Functions of the plasma membrane. Plasma membrane The structure and function of the plasma
The plasma membrane occupies a special position, since it limits the cell outside and is directly connected with the extracellular environment. It is about 10 nm thick and is the thickest cell membrane. The main components are proteins (over 60%), lipids (about 40%) and carbohydrates (about 1%). Like all other cell membranes, it is synthesized in the EPS channels.
Plasmalemma functions.
Transport.
The plasma membrane is semi-permeable, i.e. selectively different molecules pass through it at different speeds. There are two ways of transporting substances across the membrane: passive and active transport.
Passive transport. Passive transport or diffusion does not require energy input. Uncharged molecules diffuse along the concentration gradient, the transport of charged molecules depends on the concentration gradient of hydrogen protons and the transmembrane potential difference, which are combined into an electrochemical proton gradient. As a rule, the inner cytoplasmic surface of the membrane carries a negative charge, which facilitates the penetration of positively charged ions into the cell. There are two types of diffusion: simple and lightweight.
Simple diffusion is typical for small neutral molecules (H 2 O, CO 2, O 2), as well as for hydrophobic low molecular weight organic substances. These molecules can pass through the pores or channels of the membrane without any interaction with membrane proteins as long as the concentration gradient is maintained.
Facilitated diffusion is characteristic of hydrophilic molecules, which are transported across the membrane also along the concentration gradient, but with the help of special membrane carrier proteins according to the principle uniforms.
Facilitated diffusion is highly selective, since the carrier protein has a binding center complementary to the transported substance, and the transfer is accompanied by conformational changes in the protein. One of the possible mechanisms of facilitated diffusion is as follows: the transport protein (translocase) binds the substance, then approaches the opposite side of the membrane, releases this substance, assumes the initial conformation and is again ready to perform the transport function. Little is known about how the protein itself moves. Another possible transfer mechanism involves the involvement of several carrier proteins. In this case, the initially bound compound itself passes from one protein to another, sequentially binding with one or another protein until it is on the opposite side of the membrane.
Active transport. Such transport takes place when the transfer is carried out against the concentration gradient. It requires the expenditure of energy by the cell. Active transport serves for the accumulation of substances inside the cell. The energy source is often ATP. For active transport, in addition to the energy source, the participation of membrane proteins is necessary. One of the active transport systems in animal cells is responsible for the transport of Na and K + ions across the cell membrane. This system is called Na + - K * -pump. It is responsible for maintaining the composition of the intracellular environment, in which the concentration of K + ions is higher than that of Na * ions.
The concentration gradient of both ions is maintained by transferring K + to the inside of the cell, and Na + to the outside. Both transports occur against a concentration gradient. This distribution of ions determines the water content in cells, the excitability of nerve cells and muscle cells, and other properties of normal cells. Na + -K + -pump is a protein - transport ATPase. The molecule of this enzyme is an oligomer and permeates the membrane. During the full cycle of the pump operation, 3 Na + ions are transferred from the cell to the intercellular substance, and in the opposite direction - 2 K + ions, while the energy of the ATP molecule is used. There are transport systems for the transfer of calcium ions (Ca 2+ -ATPase), proton pumps (H + -ATPase), etc.
The active transfer of a substance through the membrane, carried out due to the energy of the concentration gradient of another substance, is called symptom... In this case, the transport ATPase has binding sites for both substances. Antiport is the movement of a substance against its concentration gradient. In this case, another substance moves in the opposite direction along its concentration gradient. Symptom and antiport (cotransport) can occur during absorption of amino acids from the intestine and reabsorption of glucose from primary urine, using the energy of the concentration gradient of Na + ions created by Na +, K + -ATPase.
Another 2 types of transport are endocytosis and exocytosis.
Endocytosis- capture of large particles by the cell. There are several ways of endocytosis: pinocytosis and phagocytosis. Usually under pinocytosis understand the capture of liquid colloidal particles by a cell, under phagocytosis- capture of corpuscles (denser and larger particles up to other cells). The mechanism of pino and phagocytosis is different.
In general, the entry into the cell of solid particles or liquid droplets from the outside is called heterophagy. This process is most widespread in protozoa, but it is very important in humans (as well as in other mammals). Heterophagy plays an essential role in the defense of the body (segmented neutrophils - granulocytes; macrophagocytes), remodeling of bone tissue (osteoclasts), thyroxine formation by thyroid follicles, reabsorption of protein and other macromolecules in the proximal nephron and other processes.
Pinocytosis.
In order for external molecules to enter the cell, they must first be bound by glycocalyx receptors (a set of molecules bound to the surface proteins of the membrane) (Fig.).
At the site of such binding, clathrin protein molecules are found under the plasmalemma. The plasmalemma, together with the molecules attached from the outside and the clathrin underlain by the cytoplasm, begins to invade. The protrusion becomes deeper, its edges approach and then close. As a result, a bubble is split off from the plasmalemma, which carries the captured molecules. Clathrin on its surface looks like an uneven border on electronic micrographs, therefore such bubbles are called bordered.
Clathrin prevents vesicles from adhering to intracellular membranes. Therefore, the bordered vesicles can be freely transported in the cell to precisely those parts of the cytoplasm where their contents should be used. This is how, in particular, steroid hormones are delivered to the nucleus. However, usually the bordered vesicles shed their border soon after cleavage from the plasmalemma. Clathrin is transferred to the plasmalemma and can again participate in endocytosis reactions.
At the cell surface in the cytoplasm there are more permanent vesicles - endosomes. Bordered vesicles release clathrin and merge with endosomes, increasing the volume and surface of the endosomes. Then the excess part of the endosomes is split off in the form of a new vesicle, in which there are no substances that have entered the cell, they remain in the endosome. The new vesicle is directed to the cell surface and fuses with the membrane. As a result, the decrease in the plasmalemma, which occurs during the splitting of the bordered vesicle, is restored, while its receptors also return to the plasmalemma.
Endosomes are immersed in the cytoplasm and fused with the membranes of the lysosome. The incoming substances inside such a secondary lysosome undergo various biochemical transformations. Upon completion of the process, the membrane of the lysosome can disintegrate into fragments, and the decay products and contents of the lysosome become available for intracellular metabolic reactions. For example, amino acids bind to tRNA and are delivered to the ribosomes, while glucose can enter the Golgi complex or the tubules of the agranular EPS.
Although endosomes do not have a clathrin border, not all of them fuse with lysosomes. Some of them are directed from one cell surface to another (if the cells form an epithelial layer). There, the membrane of the endosome merges with the plasmolemma and the contents are excreted. As a result, substances are transferred through the cell from one environment to another without changes. This process is called transcytosis... Protein molecules, in particular immunoglobulins, can also be transported by transcytosis.
Phagocytosis.
If a large particle has molecular groupings on its surface that can be recognized by cell receptors, it binds. By no means always are foreign particles themselves possessing such groups. However, getting into the body, they are surrounded by molecules of immunoglobulins (opsonins), which are always found in the blood and in the extracellular environment. Immunoglobulins are always recognized by phagocytic cells.
After the opsonins covering the foreign particle bind to the receptors of the phagocyte, its surface complex is activated. Actin microfilaments begin to interact with myosin, and the configuration of the cell surface changes. Outgrowths of the phagocyte cytoplasm are stretched around the particle. They cover the surface of the particle and combine over it. The outer leaves of the outgrowths merge, closing the cell surface.
Deep sheets of outgrowths form a membrane around the absorbed particle - it is formed phagosome. The phagosome merges with lysosomes, resulting in their complex - heterolysosome (heterosome, or phagolysosome). In it, lysis of the captured components of the particle takes place. Some of the lysis products are removed from the heterosome and utilized by the cell, while some may not be amenable to the action of lysosomal enzymes. These residues form residual bodies.
Potentially all cells have the ability to phagocytosis, but in the body, only a few specialize in this direction. These are neutrophilic leukocytes and macrophages.
Exocytosis.
This is the removal of substances from the cell. First, large-molecular compounds segregate in the Golgi complex in the form of transport bubbles. The latter, with the participation of microtubules, are directed to the cell surface. The membrane of the vesicle is embedded in the plasmalemma, and the contents of the vesicle are outside the cell (Fig.) The fusion of the vesicle with the plasmalemma can occur without any additional signals. This exocytosis is called constitutive. This is how most of the products of its own metabolism are removed from the cell. A number of cells, however, are designed to synthesize special compounds - secrets that are used in other parts of the body. In order for the transport vesicle with the secret to merge with the plasma membrane, external signals are required. Only then will the fusion take place and the secret will be released. This exocytosis is called regulated... Signaling molecules that promote secretion release are called liberins (releasing factors), and preventing elimination - statins.
Receptor functions.
They are mainly provided by glycoproteins located on the surface of the plasmalemma and capable of binding to their ligands. The ligand corresponds to its receptor like a key to a lock. Binding of the ligand to the receptor causes a change in the conformation of the polypeptide. With such a change in the transmembrane protein, communication is established between the extra- and intracellular environment.
Types of receptors.
Receptors associated with protein ion channels. They interact with a signaling molecule that temporarily opens or closes the channel for the passage of ions. (For example, the receptor for the neurotransmitter acetylcholine is a protein consisting of 5 subunits that form an ion channel. In the absence of acetylcholine, the channel is closed, and after attachment it opens and allows sodium ions to pass through).
Catalytic receptors. They consist of an extracellular part (the receptor itself) and an intracellular cytoplasmic part, which functions as an enzyme proline kinase (for example, growth hormone receptors).
Receptors associated with G-proteins. These are transmembrane proteins consisting of a receptor interacting with a ligand and a G-protein (guanosine triphosphate-linked regulatory protein), which transmits a signal to a membrane-bound enzyme (adenylate cyclase) or to an ion channel. As a result, cyclic AMP or calcium ions are activated. (This is how the adenylate cyclase system works. For example, in the liver cells there is a receptor for the hormone insulin. The supercellular part of the receptor binds to insulin. This causes the activation of the intracellular part - the enzyme adenylate cyclase. It synthesizes cyclic AMP from ATP, which regulates the rate of various intracellular processes, causing the activation or inhibition of those or other metabolic enzymes).
Receptors that perceive physical factors. For example, the photoreceptor protein rhodopsin. Upon absorption of light, it changes its conformation and excites a nerve impulse.
It consists of a bilipid layer, the lipids of which are strictly oriented - the hydrophobic part of the lipids (tail) is facing inward of the layer, while the hydrophilic part (head) is outward. In addition to lipids, three types of membrane proteins are involved in the construction of the plasma membrane: peripheral, integral, and semi-integral.
One of the areas of study of membranes is currently a detailed study of the properties of both various structural and regulatory lipids and individual integral and semi-integral proteins that make up membranes.
Integral membrane proteins
The main role in the organization of the membrane itself is played by integral and semi-integral proteins, which have a globular structure and are associated with the lipid phase by hydrophilic - hydrophobic interactions. The globules of integral proteins penetrate the entire thickness of the membrane, and their hydrophobic part is located in the middle of the globule and is immersed in the hydrophobic zone of the lipid phase.
Semi-integral membrane proteins
In semi-integral proteins, hydrophobic amino acids are concentrated at one of the poles of the globule, and, accordingly, the globules are only half immersed in the membrane, protruding outward from one (external or internal) membrane surface.
Functions of membrane proteins
Integral and semi-integral proteins of the plasma membrane were previously assigned two functions: general structural and specific. Accordingly, structural and functional proteins were distinguished among them. However, the improvement of methods for isolating protein fractions of membranes and a more detailed analysis of individual proteins now indicate the absence of structural proteins universal for all membranes that do not carry any specific functions. In contrast, membrane proteins with specific functions are very diverse. These are proteins that carry out receptor functions, proteins that are active and passive carriers of various compounds, and finally, proteins that are part of numerous enzyme systems. Material from the site
Properties of membrane proteins
The common property of all these integral and semi-integral membrane proteins, differing not only in functional, but also chemically, is their fundamental ability to move, "float" in the membrane plane in the liquid lipid phase. As noted above, the existence of such movements in the plasma membranes of some cells has been proven experimentally. But this is far from the only type of movement identified in membrane proteins. In addition to lateral displacement, individual integral and semi-integral proteins can rotate in the membrane plane in horizontal and even vertical directions, and can also change the degree of immersion of the molecule in the lipid phase.
Opsin. All these diverse and complex movements of protein globules are especially well illustrated by the example of the opsin protein, which is specific for the membranes of photoreceptor cells (Fig. 3). As you know, opsin in the dark is associated with the carotenoid retinal, which contains a double ciss bond; the complex of retinal and opsin forms rhodopsin, or visual purple. The rhodopsin molecule is capable of lateral movement and rotation in the horizontal plane of the membrane (Fig. 3, A). When exposed to light, retinal undergoes photoisomerization and transforms into a trans form. In this case, the conformation of the retinal changes and it is separated from the opsin, which, in turn, changes the plane of rotation from horizontal to vertical (Fig. 3, B). The consequence of such transformations is a change in the membrane permeability for ions, which leads to the appearance of a nerve impulse.
Interestingly, changes in the conformation of opsin globules induced by light energy can not only serve to generate a nerve impulse, as occurs in the cells of the retina of the eye, but are also the simplest photosynthetic system found in special purple bacteria.
Biological membranes form the basis of the structural organization of the cell. The plasma membrane (plasmalemma) is the membrane that surrounds the cytoplasm of a living cell. Membranes are composed of lipids and proteins. Lipids (mainly phospholipids) form a double layer, in which the hydrophobic "tails" of the molecules are directed towards the inside of the membrane, and the hydrophilic ones - towards its surfaces. Protein molecules can be located on the outer and inner surface of the membrane, they can partially submerge in the lipid layer or penetrate it through and through. Most of the immersed membrane proteins are enzymes. This is a fluid-bone-mosaic model of the structure of a plasma membrane. Protein and lipid molecules are mobile, which ensures the dynamism of the membrane. The membranes also contain carbohydrates in the form of glycolipids and glycoproteins (glycocalyx) located on the outer surface of the membrane. The set of proteins and carbohydrates on the membrane surface of each cell is specific and is a kind of indicator of the cell type.
Membrane functions:
- Separating. It consists in the formation of a barrier between the inner contents of the cell and the external environment.
- Ensuring the exchange of substances between the cytoplasm and the external environment. Water, ions, inorganic and organic molecules enter the cell (transport function). The products formed in the cell (secretory function) are excreted into the external environment.
- Transport. Transport across the membrane can proceed in different ways. Passive transport is carried out without energy consumption, by simple diffusion, osmosis or facilitated diffusion with the help of carrier proteins. Active transport is with the help of carrier proteins, and it requires energy (for example, a sodium-potassium pump). Material from the site
Large molecules of biopolymers enter the cell as a result of endocytosis. It is divided into phagocytosis and pinocytosis. Phagocytosis is the capture and absorption of large particles by the cell. The phenomenon was first described by I.I. Mechnikov. First, the substances adhere to the plasma membrane, to specific receptor proteins, then the membrane sags, forming a depression.
A digestive vacuole is formed. It digests the substances that entered the cell. In humans and animals, leukocytes are capable of phagocytosis. White blood cells absorb bacteria and other particulate matter.
Pinocytosis is the process of capturing and absorbing liquid droplets with substances dissolved in it. Substances adhere to membrane proteins (receptors), and a drop of solution is surrounded by a membrane, forming a vacuole. Pinocytosis and phagocytosis occur with the expenditure of ATP energy.
- Secretory. Secretion - the release by the cell of substances synthesized in the cell into the external environment. Hormones, polysaccharides, proteins, fat droplets are enclosed in vesicles bounded by a membrane, and go to the plasmalemma. The membranes merge, and the contents of the vesicle are discharged into the environment surrounding the cell.
- Connection of cells in tissue (due to folded outgrowths).
- Receptor. Membranes contain a large number of receptors - special proteins, the role of which is to transmit signals from the outside to the inside of the cell.
1. Barrier- provides a regulated, selective, passive and active metabolism with the environment.
Cell membranes have selective permeability: glucose, amino acids, fatty acids, glycerol and ions slowly diffuse through them, the membranes themselves actively regulate this process - some substances are allowed through, while others are not.
2. Transport- through the membrane, substances are transported into and out of the cell. Transport through membranes provides: delivery of nutrients, removal of end metabolic products, secretion of various substances, creation of ionic gradients, maintenance of the appropriate pH and ionic concentration in the cell, which are necessary for the work of cellular enzymes.
There are four main mechanisms for the entry of substances into the cell or their removal from the cell to the outside:
a) Passive (diffusion, osmosis) (does not require energy consumption)
Diffusion
The spread of molecules or atoms of one substance between molecules or atoms of another, leading to a spontaneous equalization of their concentrations throughout the occupied volume. In some situations, one of the substances already has an equalized concentration and they speak of the diffusion of one substance in another. In this case, the transfer of matter occurs from an area with a high concentration to an area with a low concentration (along the vector of the concentration gradient (fig. 2.4).
Rice. 2.4. Diffusion process diagram
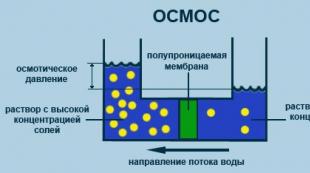
Osmosis
The process of one-way diffusion through a semipermeable membrane of solvent molecules towards a higher concentration of a dissolved substance from a volume with a lower concentration of a dissolved substance (fig. 2.5).

Rice. 2.5. Osmosis process diagram
b) Active transport (requires energy consumption)
Sodium-potassium pump- the mechanism of active conjugated transmembrane transport of sodium ions (from the cell) and potassium ions (into the cell), which provides a concentration gradient and a transmembrane potential difference. The latter serves as the basis for many functions of cells and organs: secretion of glandular cells, muscle contraction, conduction of nerve impulses, etc. (fig. 2.6).

Rice. 2.6. The scheme of the potassium-sodium pump
At the first stage, the enzyme Na + / K + -ATPase attaches three Na + ions from the inner side of the membrane. These ions change the conformation of the active center of the ATPase. After that, the enzyme is able to hydrolyze one ATP molecule. The energy released after hydrolysis is spent on changing the conformation of the carrier, due to which the three Na + ions and the PO 4 3– ion (phosphate) appear on the outer side of the membrane. Here, Na + ions are split off, and PO 4 3– is replaced by two K + ions. After that, the enzyme returns to its original conformation, and K + ions appear on the inner side of the membrane. Here the K + ions are split off, and the carrier is again ready for work.
As a result, a high concentration of Na + ions is created in the extracellular environment, and a high concentration of K + is created inside the cell. This concentration difference is used in cells during the conduction of a nerve impulse.
c) Endocytosis (phagocytosis, pinocytosis)
Phagocytosis(eating by the cell) - the process of absorption of solid objects by the cell, such as eukaryotic cells, bacteria, viruses, the remains of dead cells, etc. A large intracellular vacuole (phagosome) is formed around the absorbed object. The size of phagosomes is from 250 nm and more. By fusing the phagosome with the primary lysosome, a secondary lysosome is formed. In an acidic environment, hydrolytic enzymes break down macromolecules in the secondary lysosome. Cleavage products (amino acids, monosaccharides and other useful substances) are then transported through the lysosomal membrane into the cytoplasm of the cell. Phagocytosis is very widespread. In highly organized animals and humans, the process of phagocytosis plays a protective role. The phagocytic activity of leukocytes and macrophages is of great importance in protecting the body from pathogenic microbes and other unwanted particles entering it. Phagocytosis was first described by the Russian scientist I.I.Mechnikov (fig. 2.7)
Pinocytosis(drinking by the cell) - the process of absorption by the cell of the liquid phase from the environment containing soluble substances, including large molecules (proteins, polysaccharides, etc.). With pinocytosis, small vesicles - endosomes - are detached from the membrane inside the cell. They are smaller than phagosomes (their size is up to 150 nm) and usually do not contain large particles. After the formation of the endosome, the primary lysosome approaches it, and these two membrane vesicles merge. The resulting organelle is called a secondary lysosome. The process of pinocytosis is constantly carried out by all eukaryotic cells. (fig. 7)
Receptor-mediated endocytosis - an active specific process in which the cell membrane bulges into the cell, forming edged pits. The intracellular side of the lined fossa contains a set of adaptive proteins. Macromolecules that bind to specific receptors on the cell surface pass inside at a much higher rate than substances that enter cells through pinocytosis.

Rice. 2.7. Endocytosis
d) Exocytosis (negative phagocytosis and pinocytosis)
A cellular process in which intracellular vesicles (membrane vesicles) fuse with the outer cell membrane. During exocytosis, the contents of the secretory vesicles (exocytic vesicles) are released outside, and their membrane merges with the cell membrane. Almost all macromolecular compounds (proteins, peptide hormones, etc.) are released from the cell by this method. (fig. 2.8)

Rice. 2.8. Exocytosis scheme
3. Generation and conduction of biopotentials- with the help of the membrane, a constant concentration of ions is maintained in the cell: the concentration of the K + ion inside the cell is much higher than outside, and the concentration of Na + is much lower, which is very important, since this ensures the maintenance of the potential difference on the membrane and the generation of a nerve impulse.
4. Mechanical- provides the autonomy of the cell, its intracellular structures, as well as connection with other cells (in tissues).
5. Energy- during photosynthesis in chloroplasts and cellular respiration in mitochondria, energy transfer systems operate in their membranes, in which proteins are also involved;
6. Receptor- some proteins in the membrane are receptors (molecules through which the cell perceives certain signals).
7. Enzymatic- membrane proteins are often enzymes. For example, the plasma membranes of intestinal epithelial cells contain digestive enzymes.
8. Matrix- provides a certain mutual arrangement and orientation of membrane proteins, their optimal interaction;
9. Cage marking- there are antigens on the membrane that act as markers - "labels" that allow you to identify the cell. These are glycoproteins (that is, proteins with branched oligosaccharide side chains attached to them) that play the role of "antennas". With the help of markers, cells can recognize other cells and act in concert with them, for example, during the formation of organs and tissues. It also allows the immune system to recognize foreign antigens.
Cellular inclusions
Cellular inclusions include carbohydrates, fats and proteins. All these substances accumulate in the cytoplasm of the cell in the form of drops and grains of various sizes and shapes. They are periodically synthesized in the cell and used in the metabolic process.
Cytoplasm
It is a part of a living cell (protoplast) without a plasma membrane and nucleus. The composition of the cytoplasm includes: cytoplasmic matrix, cytoskeleton, organelles and inclusions (sometimes inclusions and contents of vacuoles are not classified as living matter of the cytoplasm). Delimited from the external environment by the plasma membrane, the cytoplasm is the internal semi-liquid medium of cells. The nucleus and various organelles are located in the cytoplasm of eukaryotic cells. It also contains a variety of inclusions - products of cellular activity, vacuoles, as well as the smallest tubules and filaments that form the skeleton of the cell. Proteins predominate in the composition of the main substance of the cytoplasm.
Cytoplasmic functions
1) the main metabolic processes take place in it.
2) unites the nucleus and all organelles into one whole, ensures their interaction.
3) mobility, irritability, metabolism and reproduction.
Mobility comes in various forms:
Intracellular movement of the cell cytoplasm.
Amoeba-shaped movement. This form of movement is expressed in the formation of pseudopodia by the cytoplasm towards this or that stimulus or away from it. This form of movement is inherent in amoeba, blood leukocytes, as well as some tissue cells.
Flickering motion. It manifests itself in the form of beating of tiny protoplasmic outgrowths - cilia and flagella (ciliates, epithelial cells of multicellular animals, sperm, etc.).
Contracting movement. It is provided due to the presence in the cytoplasm of a special organoid myofibrils, the shortening or lengthening of which contributes to the contraction and relaxation of the cell. The ability to contract is most developed in muscle cells.
Irritability is expressed in the ability of cells to respond to irritation by changing metabolism and energy.
Cytoskeleton
One of the distinctive features of a eukaryotic cell is the presence in its cytoplasm of skeletal formations in the form of microtubules and bundles of protein fibers. The elements of the cytoskeleton, closely associated with the outer cytoplasmic membrane and nuclear envelope, form complex weaves in the cytoplasm.
The cytoskeleton is formed by microtubules, microfilaments, and the microtrabecular system. The cytoskeleton determines the shape of the cell, participates in cell movements, in the division and movement of the cell itself, in the intracellular transport of organelles.
Microtubules are contained in all eukaryotic cells and are hollow unbranched cylinders, the diameter of which does not exceed 30 nm, and the wall thickness is 5 nm. They can be up to several micrometers in length. Easily disintegrate and reassemble. The wall of microtubules is mainly composed of spirally folded tubulin protein subunits (fig. 2.09)
Microtubule functions:
1) perform a supporting function;
2) form a fission spindle; ensure the divergence of chromosomes to the poles of the cell; are responsible for the movement of cell organelles;
3) take part in intracellular transport, secretion, cell wall formation;
4) are a structural component of cilia, flagella, basal bodies and centrioles.
Microfilaments are represented by filaments with a diameter of 6 nm, consisting of actin protein, which is close to muscle actin. Actin makes up 10-15% of the total protein in the cell. In most animal cells, a dense network of actin filaments and associated proteins forms under the plasma membrane itself.
In addition to actin, myosin filaments are also found in the cell. However, their number is much less. Due to the interaction of actin and myosin, muscle contraction occurs. Microfilaments are associated with the movement of the entire cell or its individual structures within it. In some cases, movement is provided only by actin filaments, in others - by actin together with myosin.
Functions of microfilaments
1) mechanical strength
2) allows the cell to change its shape and move.

Rice. 2.09. Cytoskeleton
Organelles (or organelles)
Are divided into non-membrane, single-membrane and double-membrane.
TO non-membrane organelles eukaryotic cells include organelles that do not have their own closed membrane, namely: ribosomes and organelles built on the basis of tubulin microtubules - cell center (centrioles) and organelles of movement (flagella and cilia). In the cells of most unicellular organisms and the overwhelming majority of higher (terrestrial) plants, centrioles are absent.
TO single-membrane organelles relate: endoplasmic reticulum, Golgi apparatus, lysosomes, peroxisomes, spherosomes, vacuoles and some others. All single-membrane organelles are interconnected into a single cell system. Plant cells have special lysosomes, animal cells have special vacuoles: digestive, excretory, contractile, phagocytic, autophagocytic, etc.
TO two-membrane organelles relate mitochondria and plastids.
Non-membrane organelles
A) Ribosomes- organelles found in the cells of all organisms. These are small organelles, represented by globular particles with a diameter of about 20 nm. Ribosomes are composed of two subunits of unequal size - large and small. Ribosomes include proteins and ribosomal RNA (rRNA). There are two main types of ribosomes: eukaryotic (80S) and prokaryotic (70S).
Depending on the localization in the cell, there are free ribosomes located in the cytoplasm that synthesize proteins and attached ribosomes - ribosomes connected by large subunits with the outer surface of the EPR membranes, synthesizing proteins that enter the Golgi complex, and then are secreted by the cell. During protein biosynthesis, ribosomes can combine into complexes - polyribosomes (polysomes).
Eukaryotic ribosomes are formed in the nucleolus. First, rRNA is synthesized on the nucleolar DNA, which is then covered with ribosomal proteins coming from the cytoplasm, cleaved to the required size and forming ribosome subunits. There are no fully formed ribosomes in the nucleus. The union of subunits into a whole ribosome occurs in the cytoplasm, usually during protein biosynthesis.
Ribosomes are found in the cells of all organisms. Each consists of two particles, small and large. The ribosomes include proteins and RNA.
Functions
protein synthesis.
The synthesized proteins first accumulate in the channels and cavities of the endoplasmic reticulum, and then are transported to the organelles and parts of the cell. EPS and ribosomes located on its membranes represent a single apparatus for the biosynthesis and transport of proteins (fig. 2.10-2.11).

Rice. 2.10. Ribosome structure

Rice. 2.11. Ribosome structure
B) Cell center (centrioles)
The centriole is a cylinder (0.3 µm long and 0.1 µm in diameter), the wall of which is formed by nine groups of three merged microtubules (9 triplets), interconnected at certain intervals by cross-linking. Centrioles are often paired, where they are at right angles to each other. If the centriole lies at the base of the cilium or flagellum, then it is called the basal body.
Almost all animal cells have a pair of centrioles, which are the middle element of the cell center.
Before division, the centrioles diverge to opposite poles and a daughter centriole appears near each of them. From centrioles located at different poles of the cell, microtubules are formed, growing towards each other.
Functions
1) form a mitotic spindle, which contributes to the uniform distribution of genetic material between daughter cells,
2) are the center of the organization of the cytoskeleton. Some of the spindle filaments are attached to the chromosomes.
Centrioles are self-replicating organelles of the cytoplasm. They arise as a result of duplication of existing ones. This occurs when the centrioles diverge. Immature centriole contains 9 single microtubules; apparently, each microtubule is a matrix for the assembly of triplets characteristic of a mature centriole (fig. 2.12).
Cetriols are found in the cells of lower plants (algae).

Rice. 2.12. Cell center centrioles
Single membrane organelles
D) Endoplasmic reticulum (EPS)
The entire inner zone of the cytoplasm is filled with numerous small channels and cavities, the walls of which are membranes similar in structure to the plasma membrane. These channels branch, connect to each other and form a network called the endoplasmic reticulum. The endoplasmic reticulum is heterogeneous in structure. Two types of it are known - granular and smooth.
On the membranes of the channels and cavities of the granular network, there are many small rounded bodies - ribosome which give the membranes a rough appearance. The membranes of the smooth endoplasmic reticulum do not carry ribosomes on their surface. The EPS performs many different functions.
Functions
The main function of the granular endoplasmic reticulum is participation in protein synthesis, which is carried out in the ribosomes. On the membranes of the smooth endoplasmic reticulum, lipids and carbohydrates are synthesized. All these synthesis products accumulate in channels and cavities, and then are transported to various organelles of the cell, where they are consumed or accumulate in the cytoplasm as cellular inclusions. EPS connects the main organelles of the cell (fig. 2.13).

Rice. 2.13. The structure of the endoplasmic reticulum (EPS) or reticulum
E) Golgi apparatus
The structure of this organoid is similar in the cells of plant and animal organisms, despite the diversity of its shape. It has many important functions.
Single membrane organoid. It is a stack of flattened "cisterns" with extended edges, with which a system of small one-membrane bubbles (Golgi bubbles) is connected. Golgi bubbles are mainly concentrated on the side adjacent to the EPS and on the periphery of the stacks. It is believed that they transfer proteins and lipids into the Golgi apparatus, the molecules of which, moving from cistern to cistern, undergo chemical modification.
All these substances first accumulate, chemically complicate, and then in the form of large and small bubbles enter the cytoplasm and are either used in the cell itself during its life, or are removed from it and used in the body (fig. 2.14-2.15).

Rice. 2.14. The structure of the Golgi apparatus
Functions:
Modification and accumulation of proteins, lipids, carbohydrates;
Packing in membrane vesicles (vesicles) of incoming organic substances;
Place of formation of lysosomes;
Secretory function, therefore, the Golgi apparatus is well developed in secretory cells.

Rice. 2.15. Golgi complex
E) Lysosomes
They are small rounded bodies. Inside the lysosome are enzymes that break down proteins, fats, carbohydrates, nucleic acids. Lysosomes approach the food particle that entered the cytoplasm, merge with it, and one digestive vacuole is formed, inside which there is a food particle surrounded by lysosomal enzymes.
Lysosomal enzymes are synthesized on a rough EPS, transferred to the Golgi apparatus, where they are modified and packaged into membrane vesicles of lysosomes. The lysosome can contain from 20 to 60 different types of hydrolytic enzymes. The breakdown of substances using enzymes is called lysis.
Distinguish between primary and secondary lysosomes. Primary lysosomes are called that have budded from the Golgi apparatus.
Secondary lysosomes are called, formed as a result of the fusion of primary lysosomes with endocytic vacuoles. In this case, they digest the substances that entered the cell by phagocytosis or pinocytosis, so they can be called digestive vacuoles.
Functions of lysosomes:
1) digestion of substances or particles (bacteria, other cells) captured by the cell during endocytosis,
2) autophagy - the destruction of structures unnecessary for the cell, for example, during the replacement of old organelles with new ones, or the digestion of proteins and other substances produced inside the cell itself,
3) autolysis - self-digestion of a cell, leading to its death (sometimes this process is not pathological, but accompanies the development of the body or the differentiation of some specialized cells) (fig. 2.16-2.17).
Example: When a tadpole turns into a frog, the lysosomes in the tail cells digest it: the tail disappears, and the substances formed during this process are absorbed and used by other cells of the body.

Rice. 2.16. Lysosome formation

Rice. 2.17. Functioning of lysosomes
G) Peroxisomes
Organoids, similar in structure to lysosomes, vesicles with a diameter of up to 1.5 microns with a homogeneous matrix containing about 50 enzymes.
Catalase causes the decomposition of hydrogen peroxide 2H 2 O 2 → 2H 2 O + O 2 and prevents lipid peroxidation
Peroxisomes are formed by budding from previously existing ones, i.e. belong to self-reproducing organelles, despite the fact that they do not contain DNA. They grow due to the entry of enzymes into them, peroxisome enzymes are formed on the rough EPS and in the hyaloplasm (fig. 2.18).

Rice. 2.18. Peroxisome (in the center of the crystalline nucleoid)
H) Vacuoles
Single membrane organelles. Vacuoles are "containers" filled with aqueous solutions of organic and inorganic substances. The EPS and the Golgi apparatus are involved in the formation of vacuoles.
Young plant cells contain many small vacuoles, which then, as the cells grow and differentiate, merge with each other and form one large central vacuole.
The central vacuole can occupy up to 95% of the volume of a mature cell, while the nucleus and organelles are pushed back to the cell membrane. The membrane that limits the plant vacuole is called tonoplast.
The fluid that fills the plant vacuole is called cell sap. The composition of cell sap includes water-soluble organic and inorganic salts, monosaccharides, disaccharides, amino acids, end or toxic metabolic products (glycosides, alkaloids), some pigments (anthocyanins).
Of organic substances, sugars and proteins are more often stored. Sugars - more often in the form of solutions, proteins come in the form of EPR bubbles and the Golgi apparatus, after which the vacuoles are dehydrated, turning into aleurone grains.
In animal cells, there are small digestive and autophagic vacuoles belonging to the group of secondary lysosomes and containing hydrolytic enzymes. In unicellular animals, there are also contractile vacuoles that perform the function of osmoregulation and excretion.
Functions
In plants
1) accumulation of fluid and maintenance of turgor,
2) accumulation of reserve nutrients and mineral salts,
3) coloring of flowers and fruits and thus attracting pollinators and distributors of fruits and seeds.
In animals:
4) digestive vacuoles - destroy organic macromolecules;
5) contractile vacuoles regulate the osmotic pressure of the cell and remove unnecessary substances from the cell
6) phagocytic vacuoles are formed during phagocytosis by immune cells of antigens
7) autophagocytic vacuoles are formed during phagocytosis by immune cells of their own tissues
Two-membrane organelles (mitochondria and plastids)
These organelles are semi-autonomous, since they have their own DNA and their own protein-synthesizing apparatus. Mitochondria are found in almost all eukaryotic cells. Plastids are found only in plant cells.
I) Mitochondria
These are organelles of energy supply for metabolic processes in the cell. In the hyaloplasm, mitochondria are usually distributed diffusely, but in specialized cells they are concentrated in those areas where there is the greatest need for energy. For example, in muscle cells, large numbers of mitochondria are concentrated along the contractile fibrils, along the sperm flagellum, in the epithelium of the renal tubules, in the area of synapses, etc. This arrangement of mitochondria provides less loss of ATP during its diffusion.
The outer membrane separates the mitochondria from the cytoplasm, is closed on itself and does not form invaginations. The inner membrane limits the inner content of the mitochondria - the matrix. A characteristic feature is the formation of numerous invaginations - cristae, due to which the area of the internal membranes increases. The number and degree of development of cristae depends on the functional activity of the tissue. Mitochondria have their own genetic material (fig. 2.19).
Mitochondrial DNA is a closed circular double-stranded molecule, in human cells it has a size of 16569 nucleotide pairs, which is approximately 105 times smaller than the DNA localized in the nucleus. Mitochondria have their own protein-synthesizing system, while the number of proteins translated from mitochondrial mRNA is limited. Mitochondrial DNA cannot encode all mitochondrial proteins. Most of the proteins in mitochondria are under the genetic control of the nucleus.

Rice. 2.19. Mitochondrial structure
Mitochondrial functions
1) the formation of ATP
2) protein synthesis
3) participation in specific syntheses, for example, the synthesis of steroid hormones (adrenal glands)
4) spent mitochondria can also accumulate excretion products, harmful substances, i.e. able to take on the functions of other cell organelles
K) Plastids
Plastids–Organelles, characteristic only of plants.
There are three types of plastids:
1) chloroplasts(green plastids);
2) chromoplasts(yellow, orange or red plastids)
3) leukoplasts(colorless plastids).
Usually, only one type of plastids are found in a cell.
Chloroplasts
These organelles are found in the cells of leaves and other green plant organs, as well as in a variety of algae. Higher plants usually contain several tens of chloroplasts in one cell. The green color of chloroplasts depends on the content of chlorophyll pigment in them.
Chloroplast is the main organoid of plant cells, in which photosynthesis takes place, that is, the formation of organic substances (carbohydrates) from inorganic (CO 2 and H 2 O) using the energy of sunlight. Chloroplasts are structurally similar to mitochondria.
Chloroplasts have a complex structure. They are delimited from the hyaloplasm by two membranes - external and internal. The inner content is called stroma... The inner membrane forms a complex, strictly ordered system of membranes in the form of flat bubbles, called thylakoids.
Thylakoids are collected in piles - grains resembling columns of coins . Grana are connected to each other by stromal thylakoids, passing through them along the plastid (fig. 2.20-2.22). Chlorophyll and chloroplasts are produced only by exposure to light.

Rice. 2.20. Chloroplasts under a light microscope

Rice. 2.21. Chloroplast structure under an electron microscope

Rice. 2.22. Schematic structure of chloroplasts
Functions
1) photosynthesis(the formation of organic substances from inorganic substances due to the energy of light). Chlorophyll plays a central role in this process. It absorbs light energy and directs it to carry out photosynthetic reactions. In chloroplasts, as in mitochondria, ATP is synthesized.
2) participate in the synthesis of amino acids and fatty acids,
3) serve as a repository of temporary starch reserves.
Leukoplasts- small colorless plastids that are found in the cells of organs hidden from sunlight (roots, rhizomes, tubers, seeds). Their structure is similar to the structure of chloroplasts. (fig. 2.23).
However, unlike chloroplasts, leukoplasts have a poorly developed internal membrane system, because they are involved in the synthesis and accumulation of reserve nutrients - starch, proteins and lipids. In the light, leukoplasts can turn into chloroplasts.

Rice. 2.23. Leukoplast structure
Chromoplasts- plastids of orange, red and yellow color, which is caused by pigments belonging to the group of carotenoids. Chromoplasts are found in the cells of the petals of many plants, mature fruits, rarely root crops, as well as in autumn leaves. The internal membrane system in chromoplasts, as a rule, is absent (fig. 24).

Rice. 2.24. Chromoplast structure
The significance of chromoplasts has not yet been fully elucidated. Most of them are aging plastids. They, as a rule, develop from chloroplasts, while chlorophyll and the inner membrane structure are destroyed in the plastids, and carotenoids accumulate. This happens when the fruits ripen and the leaves turn yellow in autumn. The biological significance of chromoplasts lies in the fact that they determine the bright color of flowers and fruits, which attracts insects for cross-pollination and other animals for spreading fruits. Leukoplasts can also turn into chromoplasts.
Functions of plastids
Synthesis of organic substances in chlorophyll from simple inorganic compounds: carbon dioxide and water in the presence of quanta of sunlight - photosynthesis, ATP synthesis in the light phase of photosynthesis
Synthesis of proteins on ribosomes (between the inner membranes of the chloroplast there are DNA, RNA and ribosomes, therefore, in chloroplasts, as well as in mitochondria, protein is synthesized, which is necessary for the activity of these organelles).
The presence of chromoplasts is explained by the yellow, orange and red color of the corolla of flowers, fruits, autumn leaves.
Leukoplasts contain storage substances (in stems, roots, tubers).
Chloroplasts, chromoplasts and leukoplasts are capable of inter-cell transition. So, when fruits ripen or change the color of leaves in autumn, chloroplasts turn into chromoplasts, and leukoplasts can turn into chloroplasts, for example, when potato tubers become green.
In an evolutionary sense, the primary, initial type of plastids are chloroplasts, from which the plastids of the other two types originated. Plastids share many similarities with mitochondria, distinguishing them from other components of the cytoplasm. This is, first of all, a shell of two membranes and relative genetic autonomy, due to the presence of its own ribosomes and DNA. This peculiarity of organelles formed the basis for the idea that the precursors of plastids and mitochondria were bacteria, which in the process of evolution turned out to be built into a eukaryotic cell and gradually turned into chloroplasts and mitochondria. (fig. 2.25).

Rice. 2.25. The formation of mitochondria and chloroplasts according to the theory of symbiogenesis
Cell- an elementary unit of a living system. Various structures of a living cell, which are responsible for the performance of a particular function, are called organelles, like the organs of the whole organism. Specific functions in the cell are distributed between organelles, intracellular structures that have a certain shape, such as the cell nucleus, mitochondria, etc.
Cellular structures:
Cytoplasm... The obligatory part of the cell, enclosed between the plasma membrane and the nucleus. Cytosol Is a viscous aqueous solution of various salts and organic substances, permeated by a system of protein filaments - cytoskeletons. Most of the chemical and physiological processes of the cell take place in the cytoplasm. Structure: Cytosol, cytoskeleton. Functions: includes various organelles, the internal environment of the cell
Plasma membrane... Every cell of animals, plants, is limited from the environment or other cells by the plasma membrane. The thickness of this membrane is so small (about 10 nm) that it can only be seen with an electron microscope.

Lipids they form a double layer in the membrane, and proteins permeate its entire thickness, are immersed at different depths in the lipid layer, or are located on the outer and inner surfaces of the membrane. The structure of the membranes of all other organelles is similar to the plasma membrane. Structure: double layer of lipids, proteins, carbohydrates. Functions: limiting, maintaining the shape of the cell, protecting against damage, regulating the intake and removal of substances.
Lysosomes... Lysosomes are membrane organelles. They have an oval shape and a diameter of 0.5 microns. They contain a set of enzymes that destroy organic matter. The lysosome membrane is very strong and prevents the penetration of its own enzymes into the cytoplasm of the cell, but if the lysosome is damaged by any external influences, then the whole cell or part of it is destroyed.
Lysosomes are found in all cells of plants, animals, and fungi.
Digesting various organic particles, lysosomes provide additional "raw material" for chemical and energy processes in the cell. During starvation, lysosome cells digest some organelles without killing the cell. This partial digestion provides the cell with the necessary minimum of nutrients for some time. Sometimes lysosomes digest whole cells and groups of cells, which plays an essential role in the development of animals. An example is the loss of a tail when a tadpole transforms into a frog. Structure: oval-shaped vesicles, membrane outside, enzymes inside. Functions: splitting organic matter, destroying dead organelles, destroying spent cells.
Golgi complex... The products of biosynthesis entering the lumens of the cavities and tubules of the endoplasmic reticulum are concentrated and transported in the Golgi apparatus. This organoid is 5–10 µm in size.

Structure: membrane-surrounded cavities (vesicles). Functions: accumulation, packaging, excretion of organic matter, formation of lysosomes
Endoplasmic reticulum... The endoplasmic reticulum is a system for the synthesis and transport of organic substances in the cytoplasm of the cell, which is an openwork structure of connected cavities.
A large number of ribosomes are attached to the membranes of the endoplasmic reticulum - the smallest organelles of the cell, which look like a sphere with a diameter of 20 nm. and consisting of RNA and protein. Protein synthesis occurs on ribosomes. Then the newly synthesized proteins enter the system of cavities and tubules, along which they move inside the cell. Cavities, tubules, tubules from membranes, on the surface of the ribosome membranes. Functions: synthesis of organic substances using ribosomes, transport of substances.
Ribosomes... Ribosomes are attached to the membranes of the endoplasmic reticulum or are freely located in the cytoplasm, they are located in groups, proteins are synthesized on them. Protein composition, ribosomal RNA Functions: provides protein biosynthesis (assembly of a protein molecule from).
Mitochondria... Mitochondria are energy organelles. The shape of mitochondria is different, they can be the rest, rod-shaped, filamentous with an average diameter of 1 micron. and a length of 7 microns. The number of mitochondria depends on the functional activity of the cell and can reach tens of thousands in the flying muscles of insects. The mitochondria are bounded on the outside by the outer membrane, underneath is the inner membrane, which forms numerous outgrowths - cristae.

Inside the mitochondria are RNA, DNA, and ribosomes. Specific enzymes are built into its membranes, with the help of which the energy of food substances is converted into the energy of ATP, which is necessary for the life of the cell and the body as a whole, in the mitochondria.
Membrane, matrix, outgrowths - cristae. Functions: synthesis of the ATP molecule, the synthesis of its own proteins, nucleic acids, carbohydrates, lipids, the formation of its own ribosomes.
Plastids... Only in a plant cell: lecoplasts, chloroplasts, chromoplasts. Functions: accumulation of reserve organic substances, attraction of pollinating insects, synthesis of ATP and carbohydrates. Chloroplasts are shaped like a disc or sphere with a diameter of 4–6 µm. With double membrane - outer and inner. Inside the chloroplast, there are ribosome DNA and special membrane structures - grains, connected to each other and to the inner membrane of the chloroplast. Each chloroplast contains about 50 grains, staggered for better light capture. Chlorophyll is located in the gran membranes, thanks to it, the energy of sunlight is converted into the chemical energy of ATP. ATP energy is used in chloroplasts for the synthesis of organic compounds, primarily carbohydrates.
Chromoplasts... The red and yellow pigments found in chromoplasts give different parts of the plant a red and yellow color. carrots, tomato fruits.
Leukoplasts are a place of accumulation of a reserve nutrient - starch. There are especially many leukoplasts in the cells of potato tubers. In the light, leukoplasts can turn into chloroplasts (as a result of which the potato cells turn green). In autumn, chloroplasts turn into chromoplasts and green leaves and fruits turn yellow and red.
Cell center... Consists of two cylinders, centrioles, located perpendicular to each other. Functions: thread support for dividing spindles

Cellular inclusions appear in the cytoplasm, then disappear in the process of cell life.

Dense, in the form of granules, inclusions contain reserve nutrients (starch, proteins, sugars, fats) or waste products of the cell, which cannot yet be removed. All plastids of plant cells have the ability to synthesize and accumulate reserve nutrients. In plant cells, the accumulation of reserve nutrients occurs in the vacuoles.
Grains, granules, drops Functions: non-permanent formations that store organic matter and energy
Core... Nuclear membrane of two membranes, nuclear juice, nucleolus. Functions: storage of hereditary information in the cell and its reproduction, synthesis of RNA - informational, transport, ribosomal. There are spores in the nuclear membrane, through which an active exchange of substances between the nucleus and the cytoplasm takes place. The nucleus stores hereditary information not only about all the signs and properties of a given cell, about the processes that must occur to it (for example, protein synthesis), but also about the characteristics of the organism as a whole. Information is recorded in DNA molecules, which are the main part of chromosomes. There is a nucleolus in the nucleus. The nucleus, due to the presence in it of chromosomes containing hereditary information, performs the functions of a center that controls all vital activity and development of the cell.