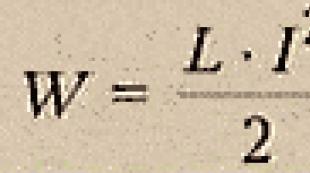Magnifik och mystisk elektrisk ål. Elektrokemisk protongradient En ökning av den elektrokemiska gradienten kommer att resultera i
Frånkopplare som dinitrofenol gör att H läcker över membranet, vilket kraftigt minskar den elektrokemiska protongradienten. Oligomycin blockerar specifikt protonflödet genom Rc
| Ris. 7-53. Förändringar i redoxpotentialen under passage av elektroder under fotosyntes med bildning av NADPH och ATP) i växter och cyanobakterier. Fotosystem II är mycket likt reaktionscentrumet för lila bakterier (se figur 7-50) som det är evolutionärt förknippat med. Fotosystem I skiljer sig från dessa två system eftersom man tror att det är evolutionärt relaterat till fotosystemen hos en annan grupp av prokaryoter - gröna bakterier. I fotosystem I passerar elektronerna i det exciterade klorofyllet genom en serie tätt bundna järn-svavelcentra. Två seriekopplade fotosystem ger det totala flödet av elektroner från vatten till NADP med bildandet av NADPH. Dessutom bildas ATP med hjälp av ATP-syntetas (ej visat) på grund av energin från den elektrokemiska protongradienten, som skapas av elektrontransportkedjan som länkar fotosystem II med fotosystem I. Detta Z-schema för ATP-bildning kallas icke-cykliskt fosforylering, i motsats till det cykliska schemat som visas i ris. 7-54 (se även figur 7-52). |
När ett frånkopplingsmedel såsom dinitrofenol tillsätts till cellerna, ökar syreupptaget av mitokondrierna kraftigt när hastigheten för elektronöverföring ökar. Denna acceleration är förknippad med förekomsten av andningskontroll. Denna kontroll tros vara baserad på den direkta hämmande effekten av den elektrokemiska protongradienten på elektrontransport. När, i närvaro av en frånkopplare, den elektrokemiska gradienten försvinner, når den okontrollerade elektrontransporten den maximala hastigheten som är möjlig med en given mängd substrat. Tvärtom saktar en ökning av protongradienten ner elektrontransporten och processen saktar ner. Dessutom, om, i ett experiment, en ovanligt hög elektrokemisk gradient skapas på konstgjord väg på det inre membranet, kommer normal elektrontransport att sluta helt, och i vissa delar av andningskedjan kommer det att vara möjligt att upptäcka ett omvänt flöde av elektroner. observation tyder på att andningskontroll helt enkelt återspeglar en balans mellan energi för att pumpa protoner, kopplat till transporten av elektroner, och för transporten av elektroner i sig, eller, med andra ord, att storleken på den elektrokemiska protongradienten påverkar både hastigheten och riktning för elektronöverföring, i princip på samma sätt som verkningsriktningen för ATP-syntetas (avsnitt 9.2.3).
Den energi som frigörs under överföringen av elektroner längs andningskedjan lagras i form av en elektrokemisk protongradient på det inre mitokondriella membranet
pP (ArP)-gradienten tvingar P-jonerna tillbaka in i matrisen, och OP-jonerna från matrisen, vilket förstärker effekten av membranpotentialen (AU), under vars verkan eventuell positiv laddning dras in i matrisen, och någon negativ laddning trycks ut ur den. Den kombinerade verkan av dessa två krafter resulterar i en elektrokemisk protongradient (Figur 7-19).
Nästan alla bakterier, inklusive strikta anaerober, upprätthåller en protonmotorisk kraft på sitt membran. Energin från den elektrokemiska protongradienten används i dem för att rotera det bakteriella flagellumet, vilket gör att cellen kan röra sig (avsnitt 12.5.4), och för
Energin i den elektrokemiska protongradienten används för syntes av ATP och transport av metaboliter och oorganiska joner in i matrisen.
I fig. 7-34 visar nivåerna av redoxpotential på olika platser i andningskedjan. En dramatisk minskning inträffar inom vart och ett av de tre stora andningskomplexen. Potentialskillnaden mellan två valfria elektronbärare är direkt proportionell mot den energi som frigörs när en elektron passerar från en bärare till en annan (Figur 7-34). Varje komplex fungerar som en energiomvandlande enhet som kanaliserar denna fria energi för att flytta protoner över membranet, vilket resulterar i en elektrokemisk protongradient när elektroner färdas genom kretsen. Denna energiomvandling kan påvisas direkt genom att separat inkorporera eventuellt isolerat respiratorisk kedjekomplex i liposomer (se figur 7-25). I närvaro av en lämplig elektrondonator och -acceptor kommer ett sådant komplex att bära elektroner, vilket resulterar i att protoner pumpas över liposommembranet.
Respiratoriska enzymkomplex kombinerar transport av elektroner, åtföljd av frigöring av energi, med pumpning av protoner från matrisen. Den elektrokemiska protongradienten som skapas i detta fall levererar energi för syntesen av ATP av ett annat transmembranproteinkomplex - ATP-syntetas, genom vilket protoner återförs till matrisen. ATP-syntetas är ett reversibelt konjugerande komplex som normalt omvandlar energin från protonflödet som riktas till matrisen till energin av ATP-fosfatbindningar, men med en minskning av den elektrokemiska protongradienten kan den också använda energin från ATP-hydrolys för att flytta protoner ut ur matrisen. Kemiosmotiska mekanismer är karakteristiska för både mitokondrier och kloroplaster, och bakterier, vilket indikerar deras exceptionella betydelse för alla celler.
När högenergielektroner passerar genom andningskedjan pumpas protoner ut ur matrisen i var och en av dess tre energilagrande regioner. Som ett resultat uppstår en elektrokemisk protongradient mellan de två sidorna av det inre membranet, under vars verkan protoner återförs tillbaka till matrisen genom ATP-syntetas - ett transmembranenzymkomplex som använder energin från protonström för att syntetisera ATP från ADP och P.
| Ris. 9-36. Den protonmotoriska kraften som genereras på det bakteriella plasmamembranet säkerställer förflyttning av näringsämnen in i cellen och utsöndring av natrium utanför. I närvaro av syre (A) skapar andningskedjan av aeroba bakterier en elektrokemisk protongradient, som används av ATP-syntetas för att syntetisera ATP. Under anaeroba förhållanden (B) får samma bakterier ATP som ett resultat av glykolys. På grund av hydrolysen av en del av denna ATP under verkan av ATP-syntetas, uppstår en transmembran proton-motorkraft, som utför transportprocesser. (Som beskrivs i texten finns det bakterier där elektrontransportkedjan pumpar ut protoner och under anaeroba förhållanden är den slutliga elektronacceptorn i detta fall inte syre, utan andra molekyler.) |
En elektrokemisk protongradient producerar en protoninjektionskraft, mätt i millivolt (mV). Eftersom pP (ApH)-gradienten i 1 pH-enhet är ekvivalent med en membranpotential på cirka 60 mV, kommer den protonmotoriska kraften att vara lika med L - 60 (ApH). I en typisk cell är denna kraft på det inre membranet av andningsmitokondrierna cirka 220 mV och består av en membranpotential på cirka 160 mV och en pH-gradient. nära -] pH-enhet.
Enligt syntesen av ATP är inte den enda processen som sker på grund av energin i den elektrokemiska gradienten. I matrisen, där enzymerna som är involverade i citronsyracykeln och andra metaboliska reaktioner finns, är det nödvändigt att upprätthålla höga koncentrationer av olika substrat, i synnerhet krävs ADP och fosfat för ATP-syntetas. Därför måste en mängd olika laddningsbärande substrat transporteras över det inre membranet. Detta uppnås med hjälp av olika bärarproteiner inbyggda i membranet (se avsnitt 6.4.4). av vilka många aktivt pumpar vissa molekyler mot sina elektrokemiska gradienter, det vill säga de genomför en process som kräver energiförbrukning. För de flesta av metaboliterna är källan till denna energi konjugering med rörelsen av vissa andra molekyler nedför deras elektrokemiska gradient (se avsnitt 6.4.9). Till exempel deltar ADP-ATP-antiportsystemet i ADP-transporten; under övergången av varje ADP-molekyl till matrisen kommer en ATP-molekyl ut från den längs dess elektrokemiska gradient. Samtidigt konjugerar symportsystemet övergången av fosfat till mitokondrierna med flödet av P riktat dit, protoner kommer in i matrisen längs sin gradient och flyttar samtidigt fosfat bakom sig. På liknande sätt överförs det till matrisen och pyruvat (fig. 7-21). Energin i den elektrokemiska protongradienten används också för att överföra Ca-joner till matrisen, som uppenbarligen spelar en viktig roll i regleringen av aktiviteten hos vissa mitokondriella enzymer, absorptionen av dessa joner av mitokondrier för att avlägsna dem från cytosolen , när Ca-koncentrationen i den senare blir farlig hög (se avsnitt 12.3.7).
Verkan av ATP-syntetas är reversibel, den kan använda både energin från ATP-hydrolys för att pumpa protoner genom det inre mitokondriella membranet och energin hos protonflödet längs den elektrokemiska gradienten för syntesen av ATP (fig. 7-26) ). Således är ATP-syntetas ett reversibelt kopplingssystem som interkonverterar energin hos den elektrokemiska protongradienten och kemiska bindningar. Regleringen av dess funktion beror på förhållandet mellan protongradientens branthet och det lokala värdet av AG för hydrolys av ATP.
Vi har tidigare visat att den fria energin vid ATP-hydrolys beror på koncentrationen av tre reaktanter - ATP, ADP och Pi (se fig. 7-22). AG för syntesen av ATP är samma värde taget med ett minus. Den fria energin för rörelse av protoner genom membranet är lika med summan (1) AG för rörelsen av en mol av alla joner mellan regioner med skillnaden i potentialerna AV och (2) AG för rörelsen av en mol av alla molekyler mellan regioner med olika koncentrationer. Ekvationen för proton-motorkraften som ges i Sec. 7.1.7, kombinerar samma komponenter, men endast koncentrationsskillnaden ersätts av en ekvivalent ökning av membranpotentialen, så att ett uttryck för protonens elektrokemiska potential erhålls. Således tar AG för rörelsen av protoner och protonmotorkraften hänsyn till samma potential, bara i det första fallet mäts det i kilokalorier och i det andra - i millivolt. Faktorn för att konvertera från en enhet till en annan är Faraday-talet. Således är AGh = -0,023 (protonmotorkraft), där AGh + uttrycks i kilokalorier per mol (kcal / mol) och protonmotorkraft uttrycks i millivolt (mV). Om den elektrokemiska protongradienten är 220 mV, så är AGh = 5,06
Om ATP-syntetas normalt inte transporterar P från matrisen, så överför andningskedjan i det inre mitokondriella membranet, under normala förhållanden, protoner genom detta membran, vilket skapar en elektrokemisk protongradient som levererar energi för syntesen av ATP. Under vissa förhållanden är det möjligt att experimentellt demonstrera andningskedjans förmåga att pumpa ut protoner från matrisen. Det är till exempel möjligt att tillhandahålla en suspension av isolerade mitokondrier med ett lämpligt substrat för oxidation och blockera protonflödet genom ATP-syntetas. Under anaeroba förhållanden kommer en liten tillsats av syre till ett sådant läkemedel att orsaka en explosion av andningsaktivitet som kommer att ta en till två sekunder - tills allt syre är förbrukat.Med hjälp av en känslig rP-elektrod kan en plötslig försurning av mediet till följd av utdrivningen av P-joner från mitokondriernas matris registreras i en sådan skur av andning.
| Ris. 7-36. Överföring av protoner över det inre mitokondriella membranet med deltagande av frikopplingsmedlet 2,4-dinitrofenol (DNP) Den laddade (protonerade) formen av DNP kan fritt |
Teiko Abe
Hur leder förlusten av en elektrokemisk gradient till värmeutveckling?
Såvitt jag förstår ger frikopplingen av protonflödet och ATP-syntas en protonbypass mellan det yttre och inre mitokondriella membranet, så att protoner inte behöver passera genom ATP-syntaset på väg till matrisen. Jag kan se hur detta resulterar i förlusten av den elektrokemiska gradienten. Men varför genereras värme?
AliceD ♦
Av samma anledning som när batteriet är kortslutet :) Samma princip, samma effekt.
Svar
Satwik Pasani
ALiceDs kommentar är helt sann. (Även om kortslutningen i verkliga fall sällan är absolut, eftersom det vanligtvis finns ett visst ändligt motstånd i kortslutningstråden.)
Du kan förstå detta på två sätt.
Intuitivt förståeligt att frikopplingen ger en kanal för vätejonernas rörelse över membranet i riktning mot deras elektrokemiska gradient utan något arbete. Därför kan energin som den får genom att korsa potentialskillnaden anses omvandlad till kinetisk energi, det vill säga att vätejonerna accelereras av potentialskillnaden, vilket gör att de tar upp hastigheten och därför rör sig med en högre hastighet än genomsnittet av det sista facket. Detta kommer att leda till fler kollisioner (och mer energiska sådana) med omgivande molekyler, vilket också kommer att öka deras kinetiska energi något, vilket i slutändan kommer att öka den genomsnittliga kinetiska energin, ett mått på vilket kallas temperatur. Om det var bundet skulle vätejonerna inte få kinetisk energi, eftersom energin de får genom att korsa potentialskillnaden skulle användas för att arbeta i ATP-syntasmekanismen.
Strängt talat du kan visa detta med hjälp av kemisk termodynamik, vilket innebär att använda Δ G "roll =" presentation "style =" position: relativ; "> Δ G Δ G "roll =" presentation "style =" position: relativ; "> Δ G "roll =" presentation "style =" position: relativ; "> Δ Δ G "roll =" presentation "style =" position: relativ; "> g funktioner, μ "roll =" presentation "style =" position: relativ; "> μ μ "roll =" presentation "style =" position: relativ; "> μ "roll =" presentation "style =" position: relativ; "> μ funktioner och några relaterade termodynamiska variabler. Låt mig veta om du vill ha den här förklaringen (även om jag riskerar att tappa kontakten med den matematiska aspekten av termodynamiken)
PS: - Även om den termodynamiska förklaringen också tar hänsyn till temperaturökningen på grund av den suddiga koncentrationsgradienten, är det svårt att förklara med den tidigare modellen. Du kan tänka på detta som att neutralisering av koncentrationsgradienten ändrar antalet kollisioner per volymenhet (och tid), och bidrar därför också till den observerade förändringen i temperatur.
WYSIWYG ♦
Sättet du förklarade mekanismen på är mycket bra +1
Teiko Abe
Tack för ditt svar. Nu är jag ganska nöjd med det intuitiva tillvägagångssättet. Jag är inte säker på om jag helt kan förstå den rigorösa matematiska förklaringen vid det här laget, jag känner att jag måste läsa lite först.
Elektrokemisk gradient, eller elektrokemisk potentialgradient- en uppsättning koncentrationsgradienter och membranpotential, som bestämmer jonernas rörelseriktning genom membranet. Den består av två komponenter: en kemisk gradient (koncentrationsgradient), eller skillnaden i koncentrationen av ett löst ämne på båda sidor av membranet, och en elektrisk gradient (membranpotential), eller en skillnad i laddningar på motsatta sidor av membranet. membran. Gradienten uppstår på grund av den ojämna koncentrationen av joner på motsatta sidor av det permeabla membranet. Joner rör sig genom membranet från ett område med högre koncentration till ett område med lägre koncentration genom enkel diffusion. Dessutom bär joner en elektrisk laddning, som bildar en elektrisk potential på membranet (membranpotential). Om det finns en ojämn fördelning av laddningar på båda sidor av membranet, så genererar skillnaden i elektrisk potential en kraft som leder till jondiffusion tills laddningarna på båda sidor är balanserade.
Collegiate YouTube
1 / 3
✪ Membranepotentialer - Del 1
✪ In Da Club - Membranes & Transport: Crash Course Biology # 5
✪ Neuronala synapser (kemiska) | Människans anatomi och fysiologi | Hälsa & medicin | Khan akademin
undertexter
Jag ska rita en liten bur. Detta kommer att vara en typisk cell och den är fylld med kalium. Vi vet att celler älskar att samla det inom sig. Mycket kalium. Låt dess koncentration vara någonstans runt 150 millimol per liter. Mycket kalium. Låt oss sätta det inom parentes, eftersom parentesen står för koncentration. Det finns också lite kalium utanför. Här blir koncentrationen cirka 5 millimol per liter. Jag kommer att visa dig hur koncentrationsgradienten kommer att ställas in. Det händer inte av sig självt. Detta tar mycket energi. Två kaliumjoner pumpas in och samtidigt lämnar tre natriumjoner cellen. Så kaliumjonerna kommer in i början. Nu när de är inne, kommer de att hållas där själva? Självklart inte. De hittar anjoner, små molekyler eller atomer med en negativ laddning och ligger nära dem. Därmed blir nettoladdningen neutral. Varje katjon har sin egen anjon. Och vanligtvis är dessa anjoner proteiner, någon form av strukturer som har en negativ sidokedja. Det kan vara klorid eller till exempel fosfat. Något. Alla dessa anjoner kommer att fungera. Jag kommer att avbilda några fler anjoner. Så här är två kaliumjoner som precis kommit in i cellen, så här ser det ut nu. Om allt är bra och statiskt, så ser de ut så här. Och faktiskt, för att vara rättvis, finns det också små anjoner som är i nivå med kaliumjonerna här. Buren har små hål genom vilka kalium kan rinna ut. Låt oss se hur detta kommer att se ut och hur det kommer att påverka vad som händer här. Så vi har dessa små kanaler. Endast kalium kan passera genom dem. Det vill säga att dessa kanaler är mycket specifika för kalium. Inget annat kan passera genom dem. Inga anjoner, inga proteiner. Kaliumjonerna verkar leta efter dessa kanaler och resonerar: ”Wow, vad intressant! Det finns så mycket kalium här! Vi borde gå ut." Och alla dessa kaliumjoner lämnar bara cellen. Gå ut. Och som ett resultat händer en intressant sak. De flesta av dem har flyttat utåt. Men det finns redan några kaliumjoner utanför. Jag sa att den här lilla jonen var här, och att den teoretiskt sett kunde tränga in i den. Han kan gå in i den här buren om han vill. Men faktum är att totalt, totalt, har du fler rörelser utåt än inåt. Nu raderar jag denna väg eftersom jag vill att du ska komma ihåg att vi har fler kaliumjoner som tenderar att komma ut på grund av närvaron av en koncentrationsgradient. Detta är det första steget. Låt mig skriva ner det här. Koncentrationsgradienten gör att kaliumet rör sig utåt. Kalium börjar röra sig utåt. Lämnar buren. Och sen då? Kom igen, jag ska rita den när jag går ut. Denna kaliumjon är nu här, och den här är här. Bara anjoner finns kvar. De fanns kvar efter att kaliumet lämnat. Och dessa anjoner börjar producera en negativ laddning. En mycket stor negativ laddning. Endast ett fåtal anjoner som rör sig fram och tillbaka skapar en negativ laddning. Och kaliumjonerna på utsidan tycker att detta är väldigt intressant. Det finns en negativ laddning här. Och eftersom han är där, attraheras de av honom, eftersom de själva har en positiv laddning. De dras till en negativ laddning. De vill gå tillbaka. Tänk på det nu. Du har en koncentrationsgradient som pressar kaliumet utåt. Men å andra sidan finns det membranpotential, - i det här fallet, negativ - vilket uppstår på grund av att kalium lämnade efter sig en anjon. Denna potential stimulerar kalium att studsa tillbaka. En kraft, koncentration, trycker kaliumjonen utåt, en annan kraft, membranpotentialen, som skapas av kalium, gör att den återvänder inåt. Frigör lite utrymme. Nu ska jag visa dig något intressant. Låt oss konstruera två kurvor. Jag ska försöka att inte missa något på denna bild. Jag kommer att rita allt här och sedan kommer ett litet fragment av detta att synas. Vi bygger två kurvor. En av dem kommer att vara för koncentrationsgradienten och den andra för membranpotentialen. Dessa kommer att vara kaliumjonerna utanför. Om du följer dem över tid - den här gången - får du något i stil med följande. Kaliumjoner tenderar att komma ut och nå jämvikt vid en viss punkt. Låt oss göra samma sak med tiden på den här axeln. Detta kommer att vara vår membranpotential. Vi börjar vid nolltidpunkten och får ett negativt resultat. Den negativa laddningen kommer att bli större och större. Vi börjar vid nollpunkten för membranpotentialen, och det är vid den punkt där kaliumjoner börjar gå ut som följande händer. V översikt allt är väldigt likt, men det sker parallellt med förändringarna koncentrationsgrad... Och när dessa två värden är lika med varandra, när antalet kaliumjoner som lämnar utanför är lika med antalet kaliumjoner som återvänder inuti, är detta en sådan platå. Och det visar sig att laddningen är minus 92 millivolt. Vid denna punkt, där det praktiskt taget inte finns någon skillnad i termer av den totala rörelsen av kaliumjoner, observeras jämvikt. Den har till och med sitt eget namn - "jämviktspotentialen för kalium." När värdet når minus 92 - och det skiljer sig beroende på typen av joner - när man når minus 92 för kalium skapas en jämvikt av potentialer. Låt mig skriva ner att laddningen för kalium är minus 92. Detta händer bara när cellen är genomsläpplig endast för ett grundämne, till exempel för kaliumjoner. Och fortfarande kan en fråga uppstå. Du kanske tänker: "Okej, vänta en sekund! Om kaliumjoner rör sig utåt - vilket de är - har vi då inte en lägre koncentration vid en viss punkt, eftersom kalium redan har lämnat här, och en högre koncentration här tillhandahålls på grund av att kalium förflyttas utåt?" Tekniskt är det så. Här utanför finns det fler kaliumjoner. Och jag nämnde inte att volymen ändras också. En högre koncentration erhålls här. Och detsamma gäller för cellen. Det är tekniskt sett en lägre koncentration. Men jag har faktiskt inte ändrat värdena. Och anledningen är följande. Titta på dessa värden, de är nattfjärilar. Och det är ett stort antal, eller hur? 6,02 gånger 10 till minus 23 är inte ett litet tal alls. Och om du multiplicerar det med 5 får du något i stil med - låt mig snabbt räkna ut vad vi gjorde. 6 gånger 5 är 30. Och här är millimol. 10 till 20 mol. Det är enkelt stor mängd kaliumjoner. Och för att skapa en negativ laddning behövs väldigt få av dem. Det vill säga att förändringarna som orsakas av jonernas rörelse kommer att vara obetydliga jämfört med 10 till 20 grader. Det är därför som förändringar i koncentrationen inte beaktas.
Översikt
Den elektrokemiska potentialen används inom elektroanalytisk kemi och inom industrin används den vid tillverkning av batterier och bränsleceller. Den representerar en av de många utbytbara formerna av potentiell energi, i form av vilken energibesparing är möjlig.
I biologiska processer passerar joner genom membranet genom diffusion eller aktiv transport, bestämt av en elektrokemisk gradient. I mitokondrier och kloroplaster används protongradienter för att generera kemiosmotisk potential som också kallas proton- drivkraft Δp eller ΔμH+... Denna potentiella energi används för att syntetisera ATP genom eller fotofosforylering. Den protondrivande kraften, enligt Mitchells kemiosmotiska teori, är en vanlig produkt av de kopplade processerna av andning och oxidativ fosforylering. Den består av två faktorer: kemisk (eller osmotisk) - skillnaden i koncentrationerna av H + -joner i mitokondriella matrisen och intermembranutrymmet, och elektrisk - på grund av skillnaden i elektriska laddningar placerade på motsatta sidor av membranet. Skillnaden i koncentration av H+-joner, mätt i pH-enheter, betecknas med ΔpH. Den elektriska potentialskillnaden indikeras med symbolen Δψ. Därför tar ekvationen formen:
Δ μ H + = Δ ψ + Δ p H (\ displaystyle \ Delta \ mu _ (H ^ (+)) = \ Delta \ psi + \ Delta pH) ,
Δ p H = p H A - p H B (\ displaystyle \ Delta pH = pH_ (A) -pH_ (B))
skillnaden i koncentrationen av H+-joner (kemisk gradient) på A(+)-sidan och B(-)-sidan av membranet.
Den elektrokemiska gradienten liknar det tryck som vattnet utövar när det strömmar genom en vattenkraftsdamm. Membrantransportproteiner som natrium-kalium ATPas är analoga med turbiner, omvandlar den potentiella energin av vatten till andra former av fysisk eller kemisk energi, och jonerna som passerar genom membranet är analoga med vatten som faller till botten av en damm. Dessutom kan energin användas för att pumpa in vatten i en sjö uppströms dammen. Likaså kan kemisk energi i celler användas för att skapa elektrokemiska gradienter.
Kemi
Termen "elektrokemisk potential" används vanligtvis när en kemisk reaktion ska ske, till exempel med överföring av en elektron i ett elektriskt batteri. I batterier balanserar den elektrokemiska potentialen som uppstår från jonernas rörelse elektrodernas reaktionsenergi. Den maximala spänningen som ett batteri kan producera kallas standardelektrokemisk potential för den reaktionen. Tillsammans med högenergiföreningar kan kemisk energi lagras på biologiska membran, som fungerar som kondensatorer, som fungerar som ett isolerande skikt för laddade joner.
Biologisk betydelse
Generering av en transmembran elektrisk potential genom rörelse av joner genom cellmembranet leder till biologiska processer som nervledning, muskelsammandragning, hormonutsöndring och sensoriska svar. Man tror att membranet hos en typisk djurcell har en transmembranelektrisk potential från -50 mV till -70 mV.
Elektrokemiska gradienter spelar också en roll för att etablera protongradienter för oxidativ fosforylering i mitokondrier. Det sista stadiet av cellandning är elektrontransportkedjan. Fyra inbyggda komplex i det inre mitokondriella membranet (cristae) utgör elektrontransportkedjan. Men bara komplex I, III och IV är protonpumpar och pumpar protoner från matrisen in i intermembranutrymmet. Totalt erhålls tio protoner som rör sig från matrisen till intermembranutrymmet och genererar en elektrokemisk potential på mer än 200 mV. Detta sätter igång ett flöde av protoner tillbaka in i matrisen genom ATP-syntas, som syntetiserar ATP genom att fästa oorganiskt fosfat till ADP-molekylen. Sålunda är genereringen av en elektrokemisk protongradient kritisk för energisyntes i mitokondrier. Den allmänna ekvationen för elektrontransportkedjan ser ut så här:
NADH + 11 H + (matris) + 1/2 O 2 ⟶ NAD + + 10 H + (IMS) + H 2 O (\ displaystyle NADH + 11H ^ (+) (matris) +1/2 \ O_ (2) \ långhögerpil NAD ^ (+) + 10H ^ (+) (IMS) + H_ (2) O) .
Elektrontransportkedjan för fotosyntes i växter fungerar också som den respiratoriska kedjan för elektronöverföring, där protoner pumpas in i kloroplasternas lumen (tylakoidlumen), och den resulterande gradienten används för syntes av ATP genom enzymet ATP-syntas. Protongradienten kan genereras med användning av acyklisk eller cyklisk fotofosforylering. Proteiner som är involverade i icke-cyklisk fotofosforylering, fotosystem II (PSII) och cytokrom b6f-komplexet är direkt kapabla att generera en protongradient. För var och en av de fyra fotoner som absorberas av PSII finns det åtta protoner som pumpas in i lumen (tylakoidlumen) från stroma. Den allmänna ekvationen för fotofosforylering är följande:
2 H 2 O + 6 H + (stroma) + 2 NADP + ⟶ O 2 + 8 H + (lumen) + 2 NADPH (\ displaystyle 2H_ (2) O + 6H ^ (+) (stroma) + 2NADP ^ (+ ) \ långhögerpil O_ (2) + 8H ^ (+) (lumen) + 2NADPH) .
Flera andra transportörer och jonkanaler spelar en roll för att generera den elektrokemiska protongradienten. En av dem är TPK 3 -kaliumjonkanalen aktiverad av Ca 2+ joner. Den flyttar K+-joner från lumen till stroma, vilket hjälper till att etablera en pH-gradient (koncentrationsgradient) inom stroma. Å andra sidan transporterar den elektriskt neutrala antiportern K+ (KEA 3) K+-joner till lumen och H+ till stroma, upprätthåller balansen mellan joner och stör inte det elektriska fältet.
Jonisk gradient
Eftersom joner bär en laddning kan de inte passera genom membranet genom underlättad diffusion. Överföringen av joner över membranet är möjlig på två sätt, genom aktiv eller passiv transport. Ett exempel på aktiv transport av joner är arbetet med Na + -K + -ATPas. Det katalyserar reaktionen av ATP-hydrolys till ADP och oorganiskt fosfat Fn. Under hydrolysen av en ATP-molekyl frigörs energi, vilket förändrar enzymets konformation, så att tre Na+-joner transporteras ut och två K+-joner transporteras in i cellen. Som ett resultat blir innehållet i cellen mer negativt laddat än miljö, genereras elektrisk potential (EMF) Vm ≈ -60 mV. Ett exempel på passiv transport är strömmen av joner genom jonkanaler (kanaler för Na +, K +, Ca 2+ och Cl -) längs koncentrationsgradienten, från regionen med högre koncentration till regionen med lägre. Till exempel, eftersom det finns en hög koncentration av Na+ utanför cellen, tenderar Na+-joner att komma in i cellen genom natriumjonkanalen. Eftersom den elektriska potentialen inuti cellen är negativ, kommer inflödet av positiva joner att orsaka membrandepolarisering, som ett resultat av vilket värdet på den transmembrana elektriska potentialen skiftar närmare noll. Na +-joner kommer dock att fortsätta att röra sig nedåt i koncentrationsgradienten så länge som den kemiska gradientens drivkraft är större än den elektriska potentialen. Efter att effekten av båda gradienterna (kemiska och elektriska) balanserar varandra (V m för Na + är ca +70 mV), kommer inflödet av Na + joner att upphöra, eftersom drivkraften (ΔG) blir noll. Ekvationen för drivkraften är följande:
Δ G = R T l n (C i n / C e x t) + Z F V m (\ displaystyle \ Delta G = RTln (C_ (in) / C_ (ext)) + ZFV_ (m)).
Protongradienter
Protongradienter är väsentliga som en form av energilagring i många olika typer av celler. Gradienten används vanligtvis för att arbeta med ATP-syntas, rotation av flagellen eller transport av metaboliter över membranet. Detta avsnitt kommer att fokusera på tre processer som hjälper till att etablera protongradienter i respektive celler: arbetet med bakteriorodopsin, icke-cyklisk fotofosforylering och oxidativ fosforylering.
Bacteriorhodopsin
Bakteriodopsinet som finns i archaea bildar en väg för protongradienten via en protonpump. Protonpumpen förlitar sig på en protonbärare (rhodopsin) som rör sig från sidan av membranet med en låg koncentration av H+-joner till sidan med en högre koncentration av H+. Protonpumpen av bacteriorhodopsin aktiveras genom att absorbera fotoner med en våglängd på 568 nm, vilket leder till fotoisomerisering av Schiff-basen (SB) i näthinnan, vilket orsakar dess övergång från Trans- vid 13 cis-form. Fotoisomerisering är extremt snabb och tar bara 200 femtosekunder. Som en konsekvens genomgår rhodopsin ett antal snabba konformationella omarrangemang: Schiff-basen förskjuts från rester Asp85 och Asp212, vilket orsakar överföringen av H+-joner till resten Asp85, samtidigt bildas tillståndet Ml (meta-I). Sedan går proteinet till M2-tillståndet (meta-II) genom att separera resten Glu204 från Glu194, som släpper ut en proton i den yttre miljön. Detta tillstånd är relativt långlivat. Schiffbasen reprotoneras vid resten Asp85 bildar tillståndet N. Det är viktigt att den andra protonen kommer från Asp96 eftersom dess deprotonerade tillstånd är instabilt och snabbt reprotoneras (återprotoneras) av en proton från cytoplasman. Protonation Asp85 och Asp96 leda till upprepad isomerisering av SB, vilket bildar tillståndet O. Även i detta fall återstoden Asp85 släpper sin proton på Glu204 och bakteriohodopsin återgår till vila.
Fotofosforylering
Efter frisättning från PSII överförs den reducerade plastokinonen PQH 2 till cytokrom b6f-komplexet, som överför två elektroner från PQH 2 till plastocyanin y-proteinet i två separata reaktioner. Denna process liknar den Q-cykel som förekommer i komplex III i ETC. I den första reaktionen binder plastokinol PQH 2 till komplexet från sidan av lumen och en elektron passerar till järn-svavelcentrum (Fe-S), som sedan överför den till cytokrom f, den senare överför en elektron till plastocyanin molekyl. Den andra elektronen överförs till hemmolekylen bL, som sedan överför den till hemen bH, den senare överför elektronen till den andra plastokinonmolekylen PQ ,. I den andra reaktionen oxideras den andra plastokinolmolekylen PQH 2, vilket överför en elektron till en annan plastocyaninmolekyl och halvreducerad PQ, som reduceras till PQH 2 och lämnar komplexet. Båda reaktionerna åtföljs av överföring av fyra protoner per lumen.
Oxidativ fosforylering
NADH + H + + UQ + 4 H + (matris) ⟶ NAD + + UQH 2 + 4 H + (IMS) (\ displaystil NADH + H ^ (+) + UQ + 4H ^ (+) (matris) \ långhögerpil NAD ^ (+) + UQH_ (2) + 4H ^ (+) (IMS))
Anteckningar (redigera)
- Nelson, David. Lehninger Principles of Biochemistry / David Nelson, Michael Cox. - New York: W.H. Freeman, 2013. - S. 403. - ISBN 978-1-4292-3414-6.
- Nath, Sunil; Villadsen, John (2015-03-01). "Oxidativ fosforylering återupptas". Bioteknik och bioteknik [engelsk]. 112 (3): 429-437. DOI: 10.1002 / bit.25492. ISSN 1097-0290.
- Kolman J., Rem K.-G. Visuell biokemi. - M.: Mir, 2011 .-- S. 128-129. - 469 sid. - 7000 exemplar. - ISBN 5-03-003304-1.
- Stroyev E.A. Biologisk kemi. - M.: Vysshaya shkola, 1986 .-- S. 210 .-- 479 s.
- Yang, Huanghe; Zhang, Guohui; Cui, Jianmin (2015-01-01). "BK-kanaler: flera sensorer, en aktiveringsgrind". Membranfysiologi och membranbiofysik. 6 : 29. DOI: 10.3389 / fphys.2015.00029. PMC. PMID.
- Shattock, Michael J.; Ottolia, Michela; Bers, Donald M.; Blaustein, Mordecai P.; Boguslavskyi, Andrii; Bossuyt, Julie; Bridge, John H. B.; Chen-Izu, Ye; Clancy, Colleen E. (2015-03-15). "Na + / Ca2 + utbyte och Na + / K + -ATPas i hjärtat". Journal of Physiology [engelsk]. 593 (6): 1361-1382. DOI: 10.1113 / jphysiol.2014.282319. ISSN 1469-7793. PMC. PMID.
- Aperia, Anita; Akkuratov, Evgeny E.; Fontana, Jacopo Maria; Brismar, Hjalmar (2016-04-01). "Na + -K + -ATPas, en ny klass av plasmamembranreceptorer". American Journal of Physiology - Cell Physiology [engelsk]. 310 (7): C491-C495. DOI: 10.1152 / ajpcell.00359.2015. ISSN 0363-6143. PMID.
- Nelson, David. Lehninger Principles of Biochemistry / David Nelson, Michael Cox. - New York: W.H. Freeman, 2013. - S. 464. - ISBN 978-1-4292-3414-6.
- Poburko, Damon; Demaurex, Nicolas (2012-04-24). "Reglering av den mitokondriella protongradienten genom cytosoliska Ca2+-signaler". Pflügers Archiv - European Journal of Physiology [engelsk]. 464 (1): 19-26.
Elektrokemisk gradient jon är drivkraften för jonflöde, vilket är en kombination av membranpotential (elektrisk gradient) och koncentrationsgradient av ämnen (kemisk gradient). Den elektriska gradienten kännetecknar rörelsen av endast joner och är riktad mot deras motsatta laddning. Den kemiska gradienten är riktad från ett område med hög koncentration av lösta ämnen till ett område med låg koncentration.
Överföringen av ämnen genom membranet kan ske passivt och aktivt. Aktiv transport kräver energiförbrukning, och passiv genomförs utan energiförbrukning. Aktiv transport går alltid emot den elektrokemiska gradienten. Passiv transport lösta ämnen kan endast förekomma längs en gynnsam elektrokemisk gradient.
Transportsystem för lösta ämnen kan klassificeras baserat på användning cellulär energi.

1. Passiv transport kräver inte ATP-hydrolys och är inte associerad med överföringen av ett annat löst ämne.
Diffusion av fettlösliga ämnen (t.ex. O 2, CO 2, alkoholer och etrar) kan förekomma direkt genom plasmamembranet.
Transporten av joner och små molekyler sker ofta genom transmembranproteiner som tjänar jonkanaler(för olika joner) eller aquapores(för vattenmolekyler).
Joniska kanaler har följande vanliga komponenter:
1)porområdet, genom vilken jonerna diffunderar.
2)selektivt filter inuti poren, som ett resultat är kanalen mycket selektiv för vissa joner (t.ex. Na+-kanaler).
3) kanalporten, som öppnar och stänger kanalen I stängt tillstånd passerar inte joner genom kanalen utan kanalen är tillgänglig för aktivering. I öppet tillstånd rör sig joner enligt sin elektrokemiska gradient. Kanalporten kan styras av en av följande mekanismer: membranspänningar (spänningsstyrda kanaler); kemiska substanser (kemiskt beroende kanaler); mekaniska krafter i membranet (sträckberoende kanaler).
Diffusion kan uppstå och via bärarproteiner, kallad enhetlig, som selektivt binder ett löst ämne till ena sidan av membranet och genomgår konformationsförändringar för att leverera det till den andra sidan. Transporten av lösta ämnen genom uniporten kallas underlättad diffusion, eftersom det är snabbare än enkel diffusion. Det är så glukos och aminosyror transporteras.

Osmos - det är rörelsen (diffusionen) av vatten genom membranets vattenporer, som drivs av vattenkoncentrationsgradienten Vattenkoncentrationen uttrycks i termer av den totala koncentrationen av det lösta ämnet; ju mer utspädd lösningen är, desto lägre koncentration av det lösta ämnet och desto högre koncentration av vatten När två lösningar separeras semipermeabelt membran(vilket tillåter transport av vatten, men inte lösta ämnen) går vattnet från en mer utspädd lösning till en mer koncentrerad. Osmolaritetär ett uttryck för den osmotiska styrkan hos en lösning . Två lösningar med samma osmolaritet kallas isoosmotisk. Lösningar som är högre än referenslösningens osmolaritet kallas hyperosmotisk, och lösningar med lägre osmolaritet kallas hypoosmotisk. En isotonisk lösning har samma osmolaritet som fungerande celler och orsakar inte förflyttning av rent vatten genom deras membran; en hypoton lösning har lägre osmolaritet än en fungerande cell och får cellerna att svälla, en hyperton lösning har högre osmolaritet än celler och får cellerna att krympa. Till exempel om patienten injiceras intravenöst hypoton lösning, tonen i den extracellulära vätskan minskar initialt, och vatten flyttas in i den intracellulära vätskan genom osmos (celler sväller). Omvänt, om du går in hypertonisk lösning, ökar tonen i den extracellulära vätskan, och vatten kommer ut ur den intracellulära vätskan (cellerna krymper).
Medan det konstgjorda lipidmembranet är praktiskt taget ogenomträngligt för joner, innehåller biologiska membran " jonkanaler ", Genom vilken individuella joner selektivt penetrerar membranet (se). Membranpermeabiliteten och polariteten beror på elektrokemisk gradient det vill säga på koncentrationen av joner på båda sidor av membranet ( koncentrationsgrad) och från skillnader elektriska potentialer mellan membranets inre och yttre sidor ( membranpotential).
I ett tillstånd av vilande celler, membranpotentialen ( vilopotential, se) är från -0,05 till -0,09 V, det vill säga ett överskott av negativa laddningar råder på insidan av plasmamembranet. Vilopotentialen tillhandahålls i första hand av Na+- och K+-katjoner, såväl som av organiska anjoner och Cl-jonen (1). Koncentrationerna utanför och inuti cellen och permeabilitetskoefficienterna för dessa joner anges i tabell (2).
Fördelningen av joner mellan den yttre miljön och cellens inre volym beskrivs Nernst-ekvationen(3), där ΔΨ G är transmembranpotentialen (i volt, V), det vill säga skillnaden i elektriska potentialer mellan de två sidorna av membranet i frånvaro av jontransport genom membranet ( jämviktspotential). För envärda joner vid 25 ° C är RT / Fn-faktorn 0,026 V. Samtidigt följer det av tabell (2) att för K + joner är ΔΨ G ungefär lika med -0,09 V, dvs värdet är av samma storleksordning som och potential för vila. För Na + joner, tvärtom, ΔΨ G ≈ +0,07 V, det vill säga högre än vilopotentialen. Därför kommer Na+-joner in i cellen när Na+-kanalen öppnas. Ojämlikhet mellan Na + och K + jonkoncentrationer upprätthålls konstant Na+/K+-ATP-as när man spenderar ATP (se).
Artiklar i avsnittet "Energihushållning på membran":
- A. Elektrokemisk gradient
2012-2019. Visuell biokemi. Molekylärbiologi. Vitaminer och deras funktioner.
Uppslagsboken i visuell form - i form av färgscheman - beskriver alla biokemiska processer. Biokemiskt viktiga kemiska föreningar, deras struktur och egenskaper, huvudprocesserna med deras deltagande, samt mekanismerna och biokemin för de viktigaste processerna i levande natur beaktas. För studenter och lärare vid kemiska, biologiska och medicinska universitet, biokemister, biologer, läkare, samt alla som är intresserade av livsprocesser.