Funcțiile membranei plasmatice. Membrana plasmatica Structura si functia plasmei
Membrana plasmatică ocupă o poziție specială, deoarece limitează celula în exterior și este direct conectată cu mediul extracelular. Are o grosime de aproximativ 10 nm și este cea mai groasă membrană celulară. Componentele principale sunt proteinele (peste 60%), lipidele (aproximativ 40%) și carbohidrații (aproximativ 1%). Ca toate celelalte membrane celulare, este sintetizat în canalele EPS.
Funcțiile plasmalemei.
Transport.
Membrana plasmatică este semi-permeabilă, adică. prin ea trec selectiv diferite molecule cu viteze diferite. Există două moduri de a transporta substanțele prin membrană: transport pasiv și activ.
Transport pasiv. Transportul pasiv sau difuzia nu necesită aport de energie. Moleculele neîncărcate difuzează de-a lungul gradientului de concentrație, transportul moleculelor încărcate depinde de gradientul de concentrație al protonilor de hidrogen și de diferența de potențial transmembranar, care sunt combinate într-un gradient de protoni electrochimic. De regulă, suprafața citoplasmatică interioară a membranei poartă o sarcină negativă, care facilitează pătrunderea ionilor încărcați pozitiv în celulă. Există două tipuri de difuzie: simplă și ușoară.
Difuzia simplă este tipică pentru molecule neutre mici (H 2 O, CO 2, O 2), precum și pentru substanțele organice hidrofobe cu greutate moleculară mică. Aceste molecule pot trece prin porii sau canalele membranei fără nicio interacțiune cu proteinele membranei atâta timp cât se menține gradientul de concentrație.
Difuzia facilitată este caracteristică moleculelor hidrofile, care sunt transportate de-a lungul membranei și de-a lungul gradientului de concentrație, dar cu ajutorul unor proteine transportoare membranare speciale conform principiului uniformele.
Difuzia facilitată este foarte selectivă, deoarece proteina purtătoare are un centru de legare complementar substanței transportate, iar transferul este însoțit de modificări conformaționale ale proteinei. Unul dintre mecanismele posibile de difuzie facilitată este următorul: proteina de transport (translocaza) leagă substanța, apoi se apropie de partea opusă a membranei, eliberează această substanță, își asumă conformația inițială și este din nou gata să îndeplinească funcția de transport. Se știe puțin despre cum se mișcă proteina în sine. Un alt posibil mecanism de transfer implică implicarea mai multor proteine purtătoare. În acest caz, compusul legat inițial în sine trece de la o proteină la alta, legându-se secvenţial cu una sau alta proteină până când se află pe partea opusă a membranei.
Transport activ. Un astfel de transport are loc atunci când transferul este efectuat în raport cu gradientul de concentrație. Necesită cheltuirea de energie de către celulă. Transportul activ servește la acumularea de substanțe în interiorul celulei. Sursa de energie este adesea ATP. Pentru transportul activ, pe lângă sursa de energie, este necesară participarea proteinelor membranare. Unul dintre sistemele de transport active din celulele animale este responsabil pentru transportul ionilor de Na și K + prin membrana celulară. Acest sistem este numit N / A + - K * -pompa. Este responsabil pentru menținerea compoziției mediului intracelular, în care concentrația ionilor K + este mai mare decât cea a ionilor Na *.
Gradientul de concentrație al ambilor ioni este menținut prin transferul K + în interiorul celulei și Na + în exterior. Ambele transporturi au loc pe un gradient de concentrație. Această distribuție a ionilor determină conținutul de apă din celule, excitabilitatea celulelor nervoase și a celulelor musculare și alte proprietăți ale celulelor normale. Na + -K + -pompa este o proteină - transport ATPaza. Molecula acestei enzime este un oligomer și pătrunde în membrană. În timpul ciclului complet al funcționării pompei, 3 ioni Na + sunt transferați de la celulă la substanța intercelulară, iar în direcția opusă - 2 ioni K +, în timp ce energia moleculei ATP este utilizată. Există sisteme de transport pentru transferul ionilor de calciu (Ca 2+ -ATPaza), pompe de protoni (H + -ATPaza), etc.
Se numește transferul activ al unei substanțe prin membrană, realizat datorită energiei gradientului de concentrație al unei alte substanțe simptom... În acest caz, ATPaza de transport are locuri de legare pentru ambele substanțe. Antiport este mișcarea unei substanțe față de gradientul său de concentrație. În acest caz, o altă substanță se mișcă în direcția opusă de-a lungul gradientului său de concentrație. Simptomul și antiportul (cotransportul) pot apărea în timpul absorbției aminoacizilor din intestin și reabsorbției glucozei din urina primară, folosind energia gradientului de concentrație al ionilor de Na + creat de Na +, K + -ATPaza.
Alte 2 tipuri de transport sunt endocitoza si exocitoza.
Endocitoza- captarea particulelor mari de către celulă. Există mai multe moduri de endocitoză: pinocitoză și fagocitoză. De obicei sub pinocitozaînțelege captarea particulelor coloidale lichide de către o celulă, sub fagocitoză- captarea corpusculilor (particule mai dense si mai mari pana la alte celule). Mecanismul de pino și fagocitoză este diferit.
În general, intrarea în celulă a particulelor solide sau a picăturilor lichide din exterior se numește heterofagie. Acest proces este cel mai răspândit la protozoare, dar este foarte important la oameni (precum și la alte mamifere). Heterofagia joacă un rol esențial în apărarea organismului (neutrofile segmentate - granulocite; macrofagocite), remodelarea țesutului osos (osteoclaste), formarea tiroxinei de către foliculii tiroidieni, reabsorbția proteinelor și a altor macromolecule în nefronul proximal și alte procese.
Pinocitoza.
Pentru ca moleculele externe să intre în celulă, acestea trebuie mai întâi legate de receptorii glicocalix (un set de molecule legate de proteinele de suprafață ale membranei) (Fig.).
La locul unei astfel de legături, moleculele de proteină clatrină se găsesc sub plasmalemă. Plasmalema, împreună cu moleculele atașate din exterior și clatrina aflată la baza citoplasmei, începe să invadeze. Proeminența devine mai adâncă, marginile ei se apropie și apoi se închid. Ca rezultat, o bulă este desprinsă de plasmalema, care transportă moleculele capturate. Clathrin pe suprafața sa arată ca o margine neuniformă pe micrografiile electronice, prin urmare astfel de bule se numesc mărginite.
Clathrin împiedică veziculele să adere la membranele intracelulare. Prin urmare, veziculele mărginite pot fi transportate liber în celulă exact în acele părți ale citoplasmei în care conținutul lor ar trebui utilizat. Acesta este modul în care, în special, hormonii steroizi sunt eliberați în nucleu. Cu toate acestea, de obicei veziculele mărginite își dispar granița la scurt timp după scindarea din plasmalemă. Clatrina este transferată în plasmalemă și poate participa din nou la reacțiile de endocitoză.
La suprafața celulei din citoplasmă există mai multe vezicule permanente - endozomi. Veziculele mărginite eliberează clatrina și se contopesc cu endozomii, crescând volumul și suprafața endozomilor. Apoi, partea în exces a endozomilor este despărțită sub forma unei noi vezicule, în care nu există substanțe care au intrat în celulă, acestea rămânând în endozom. Noua veziculă este direcționată către suprafața celulei și fuzionează cu membrana. Ca urmare, scăderea plasmalemei, care are loc în timpul divizării veziculei mărginite, este restabilită, în timp ce receptorii săi revin și ei la plasmalemă.
Endozomii sunt scufundați în citoplasmă și fuzionați cu membranele lizozomului. Substanțele care intră în interiorul unui astfel de lizozom secundar suferă diferite transformări biochimice. La finalizarea procesului, membrana lizozomului se poate dezintegra în fragmente, iar produsele de degradare și conținutul lizozomului devin disponibile pentru reacții metabolice intracelulare. De exemplu, aminoacizii se leagă de ARNt și sunt eliberați în ribozomi, în timp ce glucoza poate intra în complexul Golgi sau în tubii EPS agranulare.
Deși endozomii nu au o margine de clatrină, nu toți fuzionează cu lizozomi. Unele dintre ele sunt direcționate de la o suprafață celulară la alta (dacă celulele formează un strat epitelial). Acolo, membrana endozomului se contopește cu plasmolema și conținutul este excretat. Ca urmare, substanțele sunt transferate prin celulă dintr-un mediu în altul fără modificări. Acest proces se numește transcitoza... Moleculele de proteine, în special imunoglobulinele, pot fi de asemenea transportate prin transcitoză.
Fagocitoză.
Dacă o particulă mare are pe suprafața sa grupări moleculare care pot fi recunoscute de receptorii celulari, se leagă. În niciun caz, particulele străine nu posedă întotdeauna astfel de grupuri. Cu toate acestea, ajungând în organism, ele sunt înconjurate de molecule de imunoglobuline (opsonine), care se găsesc întotdeauna în sânge și în mediul extracelular. Imunoglobulinele sunt întotdeauna recunoscute de celulele fagocitare.
După ce opsoninele care acoperă particulele străine se leagă de receptorii fagocitei, complexul său de suprafață este activat. Microfilamentele de actină încep să interacționeze cu miozina și configurația suprafeței celulare se modifică. Excrescențe ale citoplasmei fagocitelor sunt întinse în jurul particulei. Acestea acoperă suprafața particulei și se combină peste ea. Frunzele exterioare ale excrescentelor se îmbină, închizând suprafața celulei.
Plăcile adânci de excrescențe formează o membrană în jurul particulei absorbite - se formează fagozom. Fagozomul se contopește cu lizozomii, rezultând complexul lor - heterolizozom (heterozom, sau fagolizozom).În el, are loc liza componentelor capturate ale particulei. Unii dintre produșii de liză sunt îndepărtați din heterozom și utilizați de celulă, în timp ce unii nu pot fi susceptibili de acțiunea enzimelor lizozomale. Aceste reziduuri formează corpuri reziduale.
Potenţial toate celulele au capacitatea de a fagocitoză, dar în organism, doar câteva sunt specializate în această direcţie. Acestea sunt leucocite neutrofile și macrofage.
exocitoză.
Aceasta este eliminarea substanțelor din celulă. În primul rând, compușii cu molecule mari se segregă în complexul Golgi sub formă de bule de transport. Acestea din urmă, cu participarea microtubulilor, sunt direcționate către suprafața celulei. Membrana veziculei este înglobată în plasmalemă, iar conținutul veziculei se află în afara celulei (Fig.) Fuziunea veziculei cu plasmalema poate avea loc fără semnale suplimentare. Această exocitoză se numește constitutiv. Acesta este modul în care majoritatea produselor propriului metabolism sunt îndepărtate din celulă. O serie de celule, însă, sunt concepute pentru a sintetiza compuși speciali - secrete care sunt folosite în alte părți ale corpului. Pentru ca vezicula de transport cu secretul să se îmbine cu membrana plasmatică, sunt necesare semnale externe. Abia atunci va avea loc fuziunea și secretul va fi dezvăluit. Această exocitoză se numește reglementate... Se numesc molecule de semnalizare care promovează eliberarea secreției liberine (factori de eliberare),și prevenirea eliminării - statine.
Funcțiile receptorului.
Ele sunt furnizate în principal de glicoproteinele situate pe suprafața plasmalemei și capabile să se lege de liganzii lor. Ligandul corespunde receptorului său ca o cheie a unui lacăt. Legarea ligandului la receptor determină o modificare a conformației polipeptidei. Odată cu o astfel de modificare a proteinei transmembranare, se stabilește comunicarea între mediul extracelular și cel intracelular.
Tipuri de receptori.
Receptorii asociați cu canalele ionice proteice. Ei interacționează cu o moleculă de semnalizare care deschide sau închide temporar canalul pentru trecerea ionilor. (De exemplu, receptorul pentru neurotransmițătorul acetilcolină este o proteină formată din 5 subunități care formează un canal ionic. În absența acetilcolinei, canalul este închis, iar după atașare se deschide și permite trecerea ionilor de sodiu).
Receptori catalitici. Ele constau dintr-o parte extracelulară (receptorul însuși) și o parte citoplasmatică intracelulară, care funcționează ca o enzimă prolin kinază (de exemplu, receptorii hormonului de creștere).
Receptorii asociați cu proteinele G. Acestea sunt proteine transmembranare constând dintr-un receptor care interacționează cu un ligand și o proteină G (proteina reglatoare legată de guanozin trifosfat), care transmite un semnal unei enzime legate de membrană (adenilat ciclază) sau unui canal ionic. Ca rezultat, AMP ciclic sau ionii de calciu sunt activați. (Așa funcționează sistemul de adenilat ciclază. De exemplu, în celulele hepatice există un receptor pentru hormonul insulină. Partea supercelulară a receptorului se leagă de insulină. Aceasta determină activarea părții intracelulare - enzima adenilat ciclază. Sintetizează AMP ciclic din ATP, care reglează viteza diferitelor procese intracelulare, determinând activarea sau inhibarea acelor sau a altor enzime metabolice).
Receptorii care percep factorii fizici. De exemplu, proteina fotoreceptor rodopsina. La absorbția luminii, aceasta își schimbă conformația și excită un impuls nervos.
Este format dintr-un strat bilipid, ale cărui lipide sunt strict orientate - partea hidrofobă a lipidelor (coada) este orientată spre interiorul stratului, în timp ce partea hidrofilă (capul) este în exterior. În plus față de lipide, în construcția membranei plasmatice sunt implicate trei tipuri de proteine membranare: periferice, integrale și semi-integrale.
Una dintre domeniile de studiu ale membranelor este în prezent un studiu detaliat al proprietăților atât ale diferitelor lipide structurale și de reglare, cât și ale proteinelor individuale integrale și semi-integrale care alcătuiesc membranele.
Proteinele membranare integrale
Rolul principal în organizarea membranei în sine îl au proteinele integrale și semi-integrale, care au o structură globulară și sunt asociate cu faza lipidică prin interacțiuni hidrofil-hidrofobe. Globulele proteinelor integrale pătrund în toată grosimea membranei, iar partea lor hidrofobă este situată în mijlocul globului și este scufundată în zona hidrofobă a fazei lipidice.
Proteine membranare semi-integrale
În proteinele semi-integrale, aminoacizii hidrofobi sunt concentrați la unul dintre polii globulului și, în consecință, globulele sunt doar pe jumătate scufundate în membrană, ieșind în afară de pe o suprafață (externă sau internă) a membranei.
Funcțiile proteinelor membranare
Proteinelor integrale și semi-integrale ale membranei plasmatice li s-au atribuit anterior două funcții: structurală generală și specifică. În consecință, proteinele structurale și funcționale au fost distinse între ele. Cu toate acestea, îmbunătățirea metodelor de izolare a fracțiilor proteice ale membranelor și o analiză mai detaliată a proteinelor individuale indică acum absența proteinelor structurale universale pentru toate membranele care nu au nicio funcție specifică. În schimb, proteinele membranare cu funcții specifice sunt foarte diverse. Acestea sunt proteine care îndeplinesc funcții de receptor, proteine care sunt purtători activi și pasivi ai diferiților compuși și, în sfârșit, proteine care fac parte din numeroase sisteme enzimatice. Material de pe site
Proprietățile proteinelor membranare
Proprietatea comună a tuturor acestor proteine membranare integrale și semi-integrale, care diferă nu numai ca funcțional, ci și din punct de vedere chimic, este capacitatea lor fundamentală de a se mișca, „pluti” în planul membranei în faza lipidică lichidă. După cum sa menționat mai sus, existența unor astfel de mișcări în membranele plasmatice ale unor celule a fost dovedită experimental. Dar acesta este departe de singurul tip de mișcare identificat în proteinele membranei. Pe lângă deplasarea laterală, proteinele individuale integrale și semi-integrale se pot roti în planul membranei în direcții orizontale și chiar verticale și pot modifica și gradul de imersare al moleculei în faza lipidică.
Opsin. Toate aceste mișcări diverse și complexe ale globulelor proteice sunt în mod deosebit bine ilustrate de exemplul proteinei opsină, care este specifică pentru membranele celulelor fotoreceptoare (Fig. 3). După cum știți, opsina în întuneric este asociată cu retina carotenoid, care conține o legătură dublă ciss; complexul retinian și opsină formează rodopsina sau violetul vizual. Molecula de rodopsina este capabilă de mișcare laterală și rotație în planul orizontal al membranei (Fig. 3, A). Când este expusă la lumină, retina este supusă fotoizomerizării și se transformă într-o formă trans. În acest caz, conformația retinei se modifică și este separată de opsină, care, la rândul său, schimbă planul de rotație de la orizontal la vertical (Fig. 3, B). Consecința unor astfel de transformări este o modificare a permeabilității membranei pentru ioni, ceea ce duce la apariția unui impuls nervos.
Interesant este că modificările conformației globulelor de opsină induse de energia luminoasă nu pot servi doar la generarea unui impuls nervos, așa cum se întâmplă în celulele retinei ochiului, dar sunt și cel mai simplu sistem fotosintetic găsit în bacteriile speciale violete.
Membranele biologice formează baza organizării structurale a celulei. Membrana plasmatică (plasmalema) este membrana care înconjoară citoplasma unei celule vii. Membranele sunt compuse din lipide și proteine. Lipidele (în principal fosfolipide) formează un strat dublu, în care „cozile” hidrofobe ale moleculelor sunt îndreptate spre interiorul membranei, iar cele hidrofile – spre suprafețele acesteia. Moleculele de proteine pot fi localizate pe suprafața exterioară și interioară a membranei, ele se pot scufunda parțial în stratul lipidic sau se pot pătrunde prin și prin el. Majoritatea proteinelor membranare imersate sunt enzime. Acesta este un model fluid-os-mozaic al structurii unei membrane plasmatice. Moleculele de proteine si lipide sunt mobile, ceea ce asigura dinamismul membranei. Membranele conțin și carbohidrați sub formă de glicolipide și glicoproteine (glicocalix) situate pe suprafața exterioară a membranei. Setul de proteine și carbohidrați de pe suprafața membranei fiecărei celule este specific și este un fel de indicator al tipului de celule.
Functiile membranei:
- Separarea. Constă în formarea unei bariere între conținutul interior al celulei și mediul extern.
- Asigurarea schimbului de substante intre citoplasma si mediul extern. Apa, ionii, moleculele anorganice și organice intră în celulă (funcția de transport). Produșii formați în celulă (funcția secretorie) sunt excretați în mediul extern.
- Transport. Transportul prin membrană poate avea loc în moduri diferite. Transportul pasiv se realizează fără consum de energie, prin difuzie simplă, osmoză sau difuzie facilitată cu ajutorul proteinelor purtătoare. Transportul activ se face cu ajutorul proteinelor purtătoare și necesită energie (de exemplu, o pompă de sodiu-potasiu). Material de pe site
Molecule mari de biopolimeri intră în celulă ca urmare a endocitozei. Se împarte în fagocitoză și pinocitoză. Fagocitoza este captarea și absorbția particulelor mari de către celulă. Fenomenul a fost descris pentru prima dată de I.I. Mechnikov. Mai întâi, substanțele aderă la membrana plasmatică, la proteine specifice receptorului, apoi membrana se lasă, formând o depresiune.
Se formează o vacuolă digestivă. Digeră substanțele care au intrat în celulă. La oameni și animale, leucocitele sunt capabile de fagocitoză. Celulele albe din sânge absorb bacteriile și alte particule.
Pinocitoza este procesul de captare și absorbție a picăturilor de lichid cu substanțe dizolvate în ea. Substanțele aderă la proteinele membranei (receptori), iar o picătură de soluție este înconjurată de o membrană, formând o vacuola. Pinocitoza și fagocitoza apar cu cheltuirea energiei ATP.
- Secretar. Secreție - eliberarea de către celulă a substanțelor sintetizate în celulă în mediul extern. Hormonii, polizaharidele, proteinele, picăturile de grăsime sunt închise în vezicule delimitate de o membrană și merg la plasmalemă. Membranele fuzionează, iar conținutul veziculei este descărcat în mediul din jurul celulei.
- Conexiunea celulelor în țesut (datorită excrescențelor pliate).
- Receptor. Membranele conțin un număr mare de receptori - proteine speciale, al căror rol este de a transmite semnale din exterior spre interiorul celulei.
1. Bariera- asigura un metabolism reglat, selectiv, pasiv si activ cu mediul.
Membranele celulare au permeabilitate selectivă: glucoza, aminoacizii, acizii grași, glicerolul și ionii difuzează încet prin ele, membranele în sine reglează activ acest proces - unele substanțe sunt lăsate să treacă, în timp ce altele nu.
2. Transport- prin membrana, substantele sunt transportate in si in afara celulei. Transportul prin membrane asigură: livrarea nutrienților, îndepărtarea produselor metabolice finale, secreția diferitelor substanțe, crearea gradienților ionici, menținerea pH-ului și concentrației ionice adecvate în celulă, care sunt necesare pentru funcționarea enzimelor celulare.
Există patru mecanisme principale pentru intrarea substanțelor în celulă sau îndepărtarea lor din celulă în exterior:
a) Pasiv (difuzie, osmoză) (nu necesită consum de energie)
Difuzie
Răspândirea moleculelor sau atomilor unei substanțe între molecule sau atomi ai alteia, ducând la o egalizare spontană a concentrațiilor acestora în volumul ocupat. În unele situații, una dintre substanțe are deja o concentrație egalizată și se vorbește despre difuzia unei substanțe în alta. În acest caz, transferul de materie are loc dintr-o zonă cu o concentrație mare într-o zonă cu o concentrație scăzută (de-a lungul vectorului gradientului de concentrație (fig. 2.4).
Orez. 2.4. Diagrama procesului de difuzie
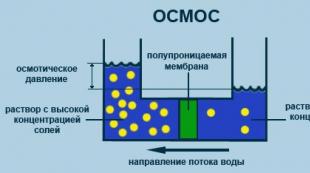
Osmoză
Procesul de difuzie unidirecțională printr-o membrană semipermeabilă a moleculelor de solvent către o concentrație mai mare de substanță dizolvată dintr-un volum cu o concentrație mai mică de substanță dizolvată (fig. 2.5).

Orez. 2.5. Diagrama procesului de osmoză
b) Transport activ (necesită consum de energie)
Pompa sodiu-potasiu- mecanismul de transport transmembranar conjugat activ al ionilor de sodiu (din celulă) și al ionilor de potasiu (în celulă), care asigură un gradient de concentrație și o diferență de potențial transmembranar. Acesta din urmă servește drept bază pentru multe funcții ale celulelor și organelor: secreția de celule glandulare, contracția musculară, conducerea impulsurilor nervoase etc. (fig. 2.6).

Orez. 2.6. Schema pompei de potasiu-sodiu
În prima etapă, enzima Na + / K + -ATPaza atașează trei ioni Na + din partea interioară a membranei. Acești ioni modifică conformația centrului activ al ATPazei. După aceea, enzima este capabilă să hidrolice o moleculă de ATP. Energia eliberată după hidroliză este cheltuită pentru modificarea conformației purtătorului, datorită căreia cei trei ioni Na + și ionul PO 4 3– (fosfat) apar pe partea exterioară a membranei. Aici, ionii Na + sunt despărțiți și PO 4 3– este înlocuit cu doi ioni K +. După aceea, enzima revine la conformația inițială, iar ionii K + apar pe partea interioară a membranei. Aici ionii K + sunt despărțiți, iar purtătorul este din nou gata de lucru.
Ca urmare, în mediul extracelular se creează o concentrație mare de ioni de Na +, iar în interiorul celulei se creează o concentrație mare de K +. Această diferență de concentrație este utilizată în celule în timpul conducerii unui impuls nervos.
c) Endocitoză (fagocitoză, pinocitoză)
Fagocitoză(mâncarea de către celulă) - procesul de absorbție a obiectelor solide de către celulă, cum ar fi celule eucariote, bacterii, viruși, rămășițe de celule moarte etc. În jurul obiectului absorbit se formează un vacuol intracelular mare (fagozom). Dimensiunea fagozomilor este de la 250 nm și mai mult. Prin fuzionarea fagozomului cu lizozomul primar, se formează un lizozom secundar. Într-un mediu acid, enzimele hidrolitice descompun macromoleculele din lizozomul secundar. Produșii de scindare (aminoacizi, monozaharide și alte substanțe utile) sunt apoi transportați prin membrana lizozomală în citoplasma celulei. Fagocitoza este foarte răspândită. La animalele foarte organizate și la oameni, procesul de fagocitoză joacă un rol protector. Activitatea fagocitară a leucocitelor și macrofagelor este de mare importanță în protejarea organismului de microbii patogeni și alte particule nedorite care intră în el. Fagocitoza a fost descrisă pentru prima dată de omul de știință rus I.I.Mechnikov (fig. 2.7)
Pinocitoza(băutură de către celulă) - procesul de absorbție de către celulă a fazei lichide din mediu care conține substanțe solubile, inclusiv molecule mari (proteine, polizaharide etc.). Cu pinocitoza, veziculele mici - endozomi - sunt desprinse de membrana din interiorul celulei. Sunt mai mici decât fagozomii (dimensiunea lor este de până la 150 nm) și de obicei nu conțin particule mari. După formarea endozomului, lizozomul primar se apropie de el, iar aceste două vezicule membranare se contopesc. Organelele rezultate se numesc lizozom secundar. Procesul de pinocitoză este efectuat în mod constant de toate celulele eucariote. (fig. 7)
Endocitoza mediată de receptor - un proces specific activ în care membrana celulară se umflă în celulă, formând gropi margini. Partea intracelulară a fosei căptușite conține un set de proteine adaptive. Macromoleculele care se leagă de receptori specifici de pe suprafața celulei trec în interior cu o viteză mult mai mare decât substanțele care intră în celule prin pinocitoză.

Orez. 2.7. Endocitoza
d) Exocitoză (fagocitoză negativă și pinocitoză)
Un proces celular în care veziculele intracelulare (veziculele membranare) fuzionează cu membrana celulară exterioară. În timpul exocitozei, conținutul veziculelor secretoare (veziculele exocitare) este eliberat în exterior, iar membrana lor se contopește cu membrana celulară. Aproape toți compușii macromoleculari (proteine, hormoni peptidici etc.) sunt eliberați din celulă prin această metodă. (fig. 2.8)

Orez. 2.8. Schema de exocitoză
3. Generarea și conducerea biopotențialelor- cu ajutorul membranei se menține o concentrație constantă de ioni în celulă: concentrația ionului K + în interiorul celulei este mult mai mare decât în exterior, iar concentrația Na + este mult mai mică, ceea ce este foarte important, întrucât aceasta asigură menţinerea diferenţei de potenţial pe membrană şi generarea unui impuls nervos.
4. Mecanic- asigură autonomia celulei, structurile sale intracelulare, precum și legătura cu alte celule (în țesuturi).
5. Energie- în timpul fotosintezei în cloroplaste și a respirației celulare în mitocondrii, în membranele acestora funcționează sisteme de transfer de energie, în care sunt implicate și proteinele;
6. Receptor- unele proteine din membrana sunt receptori (molecule prin care celula percepe anumite semnale).
7. Enzimatic- proteinele membranare sunt adesea enzime. De exemplu, membranele plasmatice ale celulelor epiteliale intestinale conțin enzime digestive.
8. Matrice- asigură o anumită aranjare și orientare reciprocă a proteinelor membranare, interacțiunea optimă a acestora;
9. Marcarea cuștii- exista antigene pe membrana care actioneaza ca markeri - "etichete" care permit identificarea celulei. Acestea sunt glicoproteine (adică proteine cu lanțuri laterale ramificate de oligozaharide atașate) care joacă rolul de „antene”. Cu ajutorul markerilor, celulele pot recunoaște alte celule și pot acționa împreună cu acestea, de exemplu, în timpul formării organelor și țesuturilor. De asemenea, permite sistemului imunitar să recunoască antigenele străine.
Incluziuni celulare
Incluziunile celulare includ carbohidrați, grăsimi și proteine. Toate aceste substanțe se acumulează în citoplasma celulei sub formă de picături și boabe de diferite dimensiuni și forme. Ele sunt sintetizate periodic în celulă și utilizate în procesul metabolic.
Citoplasma
Este o parte a unei celule vii (protoplast) fără membrană plasmatică și nucleu. Compoziția citoplasmei include: matricea citoplasmatică, citoscheletul, organitele și incluziunile (uneori incluziunile și conținutul vacuolelor nu sunt clasificate ca materie vie a citoplasmei). Delimitată de mediul extern de membrana plasmatică, citoplasma este mediul intern semi-lichid al celulelor. Nucleul și diverse organele sunt localizate în citoplasma celulelor eucariote. De asemenea, conține o varietate de incluziuni - produse ale activității celulare, vacuole, precum și cei mai mici tubuli și filamente care formează scheletul celulei. Proteinele predomină în compoziția substanței principale a citoplasmei.
Funcții citoplasmatice
1) în ea au loc principalele procese metabolice.
2) unește nucleul și toate organitele într-un singur întreg, asigură interacțiunea lor.
3) mobilitate, iritabilitate, metabolism și reproducere.
Mobilitatea vine sub diferite forme:
Mișcarea intracelulară a citoplasmei celulare.
Mișcare în formă de amibă. Această formă de mișcare se exprimă în formarea de pseudopodii de către citoplasmă către acest sau acel stimul sau departe de acesta. Această formă de mișcare este inerentă amebei, leucocitelor din sânge, precum și unor celule tisulare.
Mișcare pâlpâitoare. Se manifestă sub formă de bătaie a micilor excrescențe protoplasmatice - cili și flageli (ciliați, celule epiteliale ale animalelor multicelulare, spermatozoizi etc.).
Contractarea mișcării. Este furnizat datorită prezenței în citoplasmă a miofibrilelor organoide speciale, a căror scurtare sau alungire contribuie la contracția și relaxarea celulei. Capacitatea de contractare este cel mai dezvoltată în celulele musculare.
Iritabilitatea este exprimată în capacitatea celulelor de a răspunde la iritație prin modificarea metabolismului și a energiei.
Citoscheletul
Una dintre trăsăturile distinctive ale unei celule eucariote este prezența în citoplasma sa a formațiunilor scheletice sub formă de microtubuli și mănunchiuri de fibre proteice. Elementele citoscheletului, strâns asociate cu membrana citoplasmatică exterioară și învelișul nuclear, formează țesături complexe în citoplasmă.
Citoscheletul este format din microtubuli, microfilamente și sistemul microtrabecular. Citoscheletul determină forma celulei, participă la mișcările celulare, la diviziunea și mișcarea celulei în sine, la transportul intracelular al organitelor.
Microtubuli sunt conținute în toate celulele eucariote și sunt cilindri goali neramificati, al căror diametru nu depășește 30 nm, iar grosimea peretelui este de 5 nm. Acestea pot avea o lungime de până la câțiva micrometri. Dezintegrați și reasamblați ușor. Peretele microtubulilor este compus în principal din subunități de proteină tubulină pliate spiral (fig. 2.09)
Funcțiile microtubulilor:
1) îndeplinesc o funcție de susținere;
2) formează un fus de fisiune; asigura divergenta cromozomilor catre polii celulei; sunt responsabili de mișcarea organelelor celulare;
3) participă la transportul intracelular, secreție, formarea peretelui celular;
4) sunt o componentă structurală a cililor, flagelilor, corpilor bazali și centrioli.
Microfilamente sunt reprezentate de filamente cu diametrul de 6 nm, formate din proteina actină, care este apropiată de actina musculară. Actina reprezintă 10-15% din proteinele totale din celulă. În majoritatea celulelor animale, sub membrana plasmatică însăși se formează o rețea densă de filamente de actină și proteine asociate.
Pe lângă actină, în celulă se găsesc și filamente de miozină. Cu toate acestea, numărul lor este mult mai mic. Datorită interacțiunii actinei și miozinei, are loc contracția musculară. Microfilamentele sunt asociate cu mișcarea întregii celule sau a structurilor sale individuale în interiorul acesteia. În unele cazuri, mișcarea este asigurată numai de filamente de actină, în altele - de actină împreună cu miozina.
Funcțiile microfilamentelor
1) rezistență mecanică
2) permite celulei să-și schimbe forma și să se miște.

Orez. 2.09. Citoscheletul
Organele (sau organele)
Sunt împărțite în non-membrană, cu o singură membrană și cu membrană dublă.
LA organele nemembranare celulele eucariote includ organele care nu au propria lor membrană închisă, și anume: ribozomiși organele construite pe baza microtubulilor de tubulină - centru celular (centrioli)și organele de mișcare (flagela și cili).În celulele majorității organismelor unicelulare și a majorității covârșitoare a plantelor superioare (terestre), centriolii sunt absenți.
LA organite cu o singură membrană raporta: reticul endoplasmatic, aparat Golgi, lizozomi, peroxizomi, sferozomi, vacuole și altele. Toate organitele cu o singură membrană sunt interconectate într-un singur sistem celular. Celulele vegetale au lizozomi speciali, celulele animale au vacuole speciale: digestive, excretoare, contractile, fagocitare, autofagocitare etc.
LA organite cu două membrane raporta mitocondriile și plastidele.
Organele nemembranare
A) Ribozomi- organele găsite în celulele tuturor organismelor. Acestea sunt organite mici, reprezentate de particule globulare cu un diametru de aproximativ 20 nm. Ribozomii sunt formați din două subunități de dimensiuni inegale - mari și mici. Ribozomii includ proteine și ARN ribozomal (ARNr). Există două tipuri principale de ribozomi: eucarioți (80S) și procarioți (70S).
În funcție de localizarea în celulă, există ribozomi liberi localizați în citoplasmă care sintetizează proteine și ribozomi atașați - ribozomi legați prin subunități mari cu suprafața exterioară a membranelor EPR, sintetizând proteine care intră în complexul Golgi, iar apoi sunt secretate de celula. În timpul biosintezei proteinelor, ribozomii se pot combina în complexe - poliribozomi (polizomi).
În nucleol se formează ribozomi eucarioți. Mai întâi, ARNr este sintetizat pe ADN-ul nucleolar, care este apoi acoperit cu proteine ribozomale care provin din citoplasmă, scindate la dimensiunea necesară și formând subunități ribozomale. Nu există ribozomi complet formați în nucleu. Unirea subunităților într-un ribozom întreg are loc în citoplasmă, de obicei în timpul biosintezei proteinelor.
Ribozomii se găsesc în celulele tuturor organismelor. Fiecare constă din două particule, mici și mari. Ribozomii includ proteine și ARN.
Funcții
sinteza proteinei.
Proteinele sintetizate se acumulează mai întâi în canalele și cavitățile reticulului endoplasmatic, apoi sunt transportate în organele și părți ale celulei. EPS și ribozomii localizați pe membranele sale reprezintă un singur aparat pentru biosinteza și transportul proteinelor (fig. 2.10-2.11).

Orez. 2.10. Structura ribozomilor

Orez. 2.11. Structura ribozomilor
B) Centrul celular (centrioli)
Centriolul este un cilindru (0,3 µm lungime și 0,1 µm în diametru), al cărui perete este format din nouă grupuri de trei microtubuli îmbinați (9 tripleți), interconectați la anumite intervale prin reticulare. Centriolii sunt adesea perechi, unde sunt în unghi drept unul față de celălalt. Dacă centriolul se află la baza ciliului sau a flagelului, atunci se numește corpul bazal.
Aproape toate celulele animale au o pereche de centrioli, care sunt elementul mijlociu al centrului celular.
Înainte de divizare, centriolii diverg către poli opuși și un centriol fiică apare lângă fiecare dintre ei. Din centrioli localizați la diferiți poli ai celulei se formează microtubuli, crescând unul spre celălalt.
Funcții
1) formează un fus mitotic, care contribuie la distribuția uniformă a materialului genetic între celulele fiice,
2) sunt centrul organizării citoscheletului. Unele dintre filamentele fusului sunt atașate de cromozomi.
Centriolii sunt organele citoplasmei auto-replicabile. Ele apar ca urmare a dublării celor existente. Acest lucru se întâmplă atunci când centriolii diverg. Centriolul imatur conține 9 microtubuli unici; aparent, fiecare microtubul este o matrice pentru asamblarea tripleților caracteristice unui centriol matur. (fig. 2.12).
Cetriolii se găsesc în celulele plantelor inferioare (alge).

Orez. 2.12. Centriolii din centrul celulei
Organele cu o singură membrană
D) Reticulul endoplasmatic (EPS)
Întreaga zonă interioară a citoplasmei este umplută cu numeroase canale și cavități mici, ai căror pereți sunt membrane similare ca structură cu membrana plasmatică. Aceste canale se ramifică, se conectează între ele și formează o rețea numită reticul endoplasmatic. Reticulul endoplasmatic este heterogen ca structură. Sunt cunoscute două tipuri - granularși neted.
Pe membranele canalelor și cavitățile rețelei granulare, există multe corpuri mici rotunjite - ribozom care conferă membranelor un aspect aspru. Membranele reticulului endoplasmatic neted nu poartă ribozomi pe suprafața lor. EPS îndeplinește multe funcții diferite.
Funcții
Funcția principală a reticulului endoplasmatic granular este participarea la sinteza proteinelor, care se realizează în ribozomi. Pe membranele reticulului endoplasmatic neted se sintetizează lipidele și carbohidrații. Toți acești produși de sinteză se acumulează în canale și cavități, apoi sunt transportați în diverse organite ale celulei, unde sunt consumați sau se acumulează în citoplasmă sub formă de incluziuni celulare. EPS conectează organelele principale ale celulei (fig. 2.13).

Orez. 2.13. Structura reticulului endoplasmatic (EPS) sau reticulului
E) Aparatul Golgi
Structura acestui organoid este similară în celulele organismelor vegetale și animale, în ciuda diversității formei sale. Are multe funcții importante.
Organoid cu o singură membrană. Este un teanc de „cisterne” turtite, cu margini extinse, cu care este asociat un sistem de bule mici cu o singură membrană (bule Golgi). Bulele Golgi sunt concentrate în principal pe partea adiacentă EPS și la periferia stivelor. Se crede că transferă proteine și lipide în aparatul Golgi, ale cărui molecule, deplasându-se de la cisterna la cisternă, suferă modificări chimice.
Toate aceste substanțe se acumulează mai întâi, se complică chimic și apoi, sub formă de bule mari și mici, intră în citoplasmă și fie sunt folosite în celula însăși în timpul vieții, fie sunt îndepărtate din ea și utilizate în organism. (fig. 2.14-2.15).

Orez. 2.14. Structura aparatului Golgi
Funcții:
Modificarea și acumularea de proteine, lipide, carbohidrați;
Ambalarea în vezicule membranare (vezicule) a substanțelor organice care intră;
Locul de formare a lizozomilor;
Funcția secretorie, prin urmare, aparatul Golgi este bine dezvoltat în celulele secretoare.

Orez. 2.15. Complexul Golgi
E) Lizozomi
Sunt corpuri mici rotunjite. În interiorul lizozomului se află enzime care descompun proteinele, grăsimile, carbohidrații, acizii nucleici. Lizozomii se apropie de particula alimentară care a intrat în citoplasmă, se contopesc cu aceasta și se formează o vacuola digestivă, în interiorul căreia se află o particulă alimentară înconjurată de enzime lizozomale.
Enzimele lizozomale sunt sintetizate pe un EPS grosier, transferate în aparatul Golgi, unde sunt modificate și împachetate în vezicule membranare ale lizozomilor. Lizozomul poate conține de la 20 la 60 de tipuri diferite de enzime hidrolitice. Descompunerea substanțelor folosind enzime se numește liza.
Distingeți între lizozomii primari și secundari. Se numesc lizozomi primari care au înmugurit din aparatul Golgi.
Se numesc lizozomi secundari, formați ca urmare a fuziunii lizozomilor primari cu vacuolele endocitare. În acest caz, ele digeră substanțele care au pătruns în celulă prin fagocitoză sau pinocitoză, deci pot fi numite vacuole digestive.
Funcțiile lizozomilor:
1) digestia substanțelor sau particulelor (bacterii, alte celule) captate de celulă în timpul endocitozei,
2) autofagie - distrugerea structurilor inutile pentru celulă, de exemplu, în timpul înlocuirii organitelor vechi cu altele noi sau digestia proteinelor și a altor substanțe produse în interiorul celulei însăși,
3) autoliza - autodigestia unei celule, care duce la moartea acesteia (uneori acest proces nu este patologic, ci însoțește dezvoltarea organismului sau diferențierea unor celule specializate) (fig. 2.16-2.17).
Exemplu: Când un mormoloc se transformă în broască, lizozomii din celulele cozii îl digeră: coada dispare, iar substanțele formate în timpul acestui proces sunt absorbite și utilizate de alte celule ale corpului.

Orez. 2.16. Formarea lizozomilor

Orez. 2.17. Funcționarea lizozomilor
G) Peroxizomi
Organoizi, asemănătoare ca structură cu lizozomii, vezicule cu un diametru de până la 1,5 microni cu o matrice omogenă care conține aproximativ 50 de enzime.
Catalaza determină descompunerea peroxidului de hidrogen 2H 2 O 2 → 2H 2 O + O 2 și previne peroxidarea lipidelor
Peroxizomii sunt formați prin înmugurire din cei existenți anterior, adică. aparțin organelelor care se reproduc singur, în ciuda faptului că nu conțin ADN. Ele cresc datorită pătrunderii enzimelor în ele, enzimele peroxizomale se formează pe EPS aspru și în hialoplasmă (fig. 2.18).

Orez. 2.18. Peroxizom (în centrul nucleoidului cristalin)
H) Vacuole
Organele cu o singură membrană. Vacuolele sunt „recipiente” umplute cu soluții apoase de substanțe organice și anorganice. EPS și aparatul Golgi sunt implicate în formarea vacuolelor.
Celulele plantelor tinere conțin multe vacuole mici, care apoi, pe măsură ce celulele cresc și se diferențiază, se îmbină între ele și formează o singură vacuola centrală mare.
Vacuola centrală poate ocupa până la 95% din volumul unei celule mature, în timp ce nucleul și organelele sunt împinse înapoi către membrana celulară. Se numește membrana care limitează vacuola vegetală tonoplast.
Lichidul care umple vacuola plantei se numește seva celulară. Compoziția sevei celulare include săruri organice și anorganice solubile în apă, monozaharide, dizaharide, aminoacizi, produse metabolice finale sau toxice (glicozide, alcaloizi), unii pigmenți (antociani).
Dintre substanțele organice, zaharurile și proteinele sunt mai des stocate. Zaharuri – mai des sub formă de soluții, proteinele vin sub formă de bule EPR și aparatul Golgi, după care vacuolele sunt deshidratate, transformându-se în boabe de aleuronă.
În celulele animale, există mici vacuole digestive și autofagice aparținând grupului de lizozomi secundari și care conțin enzime hidrolitice. La animalele unicelulare, există și vacuole contractile care îndeplinesc funcția de osmoreglare și excreție.
Funcții
În plante
1) acumularea de lichid și menținerea turgenței,
2) acumularea de nutrienți de rezervă și săruri minerale,
3) colorarea florilor si fructelor si astfel atragerea polenizatorilor si distribuitorilor de fructe si seminte.
La animale:
4) vacuole digestive - distrug macromoleculele organice;
5) vacuolele contractile reglează presiunea osmotică a celulei și elimină substanțele inutile din celulă
6) vacuolele fagocitare se formează în timpul fagocitozei de către celulele imune ale antigenelor
7) vacuolele autofagocitare se formează în timpul fagocitozei de către celulele imune ale propriilor țesuturi
Organele cu două membrane (mitocondrii și plastide)
Aceste organite sunt semi-autonome, deoarece au propriul lor ADN și propriul lor aparat de sinteză a proteinelor. Mitocondriile se găsesc în aproape toate celulele eucariote. Plastidele se găsesc numai în celulele vegetale.
I) Mitocondriile
Acestea sunt organele de alimentare cu energie pentru procesele metabolice din celulă. În hialoplasmă, mitocondriile sunt de obicei distribuite difuz, dar în celulele specializate sunt concentrate în acele zone în care există cea mai mare nevoie de energie. De exemplu, în celulele musculare, un număr mare de mitocondrii este concentrat de-a lungul fibrilelor contractile, de-a lungul flagelului spermatozoid, în epiteliul tubilor renali, în zona sinapselor etc. Această aranjare a mitocondriilor asigură o pierdere mai mică de ATP. în timpul difuzării sale.
Membrana exterioară separă mitocondriile de citoplasmă, este închisă pe ea însăși și nu formează invaginări. Membrana interioară limitează conținutul interior al mitocondriilor - matricea. O trăsătură caracteristică este formarea a numeroase invaginări - cristae, datorită cărora aria membranelor interne crește. Numărul și gradul de dezvoltare a cristelor depind de activitatea funcțională a țesutului. Mitocondriile au propriul lor material genetic (fig. 2.19).
ADN-ul mitocondrial este o moleculă circulară dublu catenară închisă, în celulele umane are o dimensiune de 16569 de perechi de nucleotide, care este de aproximativ 105 ori mai mică decât ADN-ul localizat în nucleu. Mitocondriile au propriul sistem de sinteză a proteinelor, în timp ce numărul de proteine traduse din ARNm mitocondrial este limitat. ADN-ul mitocondrial nu poate codifica toate proteinele mitocondriale. Majoritatea proteinelor din mitocondrii sunt sub controlul genetic al nucleului.

Orez. 2.19. Structura mitocondrială
Funcțiile mitocondriale
1) formarea ATP
2) sinteza proteinelor
3) participarea la sinteze specifice, de exemplu, sinteza hormonilor steroizi (glandele suprarenale)
4) mitocondriile uzate pot acumula, de asemenea, produse de excreție, substanțe nocive, de exemplu. capabile să preia funcțiile altor organite celulare
K) Plastide
Plastide–Organele, caracteristice doar plantelor.
Există trei tipuri de plastide:
1) cloroplaste(plastide verzi);
2) cromoplaste(plastide galbene, portocalii sau roșii)
3) leucoplaste(plastide incolore).
De obicei, într-o celulă se găsește un singur tip de plastide.
Cloroplaste
Aceste organite se găsesc în celulele frunzelor și a altor organe ale plantelor verzi, precum și într-o varietate de alge. Plantele superioare conțin de obicei câteva zeci de cloroplaste într-o singură celulă. Culoarea verde a cloroplastelor depinde de conținutul de pigment de clorofilă din acestea.
Cloroplastul este principalul organoid al celulelor vegetale, în care are loc fotosinteza, adică formarea de substanțe organice (carbohidrați) din anorganice (CO 2 și H 2 O) folosind energia luminii solare. Cloroplastele sunt similare structural cu mitocondriile.
Cloroplastele au o structură complexă. Sunt delimitate de hialoplasmă de două membrane - externă și internă. Conținutul interior se numește stroma... Membrana interioară formează un sistem complex, strict ordonat de membrane sub formă de bule plate, numite tilacoizi.
Tilacoizii sunt adunați în grămezi - boabe asemănătoare coloanelor de monede . Grana este conectată între ele prin tilacoizi stromale, trecând prin ele de-a lungul plastidei (fig. 2.20-2.22). Clorofila și cloroplastele sunt produse numai prin expunerea la lumină.

Orez. 2.20. Cloroplaste la microscop cu lumină

Orez. 2.21. Structura cloroplastului la microscop electronic

Orez. 2.22. Structura schematică a cloroplastelor
Funcții
1) fotosinteză(formarea substantelor organice din substante anorganice datorita energiei luminii). Clorofila joacă un rol central în acest proces. Absoarbe energia luminii și o direcționează pentru a desfășura reacții fotosintetice. În cloroplaste, ca și în mitocondrii, se sintetizează ATP.
2) participă la sinteza aminoacizilor și acizilor grași,
3) servesc drept depozit pentru rezervele temporare de amidon.
Leucoplaste- plastide mici incolore care se găsesc în celulele organelor ascunse de lumina soarelui (rădăcini, rizomi, tuberculi, semințe). Structura lor este similară cu structura cloroplastelor. (fig. 2.23).
Cu toate acestea, spre deosebire de cloroplaste, leucoplastele au un sistem membranar intern slab dezvoltat, deoarece sunt implicați în sinteza și acumularea nutrienților de rezervă - amidon, proteine și lipide. La lumină, leucoplastele se pot transforma în cloroplaste.

Orez. 2.23. Structura leucoplastelor
Cromoplastele- plastide de culoare portocalie, roșie și galbenă, care este cauzată de pigmenții aparținând grupului de carotenoizi. Cromoplastele se găsesc în celulele petalelor multor plante, fructe mature, rar culturi rădăcinoase, precum și în frunzele de toamnă. Sistemul membranei interne în cromoplaste, de regulă, este absent (fig. 24).

Orez. 2.24. Structura cromoplastului
Semnificația cromoplastelor nu a fost încă pe deplin elucidată. Majoritatea sunt plastide îmbătrânite. Ele, de regulă, se dezvoltă din cloroplaste, în timp ce clorofila și structura membranei interioare sunt distruse în plastide, iar carotenoizii se acumulează. Acest lucru se întâmplă când fructele se coc și frunzele se îngălbenesc toamna. Semnificația biologică a cromoplastelor este că ele provoacă culoarea strălucitoare a florilor și fructelor, care atrage insectele pentru polenizare încrucișată și alte animale pentru răspândirea fructelor. Leucoplastele se pot transforma și în cromoplaste.
Funcțiile plastidelor
Sinteza substanțelor organice din clorofilă din compuși anorganici simpli: dioxid de carbon și apă în prezența cuantelor de lumină solară - fotosinteză, Sinteza ATP în faza luminoasă a fotosintezei
Sinteza proteinelor pe ribozomi (între membranele interioare ale cloroplastei se află ADN, ARN și ribozomi, prin urmare, în cloroplaste, precum și în mitocondrii, se sintetizează proteine, care sunt necesare activității acestor organite).
Prezența cromoplastelor se explică prin culoarea galbenă, portocalie și roșie a corolei florilor, fructelor, frunzelor de toamnă.
Leucoplastele conțin substanțe de depozitare (în tulpini, rădăcini, tuberculi).
Cloroplastele, cromoplastele și leucoplastele sunt capabile de tranziție intercelulară. Deci, când fructele se coc sau își schimbă culoarea frunzelor toamna, cloroplastele se transformă în cromoplaste, iar leucoplastele se pot transforma în cloroplaste, de exemplu, când tuberculii de cartofi devin verzi.
În sens evolutiv, tipul primar, inițial, de plastide sunt cloroplastele, din care au provenit plastidele celorlalte două tipuri. Plastidele au multe asemănări cu mitocondriile, deosebindu-le de alte componente ale citoplasmei. Aceasta este, în primul rând, o înveliș de două membrane și o autonomie genetică relativă, datorită prezenței propriilor ribozomi și ADN. Această particularitate a organitelor a stat la baza ideii că precursorii plastidelor și mitocondriilor au fost bacterii, care în procesul de evoluție s-au dovedit a fi construite într-o celulă eucariotă și s-au transformat treptat în cloroplaste și mitocondrii. (fig. 2.25).

Orez. 2.25. Formarea mitocondriilor și a cloroplastelor conform teoriei simbiogenezei
Celulă- o unitate elementară a unui sistem viu. Diverse structuri ale unei celule vii, care sunt responsabile pentru îndeplinirea unei anumite funcții, sunt numite organele, ca și organele întregului organism. Funcțiile specifice în celulă sunt distribuite între organele, structuri intracelulare care au o anumită formă, precum nucleul celular, mitocondriile etc.
Structuri celulare:
Citoplasma... Partea obligatorie a celulei, închisă între membrana plasmatică și nucleu. Citosol Este o soluție apoasă vâscoasă de diferite săruri și substanțe organice, pătrunsă de un sistem de filamente proteice - citoschelete. Majoritatea proceselor chimice și fiziologice ale celulei au loc în citoplasmă. Structura: Citosol, citoschelet. Funcții: include diverse organite, mediul intern al celulei
Membrană plasmatică... Fiecare celulă de animale, plante, este limitată de mediu sau de alte celule de membrana plasmatică. Grosimea acestei membrane este atât de mică (aproximativ 10 nm) încât poate fi văzută doar cu un microscop electronic.

Lipidele formează un strat dublu în membrană, iar proteinele pătrund în toată grosimea acesteia, sunt scufundate la diferite adâncimi în stratul lipidic sau sunt situate pe suprafețele exterioare și interioare ale membranei. Structura membranelor tuturor celorlalte organite este similară cu membrana plasmatică. Structura: strat dublu de lipide, proteine, carbohidrati. Functii: limitarea, mentinerea formei celulei, protejarea impotriva deteriorarii, reglarea aportului si indepartarea substantelor.
Lizozomi... Lizozomii sunt organite membranare. Au o formă ovală și un diametru de 0,5 microni. Conțin un set de enzime care distrug materia organică. Membrana lizozomului este foarte puternică și împiedică pătrunderea propriilor enzime în citoplasma celulei, dar dacă lizozomul este deteriorat de orice influențe externe, atunci întreaga celulă sau o parte a acesteia este distrusă.
Lizozomii se găsesc în toate celulele plantelor, animalelor și ciupercilor.
Digerând diferite particule organice, lizozomii furnizează „materie primă” suplimentară pentru procesele chimice și energetice din celulă. În timpul înfometării, celulele lizozomului digeră unele organele fără a ucide celula. Această digestie parțială oferă celulei minimul necesar de nutrienți pentru o perioadă de timp. Uneori, lizozomii digeră celule întregi și grupuri de celule, ceea ce joacă un rol esențial în dezvoltarea animalelor. Un exemplu este pierderea cozii atunci când un mormoloc se transformă într-o broască. Structura: vezicule de forma ovala, membrana in exterior, enzime in interior. Funcții: scindarea materiei organice, distrugerea organelelor moarte, distrugerea celulelor uzate.
Complexul Golgi... Produșii de biosinteză care intră în lumenele cavităților și tubilor reticulului endoplasmatic sunt concentrați și transportați în aparatul Golgi. Acest organoid are o dimensiune de 5-10 µm.

Structura: cavități (vezicule) înconjurate de membrană. Funcții: acumulare, ambalare, excreție de materie organică, formare de lizozomi
Reticulul endoplasmatic... Reticulul endoplasmatic este un sistem pentru sinteza și transportul substanțelor organice în citoplasma celulei, care este o structură deschisă a cavităților conectate.
Un număr mare de ribozomi sunt atașați de membranele reticulului endoplasmatic - cele mai mici organite ale celulei, care arată ca o sferă cu un diametru de 20 nm. și constând din ARN și proteine. Sinteza proteinelor are loc pe ribozomi. Apoi proteinele nou sintetizate intră în sistemul de cavități și tubuli, de-a lungul cărora se deplasează în interiorul celulei. Cavități, tubuli, tubuli din membrane, pe suprafața membranelor ribozomale. Funcții: sinteza substanțelor organice cu ajutorul ribozomilor, transportul substanțelor.
Ribozomi... Ribozomii sunt atașați de membranele reticulului endoplasmatic sau sunt localizați liber în citoplasmă, sunt localizați în grupuri, proteinele sunt sintetizate pe ele. Compoziția proteinelor, ARN ribozomal Funcții: asigură biosinteza proteinelor (asamblarea unei molecule proteice din).
mitocondriile... Mitocondriile sunt organite energetice. Forma mitocondriilor este diferită, ele pot fi restul, în formă de tijă, filamentoase cu un diametru mediu de 1 micron. si o lungime de 7 microni. Numărul de mitocondrii depinde de activitatea funcțională a celulei și poate ajunge la zeci de mii în mușchii zburători ai insectelor. Mitocondriile sunt delimitate la exterior de membrana exterioară, dedesubt este membrana interioară, care formează numeroase excrescențe - cristae.

În interiorul mitocondriilor se află ARN, ADN și ribozomi. În membranele sale sunt încorporate enzime specifice, cu ajutorul cărora energia substanțelor alimentare este transformată în energia ATP, care este necesară pentru viața celulei și a organismului în ansamblu, în mitocondrii.
Membrană, matrice, excrescențe - cristae. Funcții: sinteza moleculei ATP, sinteza propriilor proteine, acizi nucleici, carbohidrați, lipide, formarea propriilor ribozomi.
Plastide... Doar într-o celulă vegetală: lecoplaste, cloroplaste, cromoplaste. Funcții: acumularea de substanțe organice de rezervă, atragerea insectelor polenizatoare, sinteza de ATP și carbohidrați. Cloroplastele au forma unui disc sau sferă cu un diametru de 4–6 µm. Cu membrană dublă - exterioară și interioară. În interiorul cloroplastei există ADN ribozom și structuri membranare speciale - boabe, conectate între ele și cu membrana interioară a cloroplastei. Fiecare cloroplast conține aproximativ 50 de boabe, eșalonate pentru o captare mai bună a luminii. Clorofila este localizată în membranele granițe, datorită acesteia, energia luminii solare este transformată în energia chimică a ATP. Energia ATP este utilizată în cloroplaste pentru sinteza compușilor organici, în principal carbohidrați.
Cromoplastele... Pigmenții roșii și galbeni găsiți în cromoplaste conferă diferitelor părți ale plantei o culoare roșie și galbenă. morcovi, fructe de roșii.
Leucoplastele sunt un loc de acumulare a unui nutrient de rezervă - amidonul. Există mai ales multe leucoplaste în celulele tuberculilor de cartofi. La lumină, leucoplastele se pot transforma în cloroplaste (în urma cărora celulele cartofului devin verzi). Toamna, cloroplastele se transformă în cromoplaste, iar frunzele și fructele verzi devin galbene și roșii.
Centrul celular... Este format din doi cilindri, centrioli, situati perpendicular unul pe altul. Funcții: suport de filet pentru axele divizoare

Incluziunile celulare apar în citoplasmă, apoi dispar în procesul vieții celulare.

Incluziunile dense sub formă de granule conțin nutrienți de rezervă (amidon, proteine, zaharuri, grăsimi) sau produse reziduale ale celulei, care nu pot fi încă îndepărtate. Toate plastidele celulelor vegetale au capacitatea de a sintetiza și acumula nutrienți de rezervă. În celulele vegetale, acumularea de nutrienți de rezervă are loc în vacuole.
Boabe, granule, picături Funcții: formațiuni nepermanente care stochează materie organică și energie
Miez... Membrană nucleară a două membrane, suc nuclear, nucleol. Funcții: stocarea informațiilor ereditare în celulă și reproducerea acesteia, sinteza ARN - informațional, de transport, ribozomal. În membrana nucleară sunt spori, prin care are loc un schimb activ de substanțe între nucleu și citoplasmă. Nucleul stochează informații ereditare nu numai despre toate semnele și proprietățile unei celule date, despre procesele care trebuie să apară acesteia (de exemplu, sinteza proteinelor), ci și despre caracteristicile organismului în ansamblu. Informațiile sunt înregistrate în moleculele de ADN, care sunt partea principală a cromozomilor. Există un nucleol în nucleu. Nucleul, datorită prezenței în el a cromozomilor care conțin informații ereditare, îndeplinește funcțiile unui centru care controlează toată activitatea vitală și dezvoltarea celulei.