək olmaz. Çox güman ki, kükürdün bakteriyalar tərəfindən oksidləşməsi üçün həm onun hüceyrələrlə təması, həm də kükürdün “yaşlanması” üçün müəyyən maddələrin buraxılması vacibdir.
Kükürdün oksidləşmə yoluna gəldikdə, məlumatlar da olduqca ziddiyyətlidir. Aşağıdakı sxem hidrogen sulfidin oksidləşməsi üçün də məqbul olan ən çox ehtimal olunan sxem hesab olunur:

Ehtimal olunur ki, X ya hidrogen sulfid və ya kükürdün reaksiyaya girdiyi qlutatyon törəməsidir, ya da hüceyrə membranlarına bağlanmış tioldur. Tiosulfatın əmələ gəlməsi bakteriyaların iştirakı olmadan baş verə bilən qeyri-enzimatik bir proses hesab olunur.
Tiosulfat oksidləşməsinin öyrənilməsinə çoxlu sayda tədqiqatlar həsr edilmişdir. Bütün hüceyrələr və müxtəlif növ tiobaksillərin hüceyrəsiz preparatları ilə aparılan təcrübələrdən əldə edilən məlumatlar göstərir ki, tiosulfat çevrilməsinin ilkin mərhələsi ya onun reduktaza təsiri altında hidrogen sulfidin və sulfitin əmələ gəlməsi ilə azalması ilə əlaqələndirilə bilər:

Və ya elementar kükürd və sulfitə parçalanma ilə:

və ya nəhayət, tetrationata oksidləşmə və sonra tritionata və sulfitə çevrilmə ilə:

Bununla belə, güman edilir ki, tiosulfatın çevrilməsi üçün sonuncu yol əsas deyil. Tiosulfatın sulfit əmələ gətirmək üçün parçalanmasına gəldikdə, belə bir reaksiya kükürdü tiosiyanat şəklində bağlayan rodonaz tərəfindən həyata keçirilə bilər:
Yalnız tion və digər bakteriyalarda deyil, heyvanlarda da geniş yayılmışdır. Bununla belə, bu fermentin tionik bakteriyalar tərəfindən tiosulfat mübadiləsində iştirak edib-etməməsi qəti şəkildə müəyyən edilməmişdir.
Sülfit oksidləşmə yolu daha çox tədqiq edilmişdir. Pek laboratoriyasında T. thioparus ilə aparılan işlər göstərdi ki, sulfit adenozin monofosfatla (AMF) reaksiya verir və nəticədə adenil sulfat və ya adenozin fosfosülfat (APS) adlı birləşmə əmələ gəlir. Növbəti mərhələdə API və qeyri-üzvi fosfat (Pn) arasında adenozin difosfatın (ADP) əmələ gəlməsi və sərbəst sulfatın sərbəst buraxılması ilə reaksiya baş verir:

Adenilat kinaz fermentinin təsiri nəticəsində iki ADP molekulu ATP-yə çevrilə və yenidən AMP istehsal edə bilər:
Beləliklə, sulfit oksidləşməsinin bu yolu sözdə substratın fosforlaşması nəticəsində enerjinin (ATP) istehsalı ilə əlaqələndirilir. Eyni zamanda, sulfit oksidləşmə prosesi elektronların tənəffüs zəncirinə köçürülməsi ilə müşayiət oluna bilər, onun fəaliyyəti ATP sintezi ilə bağlıdır.
Bir sıra tion bakteriyaları, o cümlədən T. thioparus üçün elektronların tənəffüs zəncirinə keçməsini təmin edən sulfit oksidləşdirici fermentin təsiri nəticəsində sulfit oksidləşməsinin APS əmələ gəlmədən mümkün olduğu göstərilmişdir. sitoxromlar səviyyəsində:
![]()
Ola bilsin ki, eyni orqanizmdə sulfit və digər kükürd birləşmələrinin oksidləşməsi üçün müxtəlif yollar ola bilər və bu və ya digərinin əhəmiyyəti ətraf mühit şəraitindən və digər amillərdən asılıdır. Hazırda mövcud olan məlumatlara əsasən, müxtəlif kükürd birləşmələrinin tion bakteriyaları tərəfindən oksidləşməsi aşağıdakı ümumiləşdirilmiş sxemlə göstərilə bilər.

Tionik bakteriyaların elektron daşıma sistemlərinin komponentlərinə gəldikdə, bütün məlumatlara görə, müxtəlif növlərdə onların tərkibinə həmişə c tipli sitoxromlar daxildir. B tipli sitoxromlar və yəqin ki, onun sitoxrom o adlanan variantı, bəzi nümayəndələrdə isə a və d sitoxromları da aşkar edilmişdir. Bundan əlavə, elektron nəqliyyat sisteminə flavoproteinlər və ubiquinonlar da daxildir. Lakin nitrifikasiya edən bakteriyalar kimi elektronların oksigenə ötürülməsini təmin edən tionik bakteriyaların tənəffüs zənciri nisbətən qısadır, çünki oksidləşmiş substratlar kifayət qədər yüksək redoks potensialına malikdir. Buna görə də, bu mikroorqanizmlər üçün termodinamik qradientə qarşı enerjidən asılı olan elektron köçürməsi (əks elektron ötürülməsi) vacibdir. Karbon qazının və digər konstruktiv proseslərin assimilyasiyası üçün lazım olan azaldılmış NAD-ın əmələ gəlməsini təmin edir. Onu da qeyd etmək lazımdır ki, elektron daşıma sistemlərinin komponentləri və elektron ötürmə yolları yalnız tiop bakteriyalarının müxtəlif nümayəndələrində fərqli fərqlərə malik ola bilməz, həm də oksidləşmiş substratın təbiətindən asılı ola bilər. Buna görə də ümumiləşdirilmiş diaqram (şək. 140) çox şərtlidir.

Müxtəlif müəlliflərin hesablamalarına görə, tionik bakteriyaların sərbəst enerjidən istifadəsinin səmərəliliyi 2 ilə 37% arasında dəyişir. Digər kemoavtotroflar kimi tionik bakteriyalar da karbon qazını ilk növbədə Kalvin dövrü vasitəsilə mənimsəyir. Lakin onlar müəyyən metabolitlərin əmələ gəlməsi üçün vacib olan digər karboksilləşmə reaksiyalarını da həyata keçirmək qabiliyyətinə malikdirlər.
Həddindən artıq termoasidofilik kükürddən asılı olan arxelər elementar kükürdün iştirakı ilə də litotrofik böyüməyə qadirdir və 40-100 °C və pH 1-6 səviyyəsində inkişaf edə bilər; bunlar hidrotermlərin və vulkanik kalderaların sakinləridir.
Heterotrof kükürd oksidləşdirici mikroorqanizmlər (qidalanma üçün üzvi maddələrdən istifadə edən orqanizmlər).
Bəzi məcburi heterotrof bakteriyaların, mayaların və mikromisetlərin müxtəlif qeyri-üzvi kükürd birləşmələrini oksidləşdirmək qabiliyyəti kifayət qədər uzun müddətdir məlumdur, baxmayaraq ki, bu reaksiyaların heterotrofların metabolizmindəki əhəmiyyəti hələ də tam başa düşülmür. Heterotrofik kükürd oksidləşdirici bakteriyalar arasında tiosulfatın tetrationata qədər natamam oksidləşməsini həyata keçirən psevdomonad qrupunun növləri üstünlük təşkil edir.
Həmçinin S.N. Winogradsky (1887-1889) qeyd etdi ki, rəngsiz kükürd bakteriyaları çox az miqdarda üzvi maddələr olan suda inkişaf edə bilər və buna görə də karbon qazını udmaq qabiliyyətinə sahib olduqlarını güman edirdi. Beggiatoa-nın müxtəlif suşlarının fiziologiyasının tədqiqinə əsaslanaraq, Pringsheim hesab edir ki, onların arasında hidrogen sulfidini oksidləşdirən və CO 2-ni fiksasiya edən avtotroflar və üzvi birləşmələr tələb edən nümayəndələr var. Lakin bir sıra heterotrof Beggiatoa ştammları da üzvi birləşmələrin iştirakı ilə hidrogen sulfidini oksidləşdirir, ehtimal ki, enerji istehsal edir, yəni. onlar kimolitoheterotroflardır. Rəngsiz kükürd bakteriyaları ilə yanaşı, hidrogen sulfid, molekulyar kükürd və tiosulfat oksidləşməsində iştirak edən tipik heterotrof mikroorqanizmlər məlumdur. Bunlara Bacillus, Pseudomonas, Achromobacter, Sphaerotilus, həmçinin aktinomisetlər, qəliblər (Penicillium luteum, Aspergillus niger), maya və Alternaria nümayəndələri daxildir. Onlardan bəziləri, xüsusən də saplı çoxhüceyrəli bakteriya Sphaerotilus natans, hidrogen sulfidin iştirakı ilə hüceyrələrdə kükürd yatırır. Digərləri (Pseudomonas aeruginosa, Ps. fluorescens, Achromobacter stuzeri) tiosulfatı tetrationata (Na2S4O6) oksidləşdirməyə qadirdir. Heterotrof mikroorqanizmlərin qarışıq mədəniyyətləri elementar kükürdlə reaksiya verdikdə politiyonatların və sulfatların əmələ gəlməsi də qeyd edilmişdir. Oksidləşmə mexanizmi və bu prosesin heterotroflar üçün bioloji əhəmiyyəti qeyri-müəyyən olaraq qalır. Bəzi heterotrof rəngsiz kükürd bakteriyaları üzvi maddələrin oksidləşməsi zamanı hüceyrələrdə əmələ gələn superoksid radikalı olan hidrogen peroksid ilə kükürd birləşmələrini oksidləşdirməyə qadirdir: H 2 O 2 + H 2 S → S 0 + 2H 2 O. Bu prosesin fizioloji mənası. oksigenin natamam azaldılmasının zəhərli məhsullarının detoksifikasiyasıdır ( H 2 O 2, O 2-).
Kükürd birləşmələrinin sulfata qədər tam oksidləşməsi elementar kükürdlə zənginləşdirilmiş torpaqlarda yaşayan mikromisetlər üçün daha xarakterikdir. Aspergillus, Penicillium, Trichoderma, Fusarium, Mucor və Auerobasidium mikroskopik göbələklərinin bəzi növləri üçün elementar kükürd, tiosulfat və hətta metal sulfidləri sulfata oksidləşdirmək qabiliyyəti göstərilmişdir, lakin belə oksidləşmə sürəti bir və ya iki sifarişdir. litotrof bakteriyalardan daha az böyüklükdə.
kükürdün siklik çevrilməsi mikroorqanizmi
Nəticə
Son onilliklərdə təbii kükürd dövranı artan antropogen təsirə məruz qalmış, bu da zəhərli kükürd birləşmələrinin toplanmasına və təbii kükürd dövranında balansın pozulmasına səbəb olmuşdur. Xüsusilə, kükürd birləşmələrinin böyük miqyaslı emissiyaları mədən yanacaqlarının, selüloz-kağız və metallurgiya müəssisələri tərəfindən ayrılan hidrogen sulfid və uçucu üzvi sulfidlərin yandırılması zamanı, habelə bələdiyyələrin parçalanması zamanı istilik elektrik stansiyalarının buraxdığı kükürd dioksidin əmələ gəlməsi ilə nəticələnir. və kənd təsərrüfatı tullantı suları. Bu birləşmələr mikroqram konsentrasiyalarında belə zəhərlidir. Onlar havanı zəhərləyə, atmosfer kimyasına təsir edə və suda həll olunan oksigen çatışmazlığına səbəb ola bilər.
Buna görə də kükürd birləşmələrinin çevrilməsində və zəhərli kükürd birləşmələrinin detoksifikasiyasında iştirak edən prokariotların istifadəsi çox aktual görünür. Bundan əlavə, mikroorqanizmlərin son dərəcə yüksək katalitik aktivliyi onların Yer səthində baş verən kimyəvi çevrilmələrdə böyük rol oynamasına səbəb olur. Kiçik ölçülərinə görə mikroorqanizmlər heyvanlar və ali bitkilərlə müqayisədə yüksək səth-həcm nisbətinə malikdirlər ki, bu da hüceyrə ilə ətraf mühit arasında substratların və ifrazat məhsullarının sürətli mübadiləsinə səbəb olur.
Əhəmiyyətli amillər həm də əlverişli şəraitdə onların çoxalma sürətinin yüksək olması və bütün biosferdə geniş yayılmasıdır. Bununla belə, qida maddələrinin çevrilməsində prokariotların müstəsna əhəmiyyətinə baxmayaraq, maddələrin dövrəsində və xüsusən də kükürd dövrəsində prokariotik fəaliyyətin miqyası hələ tam qiymətləndirilməmişdir.
Ədəbiyyat
1. Qraboviç M.Yu. Prokaryotların kükürd dövranında iştirakı // Soros Educational Journal, 1999, No 12, s. 16-20.
2. Qromov B.V., Pavlenko G.V. Bakteriyaların ekologiyası. L.: Leninqrad Dövlət Universitetinin nəşriyyatı, 1989. 248 s.
3. Qromov B.V. Arxeyanın heyrətamiz dünyası // Soros Təhsil Jurnalı. 1997. № 4. səh. 23–26.
4. Kaldera mikroorqanizmləri / Ed. G.A. Zavarzina. M.: Nauka, 1989. 120 s.
5. Kondratyeva E.N. Xemolitotroflar və metilotroflar. M.: Moskva Dövlət Universitetinin nəşriyyatı, 1983. 172 s.
6. Malaxov V.V. Vestimentifera avtotrof heyvanlardır // Soros Təhsil Jurnalı. 1997. № 9. səh. 18–26.
Planetin biosferi vahid meqaorqanizmdir, hissələri bir-biri ilə ahəngdar şəkildə bağlıdır. Həyatın bütün müxtəlifliyi iki super krallığa bölünür - prokaryotlar (nüvədən əvvəl orqanizmlər) və eukaryotlar (nüvəsi olan). Həddindən artıq krallıqlar canlı krallıqlara bölünür:
- viruslar;
- bakteriya;
- göbələk;
- bitkilər;
- heyvanlar.
Bakteriyalar səltənəti viruslar səltənəti ilə birlikdə prokariotların fövqəl krallığına - nüvəsiz orqanizmlərə birləşir. Tarixən bu, planetdəki canlı orqanizmlərin ilk eşelonudur.
Canlı təkhüceyrəli orqanizmlər təxminən 3,8 milyard il əvvəl meydana çıxdı. Təxminən bir milyard il ərzində onlar planetin yeganə canlı sakinləri idilər - uğurla çoxaldılar, inkişaf etdilər və uyğunlaşdılar. Onların həyati fəaliyyətinin nəticəsi planetin atmosferində çoxhüceyrəli orqanizmlərin - göbələklərin, bitkilərin, çiçəklərin və heyvanların yaranmasına imkan verən sərbəst oksigenin meydana çıxması idi.
Hal-hazırda canlı bakteriyalar hər yerdə yaşayır: nadir atmosfer təbəqələrindən tutmuş ən dərin okean çökəkliklərinə qədər, Arktika buzlarında və termal geyzerlərdə yaşayırlar. Bakteriyalar təkcə boş məkanı deyil, həm də göbələklər, bitkilər və ya heyvanlar olsun, digər orqanizmlərin daxilində inkişaf edirlər.

Təbiətdə bütün heyvanlar və insanlar istisna deyil, mikroblar üçün yaşayış yeridir:
- dəri;
- ağız boşluğu;
- bağırsaqlar.
Tədqiqatçılar müəyyən ediblər ki, insan orqanizmində yaşayan mikroorqanizm hüceyrələrinin sayı onun öz hüceyrələrinin sayından 10 dəfə çoxdur. Belə yüksək kəmiyyət göstəricilərinə baxmayaraq, bədəndə yaşayan bakteriyaların çəkisi 2 kq-dan çox deyil - bu, hüceyrə ölçüsündə əhəmiyyətli bir fərqlə bağlıdır.
Bakteriyalar səltənətinin canlı nümayəndələrinin saysız-hesabsız növləri var, lakin onların hamısının ortaq cəhətləri bunlardır:
- aydın bir nüvənin olmaması;
- çox kiçik (bitki və heyvan hüceyrələri ilə müqayisədə) hüceyrə ölçüləri;
- Bioloji vahid hüceyrənin özüdür, onlar birləşdirildikdə, bir bakteriya koloniyasından danışırıq.
Məhz bakteriya krallığının nümayəndələri göbələklərin, bitkilərin və heyvanların meydana gəlməsini mümkün etdi. Planetdə peyda olan mikroorqanizmlər nəinki mövcud şəraitə uyğunlaşdılar - onlar yaşayış yerlərini aktiv şəkildə dəyişdirərək keyfiyyətcə yeni xüsusiyyətlər yaratdılar.

Təbiətdə azot və karbonun dövranı yalnız mikroorqanizmlər sayəsində baş verir. Alimlər müəyyən ediblər ki, mikroblar biosferdən çıxarılsa, planetdə həyat davam edə bilməz.
Biosfer dövrlərində prokariotların rolu
Planetdə həyatın yaranmasında bakteriya krallığının nümayəndələri biosferin formalaşmasında fəal iştirak edirdilər. Müasir biosferin fəaliyyət səviyyəsini saxlamaq üçün mikroorqanizmlərə ehtiyacı var - təbiətdəki enerji və maddənin dövranı mikroblar tərəfindən təmin edilir.
Canlı mikrobların biosfer proseslərində dominant roluna misal olaraq münbit torpaq qatının yaradılması və saxlanmasını göstərmək olar.
Qaz və oksidləşmə funksiyaları ilə yanaşı, mikroorqanizmlərin geokimyəvi funksiyaları getdikcə daha çox əhəmiyyət kəsb edir. Fermentlərin aktivliyi və konsentrasiya funksiyaları planetin geokimyasına əhəmiyyətli təsir göstərir.
Mikroorqanizmlərin növ müxtəlifliyi
Bakteriyalar krallığının nümayəndələri biosferin bütün səviyyələrində məskunlaşmışlar və mikrobların olması ilə planetin biosferinin yuxarı və aşağı sərhədlərini müəyyən edirlər. Belə müxtəlif fiziki parametrlərə malik mühitlərdə yaşayan mikroblar bir sıra xüsusiyyətlərə görə fərqlənirlər.
- Canlı bakteriya hüceyrəsinin formasına görə:
- sferik kokklar;
- çubuq şəklində;
- qıvrımlı, vibrionlara və spiroketlərə bölünür.
- Bədənin kosmosda hərəkət tərzinə görə:
- flagella olmadan (Browniana bənzər xaotik hərəkət);
- flagella istifadə edərək (sayı bütün perimetr boyunca birdən çoxa qədər dəyişir).
- Bakteriyalar krallığının nümayəndələrinin metabolik xüsusiyyətlərinə görə:
- qeyri-üzvi maddələrdən - avtotroflardan zəruri maddələrin sintezi;
- üzvi maddələrin emalı - heterotroflar.
- Enerji əldə etmə üsuluna görə:
- tənəffüs (aerob və anaerob mikroorqanizmlər);
- fermentasiya;
- fotosintez (oksigensiz və oksigensiz).

Mikroblar və viruslar arasındakı əlaqənin xüsusiyyətləri - eyni adlı krallıqların nümayəndələri
Prokariotların super krallığı iki krallığı birləşdirir - bakteriya və viruslar, oxşarlıqlardan daha çox fərqlər var. Məsələn, bakteriyalar həyatı təmin etmək üçün lazım olan bütün maddələri sintez edirsə, viruslar ümumiyyətlə zülal sintez etməyə qadir deyillər. Onlar hətta öz növbəsində çoxalda bilməzlər, ancaq başqasının hüceyrəsinə sızaraq.
Viruslar ev sahibi hüceyrənin DNT-sini bloklayır və onu özlərininki ilə əvəz edir - nəticədə tutulan hüceyrə işğalçı virusun nüsxələrini əmələ gətirir ki, bu da adətən onun ölümünə səbəb olur.