უჯრედის მემბრანის ფენა. უჯრედის მემბრანის სტრუქტურა და ფუნქციები. მნიშვნელობა და დასკვნები
ბიოლოგიური გარსები- რთული სუპრამოლეკულური სტრუქტურები, რომლებიც აკრავს ყველა ცოცხალ უჯრედს და ქმნიან მათში დახურულ, სპეციალიზებულ განყოფილებებს - ორგანელებს.
მემბრანას, რომელიც გარედან ზღუდავს უჯრედის ციტოპლაზმას, ეწოდება ციტოპლაზმური ან პლაზმური მემბრანა. უჯრედშიდა მემბრანების სახელწოდება ჩვეულებრივ მომდინარეობს იმ სუბუჯრედული სტრუქტურების სახელიდან, რომელსაც ისინი ზღუდავენ ან ქმნიან.
გამოარჩევენ:
ბირთვული
მიტოქონდრიული,
ლიზოსომური მემბრანა
გოლგის კომპლექსის გარსები
ენდოპლაზმური რეტიკულუმი და სხვა.
მემბრანაარის თხელი სტრუქტურა 7 ნმ სისქით.
მისი ქიმიური შემადგენლობის მიხედვით, მემბრანა შეიცავს:
25% ცილები,
25% ფოსფოლიპიდები,
13% ქოლესტერინი,
4% ლიპიდები,
3% ნახშირწყლები.
სტრუქტურულადმემბრანის საფუძველია ფოსფოლიპიდების ორმაგი ფენა.
ფოსფოლიპიდური მოლეკულების თავისებურება ის არის, რომ მათ შემადგენლობაში აქვთ ჰიდროფილური და ჰიდროფობიური ნაწილები. ჰიდროფილური ნაწილები შეიცავს პოლარულ ჯგუფებს (ფოსფატის ჯგუფები ფოსფოლიპიდებში და ჰიდროქსიდის ჯგუფები ქოლესტეროლში). ჰიდროფილური ნაწილებიმიმართულია ზედაპირისკენ. ა ჰიდროფობიური (ცხიმოვანი კუდები) მიმართულია მემბრანის ცენტრისკენ.
მოლეკულას აქვს ორი ცხიმოვანი კუდი, და ეს ნახშირწყალბადის ჯაჭვები გვხვდება ორ კონფიგურაციაში. გაჭიმული - ტრანს კონფიგურაცია(ცილინდრი 0,48 ნმ). მეორე ტიპი არის gosh-trans-gosh კონფიგურაცია. ამ შემთხვევაში, ორი ცხიმოვანი კუდი განსხვავდება და ფართობი იზრდება 0,58 ნმ-მდე.
ლიპიდურ მოლეკულებს ნორმალურ პირობებში აქვთ თხევადი კრისტალური ფორმა. და ამ მდგომარეობაში მათ აქვთ მობილურობა. უფრო მეტიც, მათ შეუძლიათ ორივე გადაადგილდნენ თავიანთ ფენაში და გადაბრუნდნენ. როდესაც ტემპერატურა იკლებს, მემბრანის თხევადი მდგომარეობიდან ჟელეს მსგავსზე გადასვლა ხდება და ეს ამცირებს მოლეკულის მობილურობას.
როდესაც ლიპიდური მოლეკულა მოძრაობს, წარმოიქმნება მიკროზოლები, რომლებსაც უწოდებენ მეფეებს, რომლებშიც ნივთიერებები შეიძლება იყოს ხაფანგში.. მემბრანის ლიპიდური ფენა არის ბარიერი წყალში ხსნადი ნივთიერებებისთვის, მაგრამ ის საშუალებას აძლევს ცხიმში ხსნად ნივთიერებებს გაიარონ..
დახურული ლიპიდური ორფენი განსაზღვრავს მემბრანების ძირითად თვისებებს:
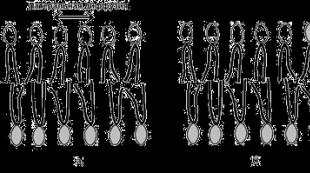
1) სითხე- დამოკიდებულია მემბრანული ლიპიდების შემადგენლობაში გაჯერებული და უჯერი ცხიმოვანი მჟავების თანაფარდობაზე. გაჯერებული ცხიმოვანი მჟავების ჰიდროფობიური ჯაჭვები ორიენტირებულია ერთმანეთის პარალელურად და ქმნიან მყარ კრისტალურ სტრუქტურას (სურათი 14.8, ა). უჯერი ცხიმოვანი მჟავები, რომლებსაც აქვთ მოხრილი ნახშირწყალბადის ჯაჭვი, არღვევს შეფუთვის კომპაქტურობას და ხდის მემბრანას უფრო თხევადს (სურათი 14.8, ბ). ქოლესტერინი, რომელიც ცხიმოვან მჟავებს შორის არის ჩასმული, ახდენს მათ კონდენსაციას და ზრდის მემბრანების სიმტკიცეს.
სურათი 14.8. ფოსფოლიპიდების ცხიმოვანი მჟავების შემადგენლობის გავლენა ლიპიდური ორშრის სითხეზე.
2) გვერდითი დიფუზია- მოლეკულების თავისუფალი მოძრაობა ერთმანეთთან შედარებით მემბრანების სიბრტყეში (სურათი 14.9, ა).

სურათი 14.9. ფოსფოლიპიდური მოლეკულების მოძრაობის სახეები ლიპიდურ ორ შრეში.
3) განივი დიფუზიის შეზღუდული შესაძლებლობები(მოლეკულების გადასვლა გარე შრედან შიდა და პირიქით, იხილეთ სურათი 14.9, ბ), რაც ხელს უწყობს შენარჩუნებას ასიმეტრიები- სტრუქტურული და ფუნქციური განსხვავებები მემბრანის გარე და შიდა ფენებს შორის.
4) შეუღწევადობაწყალში ხსნადი მოლეკულების უმეტესობისთვის დახურული ორფენა.
ლიპიდების გარდა, მემბრანა შეიცავს ცილის მოლეკულებსაც. ძირითადად გლიკოპროტეინები.
ინტეგრალური ცილები გადის ორივე ფენაში. სხვა ცილები ნაწილობრივ ჩაეფლო გარე ან შიდა ფენაში. მათ პერიფერიულ ცილებს უწოდებენ..
მემბრანის ამ მოდელს ე.წ თხევადი ბროლის მოდელი. ფუნქციურად ცილის მოლეკულები ასრულებენ სტრუქტურულ, სატრანსპორტო, ფერმენტულ ფუნქციებს. გარდა ამისა, ისინი ქმნიან იონურ არხებს დიამეტრით 0,35-დან 0,8 ნმ-მდე, რომლებშიც იონები შეიძლება გაიარონ. არხებს აქვთ საკუთარი სპეციალობა. ინტეგრალური ცილები მონაწილეობენ აქტიურ ტრანსპორტში და ხელს უწყობენ დიფუზიას.
მემბრანის შიგნითა პერიფერიული ცილები ხასიათდება ფერმენტული ფუნქციით. შიგნით - ანტიგენური (ანტისხეულები) და რეცეპტორული ფუნქციები.
ნახშირბადის ჯაჭვებიშეუძლია ცილის მოლეკულებთან მიმაგრება და შემდეგ ჩამოყალიბება გლიკოპროტეინები. ან ლიპიდებს, მაშინ მათ ეძახიან გლიკოლიპიდები.
ძირითადი ფუნქციებიუჯრედის მემბრანა იქნება:
1. ბარიერის ფუნქცია(გამოიხატება იმაში, რომ მემბრანა შესაბამისი მექანიზმების დახმარებით მონაწილეობს კონცენტრაციის გრადიენტების შექმნაში, ხელს უშლის თავისუფალ დიფუზიას. ამავდროულად მემბრანა მონაწილეობს ელექტროგენეზის მექანიზმებში. მათ შორისაა მექანიზმები, რომლებიც ქმნიან დასვენების პოტენციალი, მოქმედების პოტენციალის წარმოქმნა, ბიოელექტრული იმპულსების გავრცელების მექანიზმები ერთგვაროვან და არაერთგვაროვან აგზნებად სტრუქტურებზე.)
2. ნივთიერების გადაცემა.

სურათი 14.10.მემბრანის გასწვრივ მოლეკულების ტრანსპორტირების მექანიზმები
მარტივი დიფუზია- ნივთიერებების გადატანა მემბრანის მეშვეობით სპეციალური მექანიზმების მონაწილეობის გარეშე. ტრანსპორტი ხდება კონცენტრაციის გრადიენტის გასწვრივ ენერგიის მოხმარების გარეშე. მარტივი დიფუზიით ხდება მცირე ბიომოლეკულების ტრანსპორტირება - H 2 O, CO 2, O 2, შარდოვანა, ჰიდროფობიური დაბალი მოლეკულური წონის ნივთიერებები. მარტივი დიფუზიის სიჩქარე კონცენტრაციის გრადიენტის პროპორციულია.
გაადვილებული დიფუზია- ნივთიერებების გადატანა მემბრანაში ცილოვანი არხების ან სპეციალური გადამზიდავი ცილების გამოყენებით. იგი ხორციელდება კონცენტრაციის გრადიენტის გასწვრივ ენერგიის მოხმარების გარეშე. ტრანსპორტირდება მონოსაქარიდები, ამინომჟავები, ნუკლეოტიდები, გლიცეროლი, ზოგიერთი იონი. დამახასიათებელია გაჯერების კინეტიკა - გადატანილი ნივთიერების გარკვეული (გაჯერებული) კონცენტრაციის დროს გადაცემაში მონაწილეობს ყველა გადამზიდავი მოლეკულა და ტრანსპორტირების სიჩქარე აღწევს ზღვრულ მნიშვნელობას.
აქტიური ტრანსპორტი- ასევე მოითხოვს სპეციალური გადამზიდავი ცილების მონაწილეობას, მაგრამ გადატანა ხდება კონცენტრაციის გრადიენტის საწინააღმდეგოდ და ამიტომ მოითხოვს ენერგიას. ამ მექანიზმის დახმარებით Na +, K +, Ca 2+, Mg 2+ იონები ტრანსპორტირდება უჯრედის მემბრანაში, პროტონები კი მიტოქონდრიის მემბრანის გავლით. ნივთიერებების აქტიური ტრანსპორტირებას ახასიათებს გაჯერების კინეტიკა.
მატარებლების მიერ განხორციელებული ორგანული ნივთიერებებისა და იონების ტრანსპორტირებასთან ერთად, უჯრედში არის ძალიან სპეციალური მექანიზმი, რომელიც შექმნილია უჯრედიდან მაკრომოლეკულური ნაერთების შთანთქმისა და ამოღების მიზნით ბიომემბრანის ფორმის შეცვლით. ასეთ მექანიზმს ე.წ ვეზიკულური ტრანსპორტი.
სურათი 14.12.ვეზიკულური ტრანსპორტის სახეები: 1 - ენდოციტოზი; 2 - ეგზოციტოზი.
მაკრომოლეკულების გადაცემის დროს ხდება მემბრანით გარშემორტყმული ვეზიკულების (ვეზიკულების) თანმიმდევრული წარმოქმნა და შერწყმა. ტრანსპორტირების მიმართულებისა და გადატანილი ნივთიერებების ბუნების მიხედვით განასხვავებენ ვეზიკულური ტრანსპორტის შემდეგ ტიპებს:
ენდოციტოზი(სურათი 14.12, 1) - ნივთიერებების გადატანა უჯრედში. მიღებული ვეზიკულების ზომიდან გამომდინარე, არსებობს:
ა) პინოციტოზი - თხევადი და გახსნილი მაკრომოლეკულების (ცილები, პოლისაქარიდები, ნუკლეინის მჟავები) შეწოვა პატარა ბუშტების გამოყენებით (150 ნმ დიამეტრის);
ბ) ფაგოციტოზი - დიდი ნაწილაკების შეწოვა, როგორიცაა მიკროორგანიზმები ან უჯრედის ნამსხვრევები. ამ შემთხვევაში წარმოიქმნება დიდი ვეზიკულები, რომლებსაც 250 ნმ-ზე მეტი დიამეტრის მქონე ფაგოსომები ეწოდება.
პინოციტოზი დამახასიათებელია ევკარიოტული უჯრედების უმეტესობისთვის, ხოლო მსხვილ ნაწილაკებს შეიწოვება სპეციალიზებული უჯრედები - ლეიკოციტები და მაკროფაგები. ენდოციტოზის პირველ ეტაპზე მემბრანის ზედაპირზე შეიწოვება ნივთიერებები ან ნაწილაკები, ეს პროცესი ხდება ენერგიის მოხმარების გარეშე. შემდეგ ეტაპზე მემბრანა ადსორბირებული ნივთიერებით ღრმავდება ციტოპლაზმაში; პლაზმური მემბრანის ლოკალური ინვაგინაციები იჭრება უჯრედის ზედაპირიდან, ქმნიან ვეზიკულებს, რომლებიც შემდეგ უჯრედში გადადიან. ეს პროცესი დაკავშირებულია მიკროფილამენტების სისტემით და ენერგიაზეა დამოკიდებული. უჯრედში შემავალი ვეზიკულები და ფაგოსომები შეიძლება შეერწყას ლიზოსომებს. ლიზოსომებში შემავალი ფერმენტები ანადგურებს ბუშტუკებსა და ფაგოსომებში შემავალ ნივთიერებებს დაბალმოლეკულურ პროდუქტებად (ამინომჟავები, მონოსაქარიდები, ნუკლეოტიდები), რომლებიც ტრანსპორტირდება ციტოზოლში, სადაც მათი გამოყენება უჯრედს შეუძლია.
ეგზოციტოზი(სურათი 14.12, 2) - ნაწილაკების და დიდი ნაერთების გადატანა უჯრედიდან. ეს პროცესი, ისევე როგორც ენდოციტოზი, მიმდინარეობს ენერგიის შთანთქმით. ეგზოციტოზის ძირითადი ტიპებია:
ა) სეკრეცია - წყალში ხსნადი ნაერთების უჯრედიდან ამოღება, რომლებიც გამოიყენება ან გავლენას ახდენენ სხეულის სხვა უჯრედებზე. მისი განხორციელება შესაძლებელია როგორც არასპეციალიზებული უჯრედებით, ასევე ენდოკრინული ჯირკვლების, კუჭ-ნაწლავის ტრაქტის ლორწოვანი გარსის უჯრედებით, რომლებიც ადაპტირებულია მათ მიერ წარმოქმნილი ნივთიერებების სეკრეციისთვის (ჰორმონები, ნეიროტრანსმიტერები, პროენზიმები), სხეულის სპეციფიკური საჭიროებიდან გამომდინარე. .
საიდუმლო ცილები სინთეზირდება რიბოსომებზე, რომლებიც დაკავშირებულია უხეში ენდოპლაზმური ბადის მემბრანებთან. ეს ცილები შემდეგ ტრანსპორტირდება გოლჯის აპარატში, სადაც ხდება მათი მოდიფიცირება, კონცენტრირება, დახარისხება და შემდეგ შეფუთვა ვეზიკულებში, რომლებიც იჭრება ციტოზოლში და შემდგომში ერწყმის პლაზმურ მემბრანას ისე, რომ ვეზიკულების შიგთავსი უჯრედის გარეთაა.
მაკრომოლეკულებისგან განსხვავებით, მცირე სეკრეციული ნაწილაკები, როგორიცაა პროტონები, ტრანსპორტირდება უჯრედიდან გაადვილებული დიფუზიისა და აქტიური სატრანსპორტო მექანიზმების გამოყენებით.
ბ) გამოყოფა - უჯრედიდან იმ ნივთიერებების ამოღება, რომელთა გამოყენებაც შეუძლებელია (მაგალითად, რეტიკულოციტებიდან ერითროპოეზის დროს რეტიკულური ნივთიერების ამოღება, რომელიც წარმოადგენს ორგანელების აგრეგირებულ ნარჩენს). გამოყოფის მექანიზმი, როგორც ჩანს, მდგომარეობს იმაში, რომ თავდაპირველად გამოყოფილი ნაწილაკები ციტოპლაზმურ ვეზიკულაშია, რომელიც შემდეგ ერწყმის პლაზმურ მემბრანას.
3. მეტაბოლური ფუნქცია(მათში ფერმენტული სისტემების არსებობის გამო)
4. მემბრანები ჩართულია ელექტრული პოტენციალის შექმნადასვენებისას და როცა აღელვებული - მოქმედების დენები.
5. რეცეპტორის ფუნქცია.
6. იმუნოლოგიური(ასოცირებულია ანტიგენების არსებობასთან და ანტისხეულების გამომუშავებასთან).
7. უზრუნველყოს უჯრედშორისი ურთიერთქმედება და კონტაქტის დათრგუნვა. (როდესაც ერთგვაროვანი უჯრედები შედიან კონტაქტში, უჯრედების გაყოფა ითრგუნება. ეს ფუნქცია იკარგება კიბოს უჯრედებში. გარდა ამისა, კიბოს უჯრედები შედიან კონტაქტში არა მხოლოდ საკუთარ უჯრედებთან, არამედ სხვა უჯრედებთანაც და აინფიცირებენ მათ.)
რეცეპტორები, მათი კლასიფიკაცია: ლოკალიზაციით (მემბრანული, ბირთვული), პროცესების განვითარების მექანიზმით (იონოტროპული და მეტაიოტროპული), სიგნალის მიღების სიჩქარით (სწრაფი, ნელი), მიმღები ნივთიერებების ტიპის მიხედვით.
რეცეპტორებიარის საბოლოო სპეციალიზებული წარმონაქმნები, რომლებიც შექმნილია სხვადასხვა ტიპის სტიმულის ენერგიის გადაქცევისთვის ნერვული სისტემის სპეციფიკურ აქტივობად.
კლასიფიკაცია:
ლოკალიზაციით
მემბრანა
ბირთვული
პროცესის განვითარების მექანიზმის მიხედვით
იონოტროპული (ეს არის მემბრანული არხები, რომლებიც იხსნება ან იხურება ლიგანდთან შეერთებისას. შედეგად მიღებული იონური დინებები იწვევს ტრანსმემბრანული პოტენციალის სხვაობის ცვლილებას და, შედეგად, უჯრედის აგზნებადობას, ასევე ცვლის იონების უჯრედშიდა კონცენტრაციას, რაც მეორადად შეიძლება გამოიწვიოს უჯრედშიდა მედიატორული სისტემების გააქტიურება ერთ-ერთი ყველაზე სრულად შესწავლილი იონოტროპული რეცეპტორი არის n-ქოლინერგული რეცეპტორი.)
მეტაბოტროპული (დაკავშირებულია უჯრედშიდა შუამავლების სისტემებთან. მათი კონფორმაციის ცვლილებები ლიგანდთან შეკავშირებისას იწვევს ბიოქიმიური რეაქციების კასკადის დაწყებას და, საბოლოო ჯამში, უჯრედის ფუნქციური მდგომარეობის ცვლილებას).
სიგნალის მიღების სიჩქარით
სწრაფი
ნელი
შთამნთქმელი ტიპის მიხედვით
· ქიმიორეცეპტორები- აღიქვამენ გახსნილი ან აქროლადი ქიმიკატების ზემოქმედებას.
· ოსმორეცეპტორები- აღიქვამს ცვლილებები სითხის ოსმოსურ კონცენტრაციაში (როგორც წესი, შინაგანი გარემო).
· მექანიკური რეცეპტორები- აღიქვამს მექანიკურ სტიმულს (შეხება, წნევა, გაჭიმვა, წყლის ან ჰაერის ვიბრაცია და ა.შ.)
· ფოტორეცეპტორები- აღიქვამენ ხილულ და ულტრაიისფერ შუქს
· თერმორეცეპტორები- აღიქვამენ ტემპერატურის შემცირებას (ცივას) ან მატებას (თერმული).
· ბარორეცეპტორები- აღიქვამენ წნევის ცვლილებებს
3. იონოტროპული რეცეპტორები, მეტაბოპროპის რეცეპტორები და მათი ჯიშები. მეტაბოტროპული რეცეპტორების მოქმედების მეორადი შუამავლების სისტემები (cAMP, cGMP, ინოზიტოლ-3-ფოსფატი, დიაცილგლიცეროლი, Ca++ იონები).
პოსტსინაფსურ მემბრანაზე ორი ტიპის რეცეპტორებია – იონოტროპული და მეტაბოტროპული.
იონოტროპული
Როდესაც იონოტროპული რეცეპტორიმგრძნობიარე მოლეკულა შეიცავს არა მხოლოდ შუამავლის შეკავშირების აქტიურ ადგილს, არამედ იონურ არხსაც. „პირველადი მედიატორის“ (მედიატორის) ზემოქმედება რეცეპტორზე იწვევს არხის სწრაფ გახსნას და პოსტსინაფსური პოტენციალის განვითარებას.
მეტაბოტროპული
როდესაც მედიატორი მიმაგრებულია და მეტაბოტროპული რეცეპტორი აღგზნებულია, უჯრედშიდა მეტაბოლიზმი იცვლება, ე.ი. ბიოქიმიური რეაქციების მიმდინარეობა
მემბრანის შიგნიდან ამგვარ რეცეპტორზე მიმაგრებულია სხვა ცილები, რომლებიც ასრულებენ ფერმენტულ და ნაწილობრივ გადამცემს („შუამავალ“) ფუნქციებს (ნახ.). შუამავალ პროტეინებს მოიხსენიებენ როგორც G პროტეინებს. აღგზნებული რეცეპტორის გავლენით G- პროტეინი მოქმედებს ცილა-ფერმენტზე, ჩვეულებრივ აყენებს მას "მუშა" მდგომარეობაში. შედეგად, წარმოიქმნება ქიმიური რეაქცია: წინამორბედი მოლეკულა იქცევა სიგნალის მოლეკულად - მეორე მესინჯერად.

ბრინჯი. მეტაბოტროპული რეცეპტორის სტრუქტურისა და ფუნქციონირების სქემა: 1 - შუამავალი; 2 - რეცეპტორი; 3 - იონური არხი; 4 - მეორადი შუამავალი; 5 - ფერმენტი; 6 - G- პროტეინი; → - სიგნალის გადაცემის მიმართულება
მეორადი შუამავლები არის პატარა, სწრაფად მოძრავი მოლეკულები ან იონები, რომლებიც გადასცემენ სიგნალს უჯრედის შიგნით. ამით ისინი განსხვავდებიან "პირველადი შუამავლებისგან" - შუამავლებისგან და ჰორმონებისგან, რომლებიც გადასცემენ ინფორმაციას უჯრედიდან უჯრედში.
ყველაზე ცნობილი მეორე მესინჯერი არის cAMP (ციკლური ადენოზინმონოფოსფორის მჟავა), რომელიც წარმოიქმნება ATP-დან ფერმენტ ადენილატ ციკლაზას მიერ. ის ჰგავს cGMP-ს (გუანოზინის მონოფოსფორის მჟავას). სხვა მნიშვნელოვანი მეორადი მესინჯერებია ინოზიტოლ ტრიფოსფატი და დიაცილგლიცეროლი, რომლებიც წარმოიქმნება უჯრედის მემბრანის კომპონენტებიდან ფერმენტ ფოსფოლიპაზა C-ის მოქმედებით. Ca 2+-ის როლი, რომელიც შედის უჯრედში გარედან იონური არხებით ან გამოიყოფა სპეციალური საცავიდან. უჯრედის შიგნით არსებული ადგილები (კალციუმის „დეპო“) ძალზე მნიშვნელოვანია. ბოლო დროს დიდი ყურადღება დაეთმო მეორე მესინჯერს NO (აზოტის ოქსიდი), რომელსაც შეუძლია სიგნალის გადაცემა არა მხოლოდ უჯრედის შიგნით, არამედ უჯრედებს შორის, ადვილად გადაკვეთს მემბრანას, მათ შორის პოსტსინაფსური ნეირონიდან პრესინაფსამდე.
ქიმიური სიგნალის ჩატარების საბოლოო ნაბიჯი არის მეორე მესინჯერის მოქმედება ქიმიოსენსიტიურ იონურ არხზე. ეს ეფექტი ხდება პირდაპირ ან დამატებითი შუალედური ბმულების (ფერმენტების) მეშვეობით. ნებისმიერ შემთხვევაში, იონური არხი იხსნება და ვითარდება EPSP ან IPSP. მათი პირველი ფაზის ხანგრძლივობა და ამპლიტუდა განისაზღვრება მეორე მესინჯერის რაოდენობით, რაც დამოკიდებულია გამოთავისუფლებული შუამავლის რაოდენობაზე და რეცეპტორთან მისი ურთიერთქმედების ხანგრძლივობაზე.
ამრიგად, მეტაბოტროპული რეცეპტორების მიერ გამოყენებული ნერვული სტიმულის გადაცემის მექანიზმი მოიცავს რამდენიმე თანმიმდევრულ ეტაპს. თითოეულ მათგანზე შესაძლებელია სიგნალის რეგულაცია (დასუსტება ან გაძლიერება), რაც პოსტსინაფსური უჯრედის რეაქციას უფრო მოქნილს და არსებულ პირობებს ადაპტირებს. თუმცა ეს ასევე ანელებს ინფორმაციის გადაცემის პროცესს.
cAMP სისტემა

ფოსფოლიპაზა C

იმ პროცესების გასაგებად, რომლებიც უზრუნველყოფენ ცოცხალ უჯრედებში ელექტრული პოტენციალის არსებობას, პირველ რიგში აუცილებელია უჯრედის მემბრანის სტრუქტურისა და მისი თვისებების გაგება.
ამჟამად ყველაზე დიდი აღიარებით სარგებლობს ს.სინგერის და გ.ნიკოლსონის მიერ 1972 წელს შემოთავაზებული მემბრანის სითხე-მოზაიკური მოდელი.მემბრანის საფუძველია ფოსფოლიპიდების ორმაგი ფენა (ორმაგი შრე), მოლეკულის ჰიდროფობიური ფრაგმენტები. რომელთაგან ჩაძირულია მემბრანის სისქეში და პოლარული ჰიდროფილური ჯგუფები ორიენტირებულია გარეთ, იმ. მიმდებარე წყლის გარემოში (ნახ. 2.9).
მემბრანის ცილები ლოკალიზებულია მემბრანის ზედაპირზე ან შეიძლება ჩანერგილი იყოს ჰიდროფობიურ ზონაში სხვადასხვა სიღრმეზე. ზოგიერთი ცილა შეაღწევს მემბრანაში და მეშვეობით, და იმავე ცილის სხვადასხვა ჰიდროფილური ჯგუფები გვხვდება უჯრედის მემბრანის ორივე მხარეს. პლაზმურ მემბრანაში ნაპოვნი ცილები ძალიან მნიშვნელოვან როლს ასრულებენ: ისინი მონაწილეობენ იონური არხების ფორმირებაში, ასრულებენ მემბრანული ტუმბოების და სხვადასხვა ნივთიერების მატარებლების როლს და ასევე შეუძლიათ რეცეპტორული ფუნქციის შესრულება.
უჯრედის მემბრანის ძირითადი ფუნქციები: ბარიერი, სატრანსპორტო, მარეგულირებელი, კატალიზური.
ბარიერის ფუნქციაა მემბრანის მეშვეობით წყალში ხსნადი ნაერთების დიფუზიის შეზღუდვა, რაც აუცილებელია უჯრედების უცხო, ტოქსიკური ნივთიერებებისგან დასაცავად და უჯრედებში სხვადასხვა ნივთიერებების შედარებით მუდმივი შემცველობის შესანარჩუნებლად. ასე რომ, უჯრედის მემბრანას შეუძლია შეანელოს სხვადასხვა ნივთიერების დიფუზია 100,000-10,000,000-ჯერ.
ბრინჯი. 2.9.
ნაჩვენებია გლობულური ინტეგრალური ცილები, რომლებიც ჩაშენებულია ლიპიდურ ორ შრეში. ზოგიერთი ცილა არის იონური არხი, სხვები (გლიკოპროტეინები) შეიცავს ოლიგოსაქარიდულ გვერდით ჯაჭვებს, რომლებიც მონაწილეობენ უჯრედების ერთმანეთის ამოცნობაში და უჯრედშორის ქსოვილში. ქოლესტერინის მოლეკულები მჭიდროდ არის მიმდებარე ფოსფოლიპიდების თავებთან და აფიქსირებს "კუდების" მიმდებარე უბნებს. ფოსფოლიპიდური მოლეკულის კუდების შიდა რეგიონები არ არის შეზღუდული მათი მოძრაობაში და პასუხისმგებელია მემბრანის სითხეზე (Bretscher, 1985).
მემბრანაში არის არხები, რომლებითაც იონები შეაღწევენ. არხები არის პოტენციალზე დამოკიდებული და პოტენციური დამოუკიდებელი. პოტენციური არხებიიხსნება, როდესაც იცვლება პოტენციური განსხვავება და პოტენციურად დამოუკიდებელი(ჰორმონით რეგულირებადი) იხსნება რეცეპტორების ნივთიერებებთან ურთიერთქმედების დროს. არხების გახსნა ან დახურვა შესაძლებელია კარიბჭის წყალობით. მემბრანაში ჩაშენებულია ორი ტიპის კარიბჭე: გააქტიურება(არხის სიღრმეში) და ინაქტივაცია(არხის ზედაპირზე). კარიბჭე შეიძლება იყოს სამი მდგომარეობიდან ერთ-ერთში:
- ღია მდგომარეობა (ორივე ტიპის კარიბჭე ღიაა);
- დახურული მდგომარეობა (გააქტიურების კარიბჭე დახურულია);
- ინაქტივაციის მდგომარეობა (ინაქტივაციის კარიბჭე დახურულია). მემბრანების კიდევ ერთი დამახასიათებელი თვისებაა არაორგანული იონების, საკვები ნივთიერებებისა და სხვადასხვა მეტაბოლური პროდუქტების შერჩევითი გადაცემის უნარი. არსებობს ნივთიერებების პასიური და აქტიური გადაცემის (ტრანსპორტის) სისტემები. Პასიურიტრანსპორტირება ხორციელდება იონური არხებით გადამზიდავი ცილების დახმარებით ან მის გარეშე და მისი მამოძრავებელი ძალა არის იონების ელექტროქიმიური პოტენციალის განსხვავება უჯრედშიდა და უჯრედგარე სივრცეს შორის. იონური არხების სელექციურობა განისაზღვრება მისი გეომეტრიული პარამეტრებით და არხის კედლებისა და პირის მოპირკეთებული ჯგუფების ქიმიური ბუნებით.
ამჟამად ყველაზე კარგად შესწავლილი არხები Na + , K + , Ca 2+ იონების, აგრეთვე წყლის (ე.წ. აკვაპორინების) შერჩევითი გამტარიანობის მქონე არხებია. იონური არხების დიამეტრი, სხვადასხვა კვლევების მიხედვით, არის 0,5-0,7 ნმ. არხების გამტარუნარიანობა შეიძლება შეიცვალოს; 10 7 - 10 8 იონი წამში შეიძლება გაიაროს ერთ იონურ არხზე.
აქტიურიტრანსპორტი ხდება ენერგიის ხარჯვით და ხორციელდება ე.წ. იონური ტუმბოებით. იონის ტუმბოები არის მოლეკულური ცილის სტრუქტურები, რომლებიც ჩაშენებულია მემბრანაში და ახორციელებენ იონების გადატანას უფრო მაღალი ელექტროქიმიური პოტენციალისკენ.
ტუმბოების მუშაობა ხორციელდება ATP ჰიდროლიზის ენერგიის გამო. Na + /K + - ATPase, Ca 2+ - ATPase, H + - ATPase, H + /K + - ATPase, Mg 2+ - ATPase, რომლებიც უზრუნველყოფენ Na +, K +, Ca იონების მოძრაობას შესაბამისად. კარგად შესწავლილი 2+, H+, Mg 2+ იზოლირებული ან კონიუგირებული (Na + და K +; H + და K +). აქტიური ტრანსპორტის მოლეკულური მექანიზმი ბოლომდე არ არის განმარტებული.
ცხრილი ნომერი 2
კითხვა 1 (8)
უჯრედის მემბრანა(ან ციტოლემა, ან პლაზმალემა, ან პლაზმური მემბრანა) გამოყოფს ნებისმიერი უჯრედის შიგთავსს გარე გარემოდან, რაც უზრუნველყოფს მის მთლიანობას; არეგულირებს უჯრედსა და გარემოს შორის გაცვლას; უჯრედშიდა გარსები უჯრედს ყოფს სპეციალიზებულ დახურულ ნაწილებად - კუპეებად ან ორგანელებად, რომლებშიც შენარჩუნებულია გარკვეული გარემო პირობები.
უჯრედის ან პლაზმური მემბრანის ფუნქციები
მემბრანა უზრუნველყოფს:
1) უჯრედის სპეციფიკური ფუნქციების შესასრულებლად საჭირო მოლეკულებისა და იონების შერჩევითი შეღწევა უჯრედში და მის გარეთ;
2) იონების შერჩევითი ტრანსპორტირება მემბრანის გასწვრივ, ტრანსმემბრანული ელექტრული პოტენციალის სხვაობის შენარჩუნებით;
3) უჯრედშორისი კონტაქტების სპეციფიკა.
მემბრანაში მრავალი რეცეპტორის არსებობის გამო, რომლებიც აღიქვამენ ქიმიურ სიგნალებს - ჰორმონებს, შუამავლებს და სხვა ბიოლოგიურად აქტიურ ნივთიერებებს, მას შეუძლია შეცვალოს უჯრედის მეტაბოლური აქტივობა. მემბრანები უზრუნველყოფენ იმუნური გამოვლინებების სპეციფიკას მათზე ანტიგენების არსებობის გამო - სტრუქტურები, რომლებიც იწვევენ ანტისხეულების წარმოქმნას, რომლებსაც შეუძლიათ კონკრეტულად დაუკავშირდნენ ამ ანტიგენებს.
უჯრედის ბირთვი და ორგანელები ასევე გამოყოფილია ციტოპლაზმიდან მემბრანებით, რომლებიც ხელს უშლიან წყლისა და მასში გახსნილი ნივთიერებების თავისუფალ მოძრაობას ციტოპლაზმიდან მათკენ და პირიქით. ეს ქმნის პირობებს ბიოქიმიური პროცესების გამოყოფისთვის, რომლებიც ხდება უჯრედის შიგნით სხვადასხვა განყოფილებებში (კუპეში).
უჯრედის მემბრანის სტრუქტურა
უჯრედის მემბრანა- ელასტიური სტრუქტურა, სისქე 7-დან 11 ნმ-მდე (ნახ. 1.1). იგი ძირითადად შედგება ლიპიდებისა და ცილებისგან. ყველა ლიპიდების 40-დან 90%-მდე არის ფოსფოლიპიდები - ფოსფატიდილქოლინი, ფოსფატიდილათანოლამინი, ფოსფატიდილსერინი, სფინგომიელინი და ფოსფატიდილინოზიტოლი. მემბრანის მნიშვნელოვანი კომპონენტია გლიკოლიპიდები, რომლებიც წარმოდგენილია ცერებრიდები, სულფატიდები, განგლიოზიდები და ქოლესტერინი.
უჯრედის მემბრანის ძირითადი სტრუქტურაარის ფოსფოლიპიდური მოლეკულების ორმაგი ფენა. ჰიდროფობიური ურთიერთქმედების გამო, ლიპიდური მოლეკულების ნახშირწყლების ჯაჭვები ერთმანეთთან ახლოს არის გაფართოებულ მდგომარეობაში. ორივე ფენის ფოსფოლიპიდური მოლეკულების ჯგუფები ურთიერთქმედებენ ლიპიდურ მემბრანაში ჩაძირულ ცილის მოლეკულებთან. იმის გამო, რომ ორფენიანი ლიპიდური კომპონენტების უმეტესობა თხევად მდგომარეობაშია, მემბრანას აქვს მობილურობა და ტალღოვანი. მისი სექციები, ისევე როგორც ლიპიდურ ორშრეში ჩაძირული ცილები, შერევა ერთი ნაწილიდან მეორეში. უჯრედის მემბრანების მობილურობა (სთხევადობა) ხელს უწყობს ნივთიერებების მემბრანის მეშვეობით ტრანსპორტირებას.
უჯრედის მემბრანის ცილებიწარმოდგენილია ძირითადად გლიკოპროტეინებით.
გამოარჩევენ
ინტეგრალური ცილებიგარსის მთელ სისქეში შეღწევა და
პერიფერიული ცილებიმიმაგრებულია მხოლოდ მემბრანის ზედაპირზე, ძირითადად მის შიდა ნაწილზე.
პერიფერიული ცილებითითქმის ყველა ფუნქციონირებს როგორც ფერმენტები (აცეტილქოლინესტერაზა, მჟავა და ტუტე ფოსფატაზები და ა.შ.). მაგრამ ზოგიერთი ფერმენტი ასევე წარმოდგენილია ინტეგრალური ცილებით - ATPase.
ინტეგრალური ცილებიუზრუნველყოფს იონების შერჩევით გაცვლას მემბრანული არხებით უჯრედგარე და უჯრედშორის სითხეს შორის და ასევე მოქმედებს როგორც პროტეინები - დიდი მოლეკულების მატარებლები.
მემბრანული რეცეპტორები და ანტიგენები შეიძლება წარმოდგენილი იყოს როგორც ინტეგრალური, ისე პერიფერიული ცილებით.
ციტოპლაზმური მხრიდან მემბრანის მიმდებარე ცილები მიეკუთვნება უჯრედის ციტოჩონჩხი. მათ შეუძლიათ მემბრანის ცილებთან მიმაგრება.
Ისე, ცილის ზოლები 3ერითროციტების მემბრანების (ზოლის ნომერი ცილის ელექტროფორეზის დროს) გაერთიანებულია ანსამბლში სხვა ციტოჩონჩხის მოლეკულებთან - სპექტრინთან დაბალი მოლეკულური წონის პროტეინის ანკირინის მეშვეობით.
სპექტრინიარის ციტოჩონჩხის მთავარი ცილა, რომელიც წარმოადგენს ორგანზომილებიან ქსელს, რომელსაც აქტინი ერთვის.
აქტინიქმნის მიკროფილამენტებს, რომლებიც წარმოადგენს ციტოჩონჩხის კონტრაქტურ აპარატს.
ციტოჩონჩხისაშუალებას აძლევს უჯრედს გამოავლინოს მოქნილი ელასტიური თვისებები, უზრუნველყოფს მემბრანის დამატებით სიმტკიცეს.
ინტეგრალური ცილების უმეტესობა გლიკოპროტეინებია. მათი ნახშირწყლების ნაწილი უჯრედის მემბრანიდან გარედან გამოდის. ბევრ გლიკოპროტეინს აქვს დიდი უარყოფითი მუხტი სიალიუმის მჟავის მნიშვნელოვანი შემცველობის გამო (მაგალითად, გლიკოფორინის მოლეკულა). ეს უზრუნველყოფს უჯრედების უმეტესობის ზედაპირს უარყოფითი მუხტით, რაც ხელს უწყობს სხვა უარყოფითად დამუხტული ობიექტების მოგერიებას. გლიკოპროტეინების ნახშირწყლოვანი პროტრუზია ატარებს სისხლის ჯგუფის ანტიგენებს, უჯრედის სხვა ანტიგენურ დეტერმინანტებს და მოქმედებს როგორც ჰორმონების დამაკავშირებელი რეცეპტორები. გლიკოპროტეინები ქმნიან წებოვან მოლეკულებს, რომლებიც იწვევენ უჯრედების ერთმანეთთან მიმაგრებას, ე.ი. დახურეთ უჯრედშორისი კონტაქტები.
ბიოლოგიური გარსები.
ტერმინი „მემბრანა“ (ლათ. membrana - კანი, ფილმი) გამოყენება დაიწყო 100 წელზე მეტი ხნის წინ უჯრედის საზღვრის აღსანიშნავად, რომელიც ემსახურება, ერთი მხრივ, როგორც ბარიერს უჯრედის შიგთავსსა და გარე გარემოს შორის. , და მეორეს მხრივ, როგორც ნახევრად გამტარი დანაყოფი, რომლითაც წყალი და ზოგიერთი ნივთიერება შეიძლება გაიაროს. თუმცა, მემბრანის ფუნქციები არ არის ამოწურული,ვინაიდან ბიოლოგიური მემბრანები ქმნიან უჯრედის სტრუქტურული ორგანიზაციის საფუძველს.
მემბრანის სტრუქტურა. ამ მოდელის მიხედვით, მთავარი მემბრანა არის ლიპიდური ორფენი, რომელშიც მოლეკულების ჰიდროფობიური კუდები შიგნითაა მოქცეული, ხოლო ჰიდროფილური თავები – გარეთ. ლიპიდები წარმოდგენილია ფოსფოლიპიდებით - გლიცეროლის ან სფინგოზინის წარმოებულები. ცილები მიმაგრებულია ლიპიდურ ფენაზე. ინტეგრალური (ტრანსმემბრანული) ცილები მემბრანაში აღწევენ და მყარად არიან დაკავშირებული მასთან; პერიფერიული არ აღწევენ და ნაკლებად მყარად არიან დაკავშირებული მემბრანასთან. მემბრანის ცილების ფუნქციები: მემბრანების სტრუქტურის შენარჩუნება, გარემოდან სიგნალების მიღება და გარდაქმნა. გარემო, გარკვეული ნივთიერებების ტრანსპორტირება, მემბრანებზე მიმდინარე რეაქციების კატალიზება. მემბრანის სისქე 6-დან 10 ნმ-მდეა.
მემბრანის თვისებები:
1. სითხე. მემბრანა არ არის ხისტი სტრუქტურა; მისი ცილების და ლიპიდების უმეტესობას შეუძლია მემბრანის სიბრტყეში გადაადგილება.
2. ასიმეტრია. როგორც ცილების, ასევე ლიპიდების გარე და შიდა ფენების შემადგენლობა განსხვავებულია. გარდა ამისა, ცხოველური უჯრედების პლაზმურ მემბრანებს აქვთ გლიკოპროტეინების ფენა გარედან (გლიკოკალიქსი, რომელიც ასრულებს სიგნალის და რეცეპტორების ფუნქციებს და ასევე მნიშვნელოვანია უჯრედების ქსოვილებში გაერთიანებისთვის).
3. პოლარობა. მემბრანის გარედან დადებით მუხტს ატარებს, ხოლო შიგნიდან უარყოფით მუხტს.
4. შერჩევითი გამტარიანობა. ცოცხალი უჯრედების მემბრანები წყლის გარდა გადის მხოლოდ გარკვეულ მოლეკულებს და გახსნილი ნივთიერებების იონებს. (ტერმინის „ნახევრად გამტარიანობა“ უჯრედის მემბრანებთან მიმართებაში მთლად სწორი არ არის, რადგან ეს ცნება გულისხმობს, რომ მემბრანა გადის მხოლოდ გამხსნელს. მოლეკულები, ყველა მოლეკულის და ხსნარის იონების შენარჩუნებისას.)
გარე უჯრედის მემბრანა (პლაზმალემა) არის ულტრამიკროსკოპული ფილმი 7,5 ნმ სისქით, რომელიც შედგება ცილების, ფოსფოლიპიდების და წყლისგან. ელასტიური ფილმი, კარგად დასველებული წყლით და სწრაფად აღადგენს მთლიანობას დაზიანების შემდეგ. მას აქვს უნივერსალური სტრუქტურა, რომელიც დამახასიათებელია ყველა ბიოლოგიური მემბრანისთვის. ამ მემბრანის სასაზღვრო პოზიცია, მისი მონაწილეობა სელექციური გამტარიანობის, პინოციტოზის, ფაგოციტოზის, ექსკრეციული პროდუქტების გამოყოფისა და სინთეზის პროცესებში, მეზობელ უჯრედებთან ერთად და უჯრედის დაზიანებისგან დაცვას, მის როლს უაღრესად მნიშვნელოვანს ხდის. მემბრანის გარეთ ცხოველური უჯრედები ზოგჯერ დაფარულია თხელი ფენით, რომელიც შედგება პოლისაქარიდებისა და ცილებისგან - გლიკოკალიქსისგან. უჯრედის მემბრანის გარეთ მცენარეულ უჯრედებს აქვთ ძლიერი უჯრედის კედელი, რომელიც ქმნის გარე საყრდენს და ინარჩუნებს უჯრედის ფორმას. იგი შედგება ბოჭკოვანი (ცელულოზა) წყალში უხსნადი პოლისაქარიდისგან.
უჯრედის მემბრანა
უჯრედის მემბრანის გამოსახულება. პატარა ლურჯი და თეთრი ბურთულები შეესაბამება ფოსფოლიპიდების ჰიდროფობიურ „თავებს“, მათზე დამაგრებული ხაზები კი ჰიდროფილურ „კუდებს“. ნახატზე ნაჩვენებია მხოლოდ მემბრანის განუყოფელი ცილები (წითელი გლობულები და ყვითელი ხვეულები). მემბრანის შიგნით ყვითელი ოვალური წერტილები - ქოლესტერინის მოლეკულები გარსის გარეთა მძივების ყვითელ-მწვანე ჯაჭვები - ოლიგოსაქარიდული ჯაჭვები, რომლებიც ქმნიან გლიკოკალიქსს
ბიოლოგიურ მემბრანაში ასევე შედის სხვადასხვა ცილები: ინტეგრალური (მემბრანაში შეღწევა), ნახევრად ინტეგრალური (ერთ ბოლოში ჩაეფლო გარე ან შიდა ლიპიდურ შრეში), ზედაპირული (მდებარეობს მემბრანის გარე ან შიდა მხარეების მიმდებარედ). ზოგიერთი ცილა არის უჯრედის მემბრანის შეხების წერტილები ციტოჩონჩხთან უჯრედის შიგნით, ხოლო უჯრედის კედელთან (ასეთის არსებობის შემთხვევაში) გარეთ. ზოგიერთი ინტეგრალური ცილა ფუნქციონირებს როგორც იონური არხები, სხვადასხვა გადამტანები და რეცეპტორები.
ფუნქციები
- ბარიერი - უზრუნველყოფს გარემოსთან რეგულირებულ, შერჩევით, პასიურ და აქტიურ მეტაბოლიზმს. მაგალითად, პეროქსიზომის მემბრანა იცავს ციტოპლაზმას უჯრედისთვის საშიში პეროქსიდებისგან. შერჩევითი გამტარიანობა ნიშნავს, რომ მემბრანის გამტარიანობა სხვადასხვა ატომების ან მოლეკულების მიმართ დამოკიდებულია მათ ზომაზე, ელექტრულ მუხტზე და ქიმიურ თვისებებზე. შერჩევითი გამტარიანობა უზრუნველყოფს უჯრედის და უჯრედული ნაწილების გარემოდან გამოყოფას და მათ საჭირო ნივთიერებებით მიწოდებას.
- ტრანსპორტი - მემბრანის მეშვეობით ხდება ნივთიერებების ტრანსპორტირება უჯრედში და უჯრედის გარეთ. მემბრანების მეშვეობით ტრანსპორტი უზრუნველყოფს: საკვები ნივთიერებების მიწოდებას, მეტაბოლიზმის საბოლოო პროდუქტების მოცილებას, სხვადასხვა ნივთიერებების სეკრეციას, იონური გრადიენტების შექმნას, უჯრედში იონების ოპტიმალური კონცენტრაციის შენარჩუნებას, რაც აუცილებელია ფუნქციონირებისთვის. ფიჭური ფერმენტები.
ნაწილაკები, რომლებიც რაიმე მიზეზით ვერ კვეთენ ფოსფოლიპიდურ ორ შრეს (მაგალითად, ჰიდროფილური თვისებების გამო, რადგან მემბრანა შიგნით არის ჰიდროფობიური და არ აძლევს ჰიდროფილურ ნივთიერებებს გავლის საშუალებას, ან მათი დიდი ზომის გამო), მაგრამ აუცილებელია უჯრედისთვის. , შეუძლია შეაღწიოს მემბრანაში სპეციალური გადამზიდავი ცილების (ტრანსპორტირების) და არხის ცილების ან ენდოციტოზის მეშვეობით.
პასიურ ტრანსპორტში ნივთიერებები კვეთენ ლიპიდურ ორ შრეს ენერგიის დახარჯვის გარეშე კონცენტრაციის გრადიენტის გასწვრივ დიფუზიის გზით. ამ მექანიზმის ვარიანტია გაადვილებული დიფუზია, რომლის დროსაც სპეციფიკური მოლეკულა ეხმარება ნივთიერებას მემბრანის გავლით. ამ მოლეკულას შეიძლება ჰქონდეს არხი, რომელიც მხოლოდ ერთი ტიპის ნივთიერების გავლის საშუალებას აძლევს.
აქტიური ტრანსპორტი მოითხოვს ენერგიას, რადგან ეს ხდება კონცენტრაციის გრადიენტის საწინააღმდეგოდ. მემბრანაზე არის სპეციალური ტუმბოს პროტეინები, მათ შორის ატფ-აზა, რომელიც აქტიურად ტუმბოს კალიუმის იონებს (K +) უჯრედში და გამოაქვს ნატრიუმის იონები (Na +). - მატრიცა - უზრუნველყოფს მემბრანის ცილების გარკვეულ ფარდობით პოზიციას და ორიენტაციას, მათ ოპტიმალურ ურთიერთქმედებას.
- მექანიკური - უზრუნველყოფს უჯრედის ავტონომიას, მის უჯრედშიდა სტრუქტურებს, ასევე სხვა უჯრედებთან (ქსოვილებში) კავშირს. უჯრედის კედლები მნიშვნელოვან როლს ასრულებს მექანიკური ფუნქციის უზრუნველსაყოფად, ხოლო ცხოველებში - უჯრედშორისი ნივთიერება.
- ენერგია - ქლოროპლასტებში ფოტოსინთეზის და მიტოქონდრიებში უჯრედული სუნთქვის დროს მათ მემბრანებში მოქმედებს ენერგიის გადაცემის სისტემები, რომელშიც ცილებიც მონაწილეობენ;
- რეცეპტორი - მემბრანაში მდებარე ზოგიერთი ცილა არის რეცეპტორები (მოლეკულები, რომლებითაც უჯრედი აღიქვამს გარკვეულ სიგნალებს).
მაგალითად, სისხლში მოცირკულირე ჰორმონები მოქმედებენ მხოლოდ სამიზნე უჯრედებზე, რომლებსაც აქვთ ამ ჰორმონების შესაბამისი რეცეპტორები. ნეიროტრანსმიტერები (ქიმიკატები, რომლებიც ატარებენ ნერვულ იმპულსებს) ასევე უკავშირდებიან სამიზნე უჯრედების სპეციფიკურ რეცეპტორულ ცილებს. - ფერმენტული - მემბრანის ცილები ხშირად ფერმენტებია. მაგალითად, ნაწლავის ეპითელური უჯრედების პლაზმური მემბრანები შეიცავს საჭმლის მომნელებელ ფერმენტებს.
- ბიოპოტენციალების გენერირებისა და გატარების განხორციელება.
მემბრანის დახმარებით უჯრედში იონების მუდმივი კონცენტრაცია შენარჩუნებულია: K + იონის კონცენტრაცია უჯრედის შიგნით გაცილებით მაღალია, ვიდრე გარეთ, ხოლო Na + კონცენტრაცია გაცილებით დაბალია, რაც ძალიან მნიშვნელოვანია, ვინაიდან ეს ინარჩუნებს პოტენციურ განსხვავებას მემბრანის გასწვრივ და წარმოქმნის ნერვულ იმპულსს. - უჯრედის მარკირება – მემბრანაზე არის ანტიგენები, რომლებიც მოქმედებენ როგორც მარკერები – „ეტიკეტები“, რომლებიც უჯრედის იდენტიფიცირების საშუალებას იძლევა. ეს არის გლიკოპროტეინები (ანუ ცილები მათზე მიმაგრებული განშტოებული ოლიგოსაქარიდული გვერდითი ჯაჭვებით), რომლებიც ასრულებენ „ანტენების“ როლს. გვერდითი ჯაჭვის უამრავი კონფიგურაციის გამო, შესაძლებელია თითოეული უჯრედის ტიპისთვის კონკრეტული მარკერის გაკეთება. მარკერების დახმარებით უჯრედებს შეუძლიათ სხვა უჯრედების ამოცნობა და მათთან ერთად მოქმედება, მაგალითად, ორგანოებისა და ქსოვილების ფორმირებისას. ის ასევე საშუალებას აძლევს იმუნურ სისტემას ამოიცნოს უცხო ანტიგენები.
ბიომემბრანების სტრუქტურა და შემადგენლობა
მემბრანები შედგება ლიპიდების სამი კლასისგან: ფოსფოლიპიდები, გლიკოლიპიდები და ქოლესტერინი. ფოსფოლიპიდები და გლიკოლიპიდები (ლიპიდები მათზე მიმაგრებული ნახშირწყლებით) შედგება ორი გრძელი ჰიდროფობიური ნახშირწყალბადის „კუდისგან“, რომლებიც დაკავშირებულია დამუხტულ ჰიდროფილურ „თავთან“. ქოლესტერინი ამაგრებს მემბრანას, იკავებს თავისუფალ ადგილს ჰიდროფობიურ ლიპიდურ კუდებს შორის და ხელს უშლის მათ დახრილობას. ამიტომ მემბრანები დაბალი ქოლესტერინის შემცველობით უფრო მოქნილია, ხოლო მაღალი ქოლესტერინის შემცველობა უფრო ხისტი და მტვრევადი. ქოლესტერინი ასევე ემსახურება როგორც "საჩერებელი", რომელიც ხელს უშლის პოლარული მოლეკულების მოძრაობას უჯრედიდან და შიგნით. მემბრანის მნიშვნელოვანი ნაწილი შედგება მასში შემავალი ცილებისგან და პასუხისმგებელია მემბრანის სხვადასხვა თვისებებზე. მათი შემადგენლობა და ორიენტაცია სხვადასხვა მემბრანაში განსხვავდება.
უჯრედის მემბრანები ხშირად ასიმეტრიულია, ანუ ფენები განსხვავდება ლიპიდური შემადგენლობით, ცალკეული მოლეკულის გადასვლა ერთი ფენიდან მეორეზე (ე.წ. ფლიპ ფლოპი) ძნელია.
მემბრანული ორგანელები
ეს არის ციტოპლაზმის დახურული ცალკეული ან ურთიერთდაკავშირებული სექციები, რომლებიც გამოყოფილია ჰიალოპლაზმისგან მემბრანებით. ერთმემბრანიან ორგანელებს მიეკუთვნება ენდოპლაზმური ბადე, გოლჯის აპარატი, ლიზოსომები, ვაკუოლები, პეროქსიზომები; ორმემბრანამდე - ბირთვი, მიტოქონდრია, პლასტიდები. სხვადასხვა ორგანოილების მემბრანების სტრუქტურა განსხვავდება ლიპიდებისა და მემბრანის ცილების შემადგენლობით.
შერჩევითი გამტარიანობა
უჯრედის მემბრანებს აქვთ შერჩევითი გამტარიანობა: გლუკოზა, ამინომჟავები, ცხიმოვანი მჟავები, გლიცეროლი და იონები ნელ-ნელა დიფუზირდება მათში და თავად მემბრანები გარკვეულწილად აქტიურად არეგულირებენ ამ პროცესს - ზოგი ნივთიერება გადის, ზოგი კი არა. არსებობს უჯრედში ნივთიერებების შეყვანის ან უჯრედიდან გარედან მათი მოცილების ოთხი ძირითადი მექანიზმი: დიფუზია, ოსმოზი, აქტიური ტრანსპორტი და ეგზო- ან ენდოციტოზი. პირველი ორი პროცესი პასიური ხასიათისაა, ანუ არ საჭიროებს ენერგიას; ბოლო ორი არის აქტიური პროცესები, რომლებიც დაკავშირებულია ენერგიის მოხმარებასთან.
პასიური ტრანსპორტის დროს მემბრანის შერჩევითი გამტარიანობა განპირობებულია სპეციალური არხებით - ინტეგრალური ცილებით. ისინი შეაღწევენ მემბრანაში და მეშვეობით, ქმნიან ერთგვარ გადასასვლელს. ელემენტებს K, Na და Cl აქვთ საკუთარი არხები. კონცენტრაციის გრადიენტთან დაკავშირებით, ამ ელემენტების მოლეკულები მოძრაობენ უჯრედში და გარეთ. გაღიზიანების დროს იხსნება ნატრიუმის იონური არხები და ხდება ნატრიუმის იონების მკვეთრი შემოდინება უჯრედში. ეს იწვევს მემბრანის პოტენციალის დისბალანსს. ამის შემდეგ მემბრანის პოტენციალი აღდგება. კალიუმის არხები ყოველთვის ღიაა, მათი მეშვეობით კალიუმის იონები ნელ-ნელა შედიან უჯრედში.
იხილეთ ასევე
ლიტერატურა
- ანტონოვი ვ.ფ., სმირნოვა ე.ნ., შევჩენკო ე.ვ.ლიპიდური გარსები ფაზური გადასვლის დროს. - მ .: ნაუკა, 1994 წ.
- გენის რ.ბიომემბრანები. მოლეკულური სტრუქტურა და ფუნქციები: თარგმანი ინგლისურიდან. = ბიომემბრანები. მოლეკულური სტრუქტურა და ფუნქცია (რობერტ ბ. გენისი). - 1 გამოცემა. - M .: Mir, 1997. - ISBN 5-03-002419-0
- ივანოვი V.G., Berestovsky T.N.ბიოლოგიური მემბრანების ლიპიდური ორშერი. - მ .: ნაუკა, 1982 წ.
- რუბინ A.B.ბიოფიზიკა, სახელმძღვანელო 2 ტ. - მე-3 გამოცემა, შესწორებული და გაფართოებული. - მ .: მოსკოვის უნივერსიტეტის გამომცემლობა, 2004. - ISBN 5-211-06109-8
- ბრიუს ალბერტსი და სხვ.