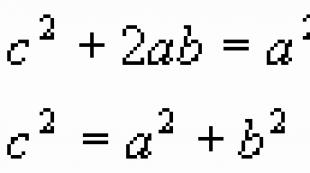Geno lokuso apibrėžimas. Dviejų genų lokusų ryšio nustatymas. Nuorodų aptikimas. Pažiūrėkite, kas yra „Locus“ kituose žodynuose
Kiaulių veisime Europoje ir Amerikoje pradedama taikyti genominė selekcija. Jo technologijos leidžia iššifruoti kiaulių genotipą jau atsivedus ir atrinkti geriausius gyvulius veisimui. Ši naujausia technologija skirta dar labiau padidinti kiaulių veisimo tikslumą ir veislinės vertės patikimumą.
Genominės atrankos protėvis yra žymenų atranka.
Žymeklių atranka – tai žymenų naudojimas kiekybinio požymio genams pažymėti, leidžiantis nustatyti tam tikrų genų (genų alelių) buvimą ar nebuvimą genome.
Genas – tai DNR dalis, tam tikra nukleotidų seka, kuri koduoja informaciją apie vienos baltymo molekulės (arba RNR) sintezę ir dėl to užtikrina bet kokio požymio susidarymą ir jo perdavimą paveldėjimo būdu.
Genai, atstovaujami populiacijoje kelių formų – alelių – yra polimorfiniai genai. Genų aleliai skirstomi į dominuojančius ir recesyvinius. Genų polimorfizmas suteikia įvairių rūšių bruožų.
Tačiau tik keli bruožai yra kontroliuojami atskirų genų (pavyzdžiui, plaukų spalva). Produktyvumo rodikliai, kaip taisyklė, yra kiekybiniai požymiai, už kurių vystymąsi ir pasireiškimą yra atsakingi daugelis genų. Kai kurie iš šių genų gali turėti ryškesnį poveikį. Tokie genai vadinami kiekybinių požymių lokusų (QTL) branduolių genais. Kiekybinio požymio lokusai (QTL) yra DNR segmentai, kuriuose yra genų arba kurie yra susieti su genais, kuriais grindžiamas kiekybinis požymis.
Pirmą kartą idėją naudoti žymenis veisime teoriškai pagrindė A. S. Serebrovskis dar 20-aisiais. Anot A. S. Serebrovskio, žymuo (tada vadintas „signal“, vėliau pradėtas vartoti angliškas terminas „marker“) yra geno alelis, turintis aiškiai išreikštą fenotipinę apraišką, lokalizuotas šalia kito alelio, lemiančio ekonomiškai svarbų. tiriamas bruožas, tačiau neturi aiškaus fenotipinio pasireiškimo; taigi, atliekant atranką dėl šio signalinio alelio fenotipinio pasireiškimo, atrenkami susietieji aleliai, lemiantys tiriamo požymio pasireiškimą.
Iš pradžių morfologiniai (fenotipiniai) požymiai buvo naudojami kaip genetiniai žymenys. Tačiau labai dažnai kiekybiniai požymiai paveldimi kompleksiškai, jų pasireiškimą lemia aplinkos sąlygos, o žymenų, kurie naudojami kaip fenotipiniai požymiai, skaičius yra ribotas. Tada genų produktai (baltymai) buvo naudojami kaip žymenys. Tačiau veiksmingiausias būdas patikrinti genetinį polimorfizmą yra ne genų produktų, o tiesiogiai genų lygyje, tai yra, naudojant polimorfines DNR nukleotidų sekas kaip žymenis.
Paprastai DNR fragmentai, esantys arti vienas kito chromosomoje, yra paveldimi kartu. Ši savybė leidžia naudoti žymeklį norint nustatyti tikslią geno, kuris dar nebuvo tiksliai nustatytas, paveldėjimo modelį.
Taigi, žymenys yra polimorfinės DNR sritys, turinčios žinomą vietą chromosomoje, bet nežinomos funkcijos, kurios gali būti naudojamos kitiems genams identifikuoti. Genetiniai žymenys turi būti lengvai atpažįstami, susieti su konkrečiu lokusu ir labai polimorfiški, nes homozigotos neteikia jokios informacijos.
Plačiai pradėti naudoti DNR polimorfizmo variantai, kaip genetiniai žymenys, 1980 m. Molekuliniai genetiniai žymenys buvo naudojami ūkinių gyvūnų veislių genofondų išsaugojimo programoms, buvo naudojami veislių kilmės ir paplitimo problemoms spręsti, giminystės nustatymui, kartografavimui. pagrindiniai kiekybinių požymių lokusai, tiriant genetines paveldimų ligų priežastis, atrankos paspartinimas pagal individualius požymius – atsparumas tam tikriems veiksniams, pagal produktyvumo rodiklius. Europoje genetiniai žymenys kiaulininkystėje naudojami nuo 1990-ųjų pradžios. išlaisvinti populiaciją nuo halotano geno, sukeliančio kiaulėms streso sindromą.
Yra keletas molekulinių genetinių žymenų tipų. Dar visai neseniai mikropalydovai buvo labai populiarūs, nes jie yra plačiai paplitę genome ir turi aukštas lygis polimorfizmas. Mikropalydovai – SSR (Simple Sequence Repeats) arba STR (Simple Tandem Repeats) susideda iš 2-6 bazinių porų ilgio DNR segmentų, kartojamų daug kartų tandemu. Pavyzdžiui, amerikiečių kompanija Applied Biosystems sukūrė 11 mikropalydovų (TGLA227, BM2113, TGLA53, ETH10, SPS115, TGLA126, TGLA122, INRA23, ETH3, ETH225, BM1824) genotipų nustatymo testavimo sistemą. Tačiau mikropalydovų nepakanka norint tiksliai nustatyti atskirų genomų regionų žemėlapius, didelės įrangos ir reagentų kainos bei automatizuotų metodų, naudojant SNP lustus, kūrimas verčia juos atsisakyti praktikos.
Labai patogus genetinių žymenų tipas yra SNP (Single Nucleotide Polymorphisms) – snip arba vieno nukleotido polimorfizmas– tai vieno nukleotido DNR sekos skirtumai tos pačios rūšies atstovų genome arba tarp homologinių individo homologinių chromosomų sričių. SNP yra taškinės mutacijos, kurios gali atsirasti dėl spontaniškų mutacijų ir mutagenų veikimo. Net vienos bazių poros skirtumas gali sukelti bruožo pasikeitimą. SNP yra plačiai paplitę genome (žmonės turi apie 1 SNP 1000 bazinių porų). Kiaulių genomas turi milijonus taškinių mutacijų. Joks kitas genomo skirtumas negali užtikrinti tokio žymenų tankio. Be to, SNP turi žemą mutacijų lygį per kartą (~ 10–8), priešingai nei mikrosatelitai, todėl jie yra patogūs populiacijos genetinės analizės žymekliai. Pagrindinis SNP pranašumas yra galimybė naudoti automatinius jų aptikimo metodus, pavyzdžiui, DNR šablonų naudojimą.
Siekdamos padidinti SNP žymenų skaičių, nemažai užsienio kompanijų neseniai suvienijo jėgas, kad sukurtų vieną duomenų bazę, kad būtų galima išbandyti daugybę gyvūnų, kurių produktyvumas buvo išbandytas dėl polimorfizmo, ir nustatyti ryšių tarp žinomo taško buvimą. mutacijos ir produktyvumas.
Šiuo metu nustatyta labai daug polimorfinių genų variantų ir jų tarpusavio įtakos kiaulių produktyvumo požymiams. Kai kurie genetiniai testai, naudojant efektyvumo žymenis, yra viešai prieinami ir naudojami veisimo programose. Naudodami tokius žymeklius galite pagerinti kai kuriuos produktyvumo rodiklius.
Produktyvumo žymeklių pavyzdžiai:
- vaisingumo žymenys: ESR, estrogenų receptoriaus genas, EPOR, eritropoetino receptoriaus genas;
- atsparumo ligoms žymenys – ECR F18 receptoriaus genas;
- augimo efektyvumo, mėsos produktyvumo žymenys - MC4R, HMGA1, CCKAR, POU1F1.
MC4R – kiaulių melanokortino 4 receptoriaus genas yra 1 chromosomoje (SSC1) q22-q27. Vieną nukleotidą A pakeitus G, pasikeičia MC4 receptoriaus aminorūgščių sudėtis. Dėl to pažeidžiamas riebalinio audinio ląstelių sekrecijos reguliavimas, dėl kurio sutrinka lipidų apykaita ir tiesiogiai veikia požymių, apibūdinančių kiaulių penėjimo ir mėsos savybes, susidarymo procesą. A alelis lemia greitą augimą ir didelį šoninės storį, o G alelis atsakingas už augimo efektyvumą ir didelį liesos mėsos procentą. Homozigotinės kiaulės, turinčios AA genotipą, pasiekia rinkos svorį trimis dienomis greičiau nei homozigotinės pagal G alelį (GG), tačiau kiaulės, turinčios GG genotipą, turi 8% mažiau riebalų ir turi didesnę pašaro konversiją.
Kiti genai, valdantys susijusių fiziologinių procesų kompleksą, taip pat turi įtakos mėsos ir penėjimo produktyvumui. POU1F1 genas, hipofizės transkripcijos faktorius, yra reguliuojantis transkripcijos faktorius, lemiantis augimo hormono ir prolaktino ekspresiją. Kiaulių POU1F1 lokusas yra susietas su 13 chromosoma. Jo polimorfizmą lemia taškinė mutacija, dėl kurios susidaro du aleliai – C ir D. C alelio buvimas kiaulių genotipe yra susijęs su padidėjusiu vidutiniu paros svoriu. pelnas ir didesnis ankstyvumas.
Žymekliai taip pat leidžia patikrinti šerno genotipą dėl ribojamų lyties požymių, atsirandančių tik paršavedėms. Tai, pavyzdžiui, vaisingumas (paršelių skaičius per paršiuką), kurį šernas perduoda palikuonims. Pavyzdžiui, ištyrus šerno genotipą dėl estrogenų receptorių (ESR) žymenų, bus galima atrinkti veisimui tuos šernus, kurie savo dukterims perduos aukštesnes reprodukcines savybes.
Naudojant žymenų atrankos rezultatus, galima įvertinti pageidaujamų ir nepageidaujamų veislei ar linijai alelių atsiradimo dažnį, o tolesnė selekcija turėtų būti atliekama taip, kad visi veislės gyvūnai turėtų tik pageidaujamus genų alelius.

Ryžiai. 1. Oligonukleotidų biolusto veikimo principas
DNR lustas yra substratas su ląstelėmis, ant kurių nusėda reagentinė medžiaga. Tiriama medžiaga pažymima įvairiomis etiketėmis (dažnai fluorescenciniais dažais) ir uždedama ant biočipo. Kaip parodyta paveikslėlyje, reagento medžiaga – oligonukleotidas – jungiasi tiriamojoje medžiagoje – fluorescenciniu būdu pažymėti DNR fragmentai – tik komplementarus fragmentas. Dėl to ant šio biolusto elemento pastebimas švytėjimas.
2009 metais kiaulės genomas buvo iššifruotas. Sukurtas SNP lustas ( DNR mikroschemos variantas), kuriame yra 60 000 genetinių genomo žymenų. Siekiant paspartinti tyrimus, netgi buvo sukurti specialūs robotai, kurie nuskaito skilteles. Kiaulės DNR mėginys gali būti ištirtas, ar nėra beveik visų svarbių taškinių mutacijų, kurios lemia produktyvumo požymius. Taigi geriausių gyvūnų atranka gali būti pagrįsta genetiniais žymenimis, nematuojant fenotipinių rodiklių.
Dėl šios pažangos buvo įdiegta nauja technologija – genominė atranka. Genominė atranka – tai genomo iš karto tikrinimas dėl daugybės žymenų, apimančių visą genomą, kad kiekybiniai bruožų lokusai (QTL) būtų nesubalansuoti su bent vienu žymeniu. Genominio veisimo metu genomo nuskaitymas atliekamas naudojant lustus (matricas) su 50-60 tūkstančių SNP (kurie žymi pagrindinius kiekybinių požymių genus), siekiant nustatyti vieno nukleotido polimorfizmus išilgai gyvūno genomo, nustatyti genotipus su norimu produktyvių požymių rinkinio pasireiškimu. , ir įvertinti gyvūno veislinę vertę.
Terminą „genominė atranka“ Haley ir Wisher pirmą kartą įvedė 1998 m. Meuwissen ir kt., 2001 m., sukūrė ir pristatė veislinės vertės analitinio vertinimo metodiką naudojant genomo masto žymenų žemėlapį.
Praktiškai genominė atranka pradėta taikyti 2009 m.
Nuo 2009 metų didžiausios JAV (Cooperative Resources International), Nyderlandų, Vokietijos ir Australijos įmonės pradėjo diegti genominę selekciją į galvijų auginimo programas. Įvairių veislių bulių genotipas buvo nustatytas daugiau nei 50 000 SNP.
„Hypor“ pirmasis paskelbė visą rinkos genominio veisimo programą, kuris padidins veisimo tikslumą kiaulininkystėje. 2012 m. birželį žiniasklaidoje buvo paskelbta, kad „Hypor“ savo klientams gali pasiūlyti Genomic Breeding Value atsargų.

Genetikos įmonė „Hypor“ genominę atranką taiko nuo 2010 m., glaudžiai bendradarbiaudama su centru moksliniai tyrimai ir naujos Hendrix Genetics grupės technologijos (Hendrix Genetics). Hendrix Genetics išbando daugiau nei 60 000 SNP žymenų ir naudoja šią informaciją DNR tyrimams. Kiaulių genetinio potencialo genominis indeksas apskaičiuojamas išanalizavus 60 000 gyvūno genų žymenų (snip). Teoriškai, jei yra pakankamai genetinių žymenų, kad apimtų visą kiaulės DNR (jos genomą), galima apibūdinti visas genetines variacijas visoms išmatuojamoms savybėms. Rengiama moderni matematinė-genetinė duomenų apdorojimo programinė įranga.
Genetikos įmonė „Hendrix Genetics“ turi didelį biobanką – jame saugomi kelių ūkių ir kartų veislinių gyvūnų kraujo ir audinių mėginiai DNR tyrimams (gyvūnų genetinės vertės nustatymui) ir gyvūno genotipo analizei. Hypor daugiau nei dvejus metus atlieka kiaulių DNR tyrimus savo veisimo patalpose. Visi mėginiai iš skirtingų veisimo gamyklų, esančių skirtingose šalyse, siunčiami perdirbti į naują Hendrix Genetics centrinę genomo laboratoriją Ploufragant mieste (Prancūzija). Gerardas Albersas, Tyrimų ir naujųjų technologijų centro direktorius, pabrėžia: „Genominė laboratorija yra vertingas turtas, kuriuo dalijasi visos genetikos įmonės, sudarančios Hendrix Genetics, ir yra tikrai unikali kiaulių pramonėje“.
Genominė atranka yra galinga priemonė naudoti ateityje. Šiuo metu genominės atrankos efektyvumą riboja skirtingas kiekybinių požymių lokusų sąveikos pobūdis, skirtingų veislių kiekybinių požymių kintamumas ir aplinkos veiksnių įtaka požymio pasireiškimui. Tačiau daugelio šalių tyrimų rezultatai patvirtino, kad statistinių metodų naudojimas kartu su genominiu skenavimu padidina veislinės vertės prognozavimo patikimumą.
Kiaulių atranka statistiniais metodais kai kuriems rodikliams (pavyzdžiui, atsparumas ligoms, mėsos kokybė, vaisingumas) pasižymi mažu efektyvumu. Tai atsitinka dėl šių veiksnių:
- mažas savybių paveldimumas,
- didelę įtaką šiam aplinkos veiksnių ženklui,
- dėl pasireiškimo pagal lytį,
- bruožo pasireiškimas tik veikiant tam tikriems veiksniams,
- kai simptomas pasireiškia palyginti vėlai,
- dėl to, kad charakteristikas sunku išmatuoti (pavyzdžiui, sveikatos charakteristikos),
- paslėptų nešėjų-ženklų buvimas.
Pavyzdžiui, toks kiaulių trūkumas kaip jautrumas stresui yra sunkiai diagnozuojamas ir pasireiškia padidėjusiu paršelių mirtingumu streso (transportavimo ir kt.) įtakoje bei mėsos kokybės pablogėjimu. DNR tyrimas naudojant genų žymenis leidžia nustatyti visus šio defekto nešiotojus, taip pat ir latentinius, ir atsižvelgiant į tai atlikti atranką.
Norint įvertinti sunkiai statistiniais metodais nuspėjamus produktyvumo rodiklius, patikimesniam jų įvertinimui reikalinga palikuonių analizė, tai yra reikia sulaukti palikuonių ir išanalizuoti jo veislinę vertę. O DNR žymenų naudojimas leidžia analizuoti genotipą iškart gimus, nelaukiant, kol pasireikš bruožas ar pasirodys palikuonys, o tai žymiai pagreitina atranką.
Gyvulių indeksinis vertinimas atliekamas pagal eksterjero ir produktyvumo savybes (paršelių ankstyva branda ir kt.). Abiem atvejais naudojami fenotipiniai rodikliai, todėl norint šiuos požymius panaudoti skaičiavimuose, būtina žinoti jų paveldimumo koeficientą. Tačiau net ir šiuo atveju sprendžiame bet kurio požymio genetinio pagrindimo tikimybę, jo protėvių ir palikuonių vidutinius rodiklius (nėra galimybės nustatyti, kokius genus jaunas gyvūnas paveldėjo: geresnius ar blogesnius už šį vidurkį). ). Genotipo analizės pagalba galima tiksliai nustatyti tam tikrų genų paveldėjimo faktą jau gimus, genotipus įvertinti tiesiogiai, o ne per fenotipines apraiškas.
Tačiau jei kiaulės atrenkamos dėl labai paveldimų savybių, pavyzdžiui, lengvai kiekybiškai įvertinamo spenių skaičiaus, genomo atranka didelės naudos neduos.
Žymeklių atranka nepaneigia tradicinių veislinės vertės nustatymo metodų. Statistinė analizė ir genominės atrankos technologijos papildo viena kitą. Genetinių žymenų naudojimas leidžia pagreitinti gyvūnų atrankos procesą, o indeksavimo metodai – tiksliau įvertinti šios atrankos efektyvumą.
Genominis veisimas yra galimybė kiaulininkystę paversti tikslia produkcija. Genominės atrankos technologijų panaudojimas leis gaminti įvairius mėsos gaminius, atitinkančius vartotojų poreikius.
Perpublikuoti medžiagą iš šios svetainės leidžiama tik tuo atveju, jei nurodote hipersaitą į informacijos šaltinį!
PASKAITA 1. Klasikinė ir molekulinė genetika. Pagrindinės sąvokos: požymis, fenotipas, genotipas, genas, lokusas, alelis, homozigotas, heterozigotas, hemizigotas.
ICG SB RAS ir FEN NSU, Novosibirskas, 2012 m
1.1. Klasikinė ir molekulinė genetika
Šios dienos paskaita įžanginė, prie specifikos pereisime vėliau. Kaip ir beveik bet kurio mokslo atveju, gana sunku nubrėžti genetikos ribas, o apibrėžimas yra labai bendras. genetika – paveldimumo mokslas“ nėra ypač vaisingas. Pavyzdžiui, Žimulevas kažkada yra sakęs, kad dabar genetika yra visur – medicinoje, kriminalistikoje, evoliucijos teorijoje, archeologijoje, o pačioje genetikoje net nukleino rūgštys yra beveik nematomos – visiškai baltymų sąveika. Taigi jis iš tikrųjų padėjo lygybės ženklą tarp genetikos ir visos šiuolaikinės biologijos. Kita vertus, maždaug pirmuosius du XX amžiaus trečdalius genetika buvo bene labiausiai izoliuota ir aiškiai apibrėžta biologijos sritis, kuri pirmiausia išsiskyrė sintetine metodika, priešingai nei kitų biologijos šakų analitinė metodika. Siekdama išsiaiškinti savo objekto sandarą, ji nedalijo jo į dalis, o apie dalis vertino netiesiogiai, stebėdama visumą (būtent stebėdama ženklų elgesį perėjose) ir remdamasi matematika ir įsitikino. apie jos išvadų teisingumą, gaudama gyvus organizmus su numatomomis savybėmis. Taigi genetika nuo pat pradžių turėjo galimybę sukurti kažką naujo, o ne tik aprašyti stebimą. Tuo pat metu XX amžiaus antroje pusėje sparčiai vystėsi molekulinė biologija – iš pradžių grynai analitinis mokslas, suskaidytas į dalis. Tačiau jo pažanga daugiausia buvo atlikta naudojant genetinius metodus – pavyzdžiui, atminkite, kad genetinis kodas buvo nustatytas Benzerio ir Cricko eksperimentuose, naudojant bakteriofagų mutacijas. Tačiau šiuo atveju buvo pasitelkta mikroorganizmų genetika, o „klasikinės“ genetikos pažanga visada buvo siejama su eukariotų genetika.
Dėl to molekulinė biologija gavo beveik išsamias žinias apie tai, kas ir kaip yra išdėstytas gyvas organizmas. Molekulinės biologijos ir genetikos dalykai daugeliu atžvilgių sutapo: abu tyrė paveldimos informacijos perdavimą ir įgyvendinimą (o gyvas organizmas yra paveldimos informacijos realizavimas), tačiau perėjo prie šio dalyko supratimo iš priešingų pusių – genetikos “. iš išorės“, molekulinė biologija „iš vidaus“.
Paskutiniame dvidešimtojo amžiaus trečdalyje, galima sakyti, susitiko molekulinė biologija ir genetika, įskaitant eukariotų tyrimą. Spekuliaciniai genetikos objektai virto visiškai specifiniais žinomos struktūros fiziniais ir cheminiais objektais, o molekulinė biologija tapo sintetiniu mokslu, galinčiu savo nuožiūra paveikti net aukštesnius daugialąsčius organizmus – pavyzdžiui, genetinę modifikaciją. Čia genetikos, kaip mokslo, ribos buvo ištrintos iki neatskiriamų – tapo neįmanoma pasakyti, kur baigiasi molekulinė biologija ir prasideda genetika. Be to, terminas „molekulinė genetika“ reiškia susidariusį sintetinį mokslą, todėl tapo neaišku, kas tiksliai liko genetikoje už pastarojo ribų. Ikimolekulinio laikotarpio genetikai su visais kryžiais ir tikimybių teorija paremtais požiūriais suteiktas „klasikinės genetikos“ garbės vardas. Kita vertus, su šiuo titulu ji tarsi buvo išsiųsta į garbingą pensiją. Galima prisiminti, kaip Watsonas ir Crickas atsisakė aptarti savo DNR struktūros modelį savo dokumente „Nature“, nes pasekmės buvo per didelės ir akivaizdžios. Tam tikru momentu gali atrodyti, kad visa genetika išplaukia iš šio modelio.
Susidaro paradoksali situacija. Visi genetikos kursai prasideda šio mokslo istorija. Ji supranta, kaip Mendelis dirbo su žirniais, ką gavo ir kaip tai interpretavo remdamasis savo žiniomis, tada kaip Morganas ir jo mokykla dirbo su Drosophila, ką gavo ir kaip tai interpretavo. Neįmanoma praleisti abiejų šių temų – Mendelis yra pavyzdys žmogaus, kuris vystėsi nuo nulio ir puikiai pritaikė matematika pagrįstą genetinę metodiką, o Morgano mokykla sukūrė chromosomų paveldimumo teoriją ir, tiesą sakant, visą klasikinę genetiką m. pirmieji trys dvidešimtojo amžiaus dešimtmečiai. Be to, genetikos kursus galima suskirstyti į dvi plačias klases. Kai kurie detaliai apdirba visą šio mokslo raidos istoriją ir vidinę logiką, parodydami ir jo metodologijos galią, ir žmogaus proto galimybes spekuliaciniu būdu skverbtis į daiktų gelmes. Kiti kursai, greitai praleidę šį istorinį laikotarpį, pereina prie molekulinės genetikos ir ten svarsto, kas šiuo metu žinoma apie genų struktūrą ir darbą. Tiesą sakant, abiejų tipų kursai klasikinę genetiką laiko praeityje ir skiriasi tik retrospektyvos detalėmis. Pasirodo, klasikinė genetika turi tarsi tik istorinę reikšmę. Tačiau jos galinga metodika niekur nedingo ir yra būtina labai įvairioms studijoms. Jei pažvelgsime į straipsnius gana molekuliniais biologiniais pavadinimais ir publikuotus geriausiuose žurnaluose, pamatysime, kad jie visi yra pagrįsti plačia medžiaga apie šimtus atskirų mutacijų ir jų derinių, atsižvelgiant į ryšį tarp mutacijų prigimties ir fenotipo. kad jie sukelia. Tai pasakytina ir apie Drosophila ar peles, kurioms surinktos didžiulės genetinės kolekcijos ir sukurtos specialios laboratorinės linijos (vienos maždaug prieš šimtą metų, kitos neseniai), tiek žmonėms, kur puiki suma medicininiai genetiniai – iš tikrųjų populiaciniai genetiniai – duomenys, susiję su paveldimomis ligomis. Ir kuo turtingesnis šis žinių ir pavyzdinių organizmų arsenalas, tuo darbas elegantiškesnis. Visi šie daugiau nei rimti tyrimai neįmanomi be klasikinės ir molekulinės genetikos metodologijos įvaldymo vienu metu. Todėl geriausia šias „dvi genetikas“ tirti lygiagrečiai, kad ir kaip sunku būtų organizuoti.
Šiuolaikiniame moksle galima pastebėti ir pavyzdžių, kaip „pasenusios“ klasikinės genetikos nepaisymas sukelia kuriozų. Pavyzdžiui, grupei Europos mokslininkų reikėjo gauti heterozigotą, skirtą perkėlimui į žirnį. (Dabar kalbu remdamasis tuo, kad jūs turite tam tikrą supratimą apie tai, kas yra pavojuje. Jei to neturite, nesvarbu, mes visa tai apsvarstysime beveik per daug išsamiai; kol kas mes kalbame apie genetinių žinių poreikį). Jie tai gavo susiliejus tėvų linijų protoplastams. Regeneracija iš žirnelių ląstelių kultūros yra nepaprastai sunki, tai nepaprastai daug pastangų reikalaujantis kelias. Kodėl jie tai padarė? Matyt, jie manė, kad translokacijos nešiotojai su paprastais žirniais nekryžmina! Tiesą sakant, problemų su reprodukcija kryžminant tėvus, kurių perkėlimas skiriasi, iškyla, tačiau tik kitos kartos metu ir prarandama tik pusė vaisingumo.
Tačiau šiems mokslininkams reikėjo bent jau heterozigoto. Tuo tarpu bendras susižavėjimas molekuline biologija ir klasikinės genetikos nepaisymas veda prie to, kad heterozigotų egzistavimas – tai yra, kad eukariotuose kiekvienas genas yra pavaizduotas dviem kopijomis, kurios gali skirtis arba gali būti identiškos – dažnai visiškai pamirštamas. . Pavyzdžiui, į mano apžvalgą atėjo vokiečių autorių straipsnis, kuriame jie tiesiogiai perskaitė tam tikrą nekoduojančią DNR seką iš 38 laumžirgių individų, sugautų skirtinguose regionuose (Vakarų Europoje, Vakarų Sibire, Japonijoje ir Šiaurės Amerikoje) ir rado 20 jų variantų. tai. Buvo parašyta taip, lyg kiekviename individe būtų rastas tik vienas variantas. Tačiau jei kintamumas iš tiesų yra toks didelis, kaip jie teigia, tada tikimybė, kad jų imtyje yra bent vienas asmuo, kurio abi šios sekos kopijos yra vienodos, nelabai skiriasi nuo nulio. Ir apie tai net nebuvo diskutuota. Po peržiūros jie parašė, kad penkiais atvejais kilo įtarimas dėl heterozigotiškumo. Jei iš tikrųjų jų yra tik penki, tada jų rankose buvo nuostabus reiškinys, kai heterozigotai virsta homozigotais mechanizmais, kurie iki šiol nežinomi, bet atrodė, kad jie to net nesuprato.
Dabar plačiai paplitusios filogeninės rekonstrukcijos, pagrįstos tam tikromis DNR sekomis. Taigi, gana dažnai pagal populiacijų išsiskyrimo laiką bandoma spręsti, ar šios populiacijos priklauso tai pačiai biologinei rūšiai, ar skirtingoms. (Atkreipkite dėmesį, kad įvertinamas skirtumo laikas, nes tiriami genai, kurių kintamumas laikui bėgant yra daugiau ar mažiau pastovus, akivaizdžiai nėra tie genai, kurių pasikeitimas gali būti susijęs su specifika). Tuo tarpu divergencijos laikas paprastai turi mažai ką bendro su šia problema – tam tikros vietos populiacijos reprodukcinės izoliacijos momentai, ty rūšiavimo momentai, atsiranda tam tikromis sąlygomis ir paprastai neužima daug laiko nuo paleontologinio tyrimo. požiūrio (nuo dešimtys iki šimtų tūkstančių metų), tada kaip populiacijos gali skirtis ilgą laiką be specifikacijos. Klausimas yra būtent išsiaiškinti, ar tarp populiacijų egzistuoja reprodukcinė izoliacija (bent jau potenciali). Norėdami tai padaryti, turėtumėte pamatyti, ar tarp jų vyksta genų mainai (jei tai fiziškai įmanoma), ar ne. Čia tiesiog labai svarbu išsiaiškinti, ar populiacijų sandūroje yra heterozigotų pagal kiekvienai iš jų būdingus alelius ir koks jų dažnis. Tačiau beveik niekas to nedaro, o tai, ar populiacijos priklauso tai pačiai, ar skirtingoms rūšims, sprendžiama pagal jų skirtumų lygį, lyginant juos su skirtumais tais atvejais, kurie, kaip manoma, yra neabejotini.
Apskritai, jei vieną organizmą (pavyzdžiui, kaip jo rūšies atstovą) galima tirti naudojant molekulinės genetikos metodus, tada, kai tik kalbama apie daugybę organizmų, tai yra, iškyla populiacijos genetinė problema – ir tokia. problema gana dažnai iškyla, pavyzdžiui, populiacijos biologijoje ir veislininkystėje – neapsieinama be klasikinės genetikos požiūrių. Klasikinė genetika yra būtina visame, kas susiję su individualiais skirtumais ir daugelio tos pačios rūšies individų savybėmis. Būtent tai yra jos elementas, ir kaip tik jame tie dabartiniai mokslininkai, kurie klasikinį genetinį išsilavinimą pakeitė molekuliniu biologiniu išsilavinimu, dažnai atsiduria bejėgiai.
Remdamasis tuo, kas išdėstyta, aš matau savo uždavinį pristatyti klasikinę genetiką ne tiek istoriniu aspektu, sekant didžiuosius praeities mokslininkus, o pradedant nuo dabartinės mokslo padėties, neatsiejant nuo jau turimų žinių. gautas molekulinės biologijos kursuose.ir citologijos. Tuo pačiu metu kai kurie modeliai, atrasti kaip grynai empiriniai organizmų lygmeniu, įgyja visiškai natūralų aiškinimą molekuliniu lygmeniu ir atrodo beveik trivialiai. Tuo pačiu metu šie dėsningumai turėtų būti aiškiai suprasti, nes jie turėtų būti naudojami organizmų lygmeniu. Tam tikra prasme toks genetikos kursas suvokiamas kaip kažkas panašaus į „gudrybių demonstravimą su apreiškimais“ – kur ir pats triukas, ir jo pagrindas yra vienodai „medicininiai faktai“. Toks kursas būtų skirtas išmokyti labai produktyvios metodikos: pereiti nuo bruožo prie genų ir, perpratus jų veikimo mechanizmą, vėl kopti į naujų bruožų sintezę.
Kaip jau supratote, šiuo metu genetikos turinys yra didžiulis ir nevienalytis, todėl mums skirto laiko vargu ar užtenka net trumpam įvadui. Tai verčia užkulisiuose palikti genetikos istoriją kaip savarankišką temą, kuriai turėjo būti skirtas specialus kursas.
Deja, aukščiau aprašytas genetikos studijų idealas dabartiniame etape – nuo bruožo iki geno ir atgal – neatitinka jokio turimo vadovėlio, greičiausiai dėl to, kad šis mokslas dabar vystosi per greitai. Kaip kompensaciją už šią aplinkybę pasistengsiu savo kuklias paskaitas patalpinti savo tinklalapyje, kur su jomis galės susipažinti tie, kuriems duosiu adresą – tai yra jums. Rekomenduočiau remtis vadovėliu - Vechtomovo „Genetika su atrankos pagrindais“. Taip pat gerai žinomas akademiko Igorio Fedorovičiaus Žimulevo vadovėlis „Bendroji ir molekulinė genetika“, kuriame pagrindinis akcentas – molekulinė genetika, o Leonidas Vladimirovičius rekomenduoja jį kaip pagrindinį vadovėlį. Suprantu, kad du pagrindiniai vadovėliai nėra pati patogiausia situacija išlaikyti egzaminą. Tačiau tai prisideda prie dalyko supratimo. Galiu pasakyti, kad aš asmeniškai čia ir apskritai dirbu Citologijos ir genetikos institute vien todėl, kad išklausiau Vladimiro Aleksandrovičiaus Berdnikovo genetikos kursą. Tai buvo geriausias mano girdėtas kursas ir visiškai neatitiko jokio vadovėlio, nes V. A. jį parengė remdamasis naujausiomis recenzijomis mokslinėje periodinėje spaudoje, kurios dar nebuvo įtrauktos į jokius vadovėlius. Igoris Fedorovičius savo originalų paskaitų kursą taip pat pavertė vadovėliu.
Labai nuodugniai paliesime genetikos pagrindus, kad juos gerai pajustume. Pradėsime nuo pat pradžių, nepaisant to, kad mokykloje yra padengiami elementariausi genetikos pagrindai, kad, neduok Dieve, neprarastume kažko paprasto, bet svarbaus. Kita vertus, aš bendrauju su tetos studentėmis, kurios jau yra išklausiusios molekulinės biologijos kursą ir šiuo metu studijuoja tikimybių teoriją ir matematinę statistiką, o tai leidžia man per daug nesiblaškyti dėl šių kursų medžiagos, kuri yra tokia reikalinga. genetikos studijoms. Pavyzdžiui, manysiu, kad žinote (arba žinosite tinkamu laiku), kas yra alternatyvus sujungimas arba Puasono paskirstymas.
Standartinė biologijos pristatymo universitetų kursuose logika yra pereiti iš apačios į viršų, nuo atomų prie molekulių ir makromolekulių, tada prie ląstelės struktūrų, prie pačios ląstelės gyvybės ir tada prie daugialąsčio organizmo. Kai žinome gyvenimo organizavimo principus iki galo, tokia pateikimo tvarka pasirodo organiška ir natūrali. Šie principai taip pat apima nukleorūgščių, kaip informacijos nešėjų, veikimo mechanizmą, pirmiausia apie įvairius baltymus ir funkcines RNR (kurios, atradus mažas RNR, pasirodė įvairesnės, nei manyta anksčiau), ne tik apie jų struktūrą, bet ir apie tai, kada, kur ir kiek tam tikrų RNR ar baltymų turėtų būti susintetinta. Šių procesų kontrolė vėlgi atliekama tam tikrų baltymų (o dažnai ir RNR) pagalba. Genetinės kontrolės sistemos vystosi kaskados principu – genai koduoja baltymus (RNR), reikalingus kontroliuoti genus, koduojančius kitus baltymus (RNR) ir kt. Kadangi beveik viską organizme „gamina“ baltymai (taip pat kai kurie RNR), paaiškėja, kad iš tikrųjų informacija apie visą organizmą yra įrašyta į nukleino rūgštis – tačiau šios informacijos nuskaityti neįmanoma be anksčiau susintetintų (vėlgi pagal DNR matricą) baltymų, kurie veikia DNR.
Tokia pateikimo tvarka visiškai sutampa su tvarka, kuria vystėsi pats gyvenimas. Iš pradžių tai buvo kažkokios „paprastos“ (bet tik palyginus su tuo, kas vėliau iš jų atsirado) savaime besidauginančių makromolekulių, matyt, nukleino rūgščių sistemos. Tada jie atsitiktinai apsupo save fosfolipidine membrana, kuri leido jiems sukurti savo mikrokosmosą. Taip gimė ląstelės. Baltymai vaidino vis svarbesnį vaidmenį šių pirmųjų gyvų būtybių veikloje, tačiau nukleorūgštys išlaikė visišką kontrolę. Ląstelės tapo sudėtingesnės ir išmoko vis teisingiau dalytis. Po pasidalijimo jie kartais neišsiskirstė, sudarydami kolonijas. Šios kolonijos susidūrė su vis sudėtingesnėmis problemomis dėl savo dydžio ir formos – visos kolonijos ląstelės turėjo būti aprūpintos viskuo, kas reikalinga gyvybei. Šios problemos buvo išspręstos dėl tam tikros kolonijų struktūros ir darbo pasidalijimo tarp jas sudarančių ląstelių. Paprastos kolonijos virto ląstelių būsenomis, tai yra daugialąsčiais organizmais. Taip pat buvo išspręstos jų, kaip sudėtingų struktūrų, savaiminio dauginimosi problemos, ir tai buvo įgyvendinta taip, kad kiekvienas organizmas galėtų vystytis iš vienos ląstelės, įdiegus sudėtingą genetinę programą, reguliuojančią ląstelių dalijimąsi ir tarpusavio sąveiką.
Tačiau ši standartinė biologinių žinių pateikimo tvarka skiriasi nuo to, kaip jos buvo gautos. Ir jie buvo gauti mokslui vystantis priešinga kryptimi – nuo organizmų iki organų, ląstelių, makromolekulių ir atomų. Naršydami į kiekvieną iš šių lygių, mokslininkai galėjo tik spėlioti, kaip veikia gilesnis lygis. Kažkada didžiausias dalykas, kurį jie galėjo padaryti, buvo atidaryti kūną, pažvelgti į organus ir atspėti, kaip jie veikia. Atidarius narvus, pirmiausia manyta, kad juose prisipildė tuštuma. Tada jie atrado protoplazmą, bet iš pradžių matė ją tik kaip klampų skystį, kuriame vis dėlto kažkokiu paslaptingu būdu buvo glūdi gyvybės esmė. Atrado ląstelės branduolį ir organelius. Jie rado dažų, kurie juos spalvina skirtingai, ir taip priartėjo prie jų cheminės sudėties. XIX amžiaus pabaigoje. atrado nukleino rūgštis ir išsiaiškino apytikslę jų cheminę sudėtį, tačiau specifinė jų struktūra ilgai liko paslaptimi, kurios sprendimas atrodė taip puikiai. Šis nardymas į biologijos gelmes, ko gero, sustojo. Atėjo duomenų kaupimosi šiame giliame molekuliniame lygmenyje laikotarpis. Buvo neįprastai daug detalių. Dabar išgyvename laikotarpį, kai šis didžiulis detalių skaičius pradeda jungtis į tam tikrą vientisą paveikslą – gyvo organizmo sandaros modelį. Be to, šis modelis yra toks sudėtingas, kad jo žmogaus protas negali iki galo suvokti, todėl ne tik jo konstrukcija, bet ir vizualinis aprašymas bei panaudojimas neįmanomas be šiuolaikinių kompiuterių. Tačiau iki XX amžiaus pabaigos. buvo atrasti visi pagrindiniai biologijos principai. Klasikinė genetika kelių talentingų mokslininkų pastangomis per pirmuosius tris XX amžiaus dešimtmečius išsivystė beveik visapusiškai kaip nuoseklus ir logiškas mokslas.
Klasikinė genetika yra tik ryškus tyrėjo judėjimo iš makrolygio į mikrolygį pavyzdys. Jis atkuria sistemos schemą pagal jos elgesį, priartėdamas prie jos kaip į juodąją dėžę. Tarsi ateiviai nežinomo prietaiso mechanizmai pateko į mokslininkų rankas be jokių jiems skirtų schemų ir nurodymų. Galima išskirti dvi pagrindines savybes. Pirma, tai yra nuostabus rekonstrukcijos gylis, kurį ji pasiekė neturėdama tiesioginės informacijos apie objekto struktūrą. Klasikinio genetinio požiūrio galia įspūdinga: susidorojus tik su matomais ženklais, buvo galima susidaryti idėją apie suprantamus genus, apie jų išsidėstymą kažkokiuose paslaptinguose linijiniuose nešikliuose, apie genų ir šių nešėjų pokyčius. Remiantis požymių paveldėjimo modeliu, jo pagalba buvo gautos idėjos apie genetinės informacijos nešėjų struktūrą, šios informacijos perdavimą palikuonims ir pavertimą gyvu kūnu. Antrasis bruožas – jau minėtas sintetinis, o ne analitinis genetinių žinių pobūdis, kurio pagrįstumas, pačiame jų gavimo procese, iš karto buvo įkūnytas kuriant kažkokį naują – organizmus su naujais bruožais. Pakanka gerai išstudijuoti kelių modelių objektų genetiką, tada apie kitus objektus galima spręsti pagal panašumą. Gerai žinomas Thomaso Morgano aforizmas „kas tiesa apie musę, tas ir apie dramblį“, žinoma, yra gana stiprus perdėjimas, ir tai pamatysime. Tačiau šis požiūris (kuris taip pat išreiškiamas vadinamajame homologinių eilučių dėsnyje) vis dar veikia.
Kryžminimas yra pagrindinis klasikinės genetikos metodas. Daugumą išvadų genetikai padarė stebėdami tėvų ir palikuonių bruožus, o tyrėjo veiksmus su kiekviena nauja karta lemia ankstesnėje kartoje gauti rezultatai. Todėl genetiniai tyrimai šiek tiek primena šachmatų žaidimą. Tokių tyrimų metu padarytos išvados buvo itin išsamios ir, kaip parodė tolimesnė mokslo raida, teisingos. Gregoras Mendelis savo eksperimentuose su žirniais XIX amžiaus pabaigoje. iš tikrųjų postulavo chromosomų egzistavimą ir apibūdino chromosomų elgesį mejozės metu, neturėdamas nė menkiausio supratimo apie chromosomas. Genų ryšys su chromosomomis buvo nustatytas tik XX amžiaus pradžioje ir beveik iki jo vidurio buvo stipriai įtariama, kad baltymai yra materialus paveldimumo nešėjas. Kitaip tariant, jei kitos biologijos šakos nebuvo labai atitrūkusios nuo aprašomojo požiūrio, tai genetika savo modeliuose gerokai pralenkė laiką, kai jos tiriamus objektus buvo galima apibūdinti kaip materialias esybes. Tragišku Rusijos mokslo istorijos laikotarpiu, kuris praėjusio amžiaus 30–50-aisiais pateko į ideologinį diktatą, tai paskatino genetiką paskelbti idealistiniu pseudomokslu ir mesti savo priešakyje buvusią šalį toli atgal. ir fiziškai sunaikinti geriausius genetikus.
Tokia klasikinės genetikos, kaip mokslo, galinčio padaryti teisingas išvadas apie tam tikrų ląstelių mikrostruktūrų elgseną, remiantis bruožų elgesiu kryžmuose, kognityvinė galia, net ir neįsivaizduojant, iš ko jos susideda, pirmiausia yra dėl to, kad kad genetika apima daug matematikos iš įvairių jos pramonės šakų. Ir ši aplinkybė egzistuoja dėl to, kad genetikos objektas yra ne tam tikra biologinė struktūra, o informacija. Informacija gali būti tiriama neatsižvelgiant į tai, kokioje materialioje terpėje ji įdiegta. Taigi programuotojas savo darbe neturi žinoti, kaip tiksliai jo programa bus įkūnyta kristalų būsenoje kompiuterio procesoriuje, nors jis žino, kad ji bus įgyvendinta būtent šiuo fiziniu pagrindu. Genetika iš esmės yra biologinė informatika. Anksčiau kompiuterių mokslas buvo vadinamas kibernetika. Ir tai buvo dar vienas „pseudomokslas“, kuris buvo persekiojamas Stalino ir Chruščiovo laikais, nepaisant visų skirtumų. (Laimei, tuo metu ji nebuvo taip išvystyta kaip matematikos šaka, kaip genetika kaip biologijos šaka, ir dėl to ši įmonė padarė mažiau žalos).
„klasikinė“ genetika(kartais vadinamas Mendelio, nors turima galvoje daug plačiau nei tai, ką atrado Mendelis, ir čia labiau tiktų liūdnai pagarsėjusi ideologinė stigma „Mendelizmas-Morganizmas“) gali būti apibrėžta kaip paveldimumo mokslas, veikiantis su abstrakčiais organizmo vystymosi valdymo sistemos elementais, yra atitraukti nuo savo materialaus nešiklio ir, tiesą sakant, jiems to nereikia. Atitinkamai, molekulinė genetika galima apibrėžti kaip mokslas apie molekulinius mechanizmus, kuriais grindžiamas paveldimumas. Tikiuosi, būtų nereikalinga raginti neteikti didelės reikšmės šiems ir panašiems formaliems apibrėžimams. Realioje mokslinėje „dviejų genetikų“ praktikoje, juo labiau, tarp jų nėra ribos, o patys aukščiau pateikti apibrėžimai tik nurodo bendrą mąstymo kryptį...
Tačiau yra žinoma, kad bet koks ko nors apibrėžimas yra netobulas, nes mūsų mąstymas nėra matematinė logika, o sąvokos - tai, kuo veikia mūsų mąstymas - nesiremia žodžiais - tai, kurių pagalba mes nustatome ir bendraujame su tam tikrais praradimais. mąstymo rezultatai. Sąvokos gali tik suprasti(su įvairaus ryškumo laipsniais), stebint jų sąveiką su anksčiau liudininkai sąvokos tekstų rinkinyje, kur sąvokos žymimos žodžiais. Apibrėžimas yra tik glausčiausias ir efektyviausias tekstas, priartinantis jus prie supratimo, tačiau visada bus situacijų, kai bet koks apibrėžimas neveikia (nepaisant to, kad sąvokos veikia). Kur įmanoma, stengiuosi pateikti apibrėžimus, kurie man atrodo sėkmingiausi, nesidomėdamas, kaip jie atitinka anksčiau pasiūlytus ar originalius, tačiau nežiūriu į juos per daug rimtai ir esu labai toli nuo minties, kad užrašydama juos iš diktantas ir įsiminimas gali padėti lengviau suprasti dalyką.
Iš pradžių genetiką sudarė vienintelio mokslininko, kurio nesuprato joks amžininkas ir kuris dėl asmeninio genialumo ir įvairiapusio išsilavinimo, pasiūlęs vaisingą metodiką, skrupulingai atlikęs ilgus ir plačius eksperimentus bei padaręs neakivaizdų. spekuliatyvi prielaida. Netrukus po genetikos iš naujo atradimo, tai yra, kai ji tapo daugelio mokslu, buvo nustatyta, kad paveldimumo veiksniai yra išsidėstę griežtai apibrėžta tvarka ir tam tikru atstumu vienas nuo kito keliose linijinėse struktūrose, skaičius, santykinis. kurių dydis ir elgsena sutapo su chromosomų skaičiumi, santykiniu dydžiu ir elgesiu mejozės metu. Chromosomų paveldimumo teorija buvo suformuluota 1900–1903 m. Amerikiečių citologas Williamas Settonas ir vokiečių embriologas Theodore'as Boveri, o toliau plėtojo žymus amerikiečių genetikas Thomas Morgan ir jo mokykla - Möller, Sturtevant, Brizhdes. (Tai buvo pirmas kartas nuo 1906 m., kai jie pradėjo atlikti Drosophila tyrimus ir iš pradžių planavo triušius, tačiau šio plano nepraleido jų universiteto finansų vadovas. Charlesas Woodworthas pirmasis išaugino Drosophila, jis taip pat pasiūlė kad jis galėtų tapti patogiu objektu tiriant paveldimumą.) Ir ši taip anksti gauta svarbi išvada apie paveldimumo faktorių radimą chromosomose buvo atmesta oficialaus SSRS mokslo nuo 1940-ųjų pabaigos iki septintojo dešimtmečio pradžios!
Palyginus spekuliacinius genetinius žemėlapius (santykinę genų vietą šiose struktūrose) ir įvairias chromosomų dalis, paaiškėjo, kad genai yra jose. Bet tai nėra taip reikalinga klasikinei genetikai – jos modeliai, patikrinti kryžminimo rezultatais, sudeda genus į savotiškas „virtualias chromosomas“. Taigi iki šios dienos daugumai objektų yra dviejų tipų chromosomų žemėlapiai: fizinės kortelės, tiksliai parodo, kur po mikroskopu matomose chromosomose arba DNR molekulėje yra genai, ir genetinė, arba rekombinacinės kortelės, rekonstruojant tarpusavio susitarimas genai iš kryžių. Genų tvarka šiuose dviejų tipų žemėlapiuose visiškai sutampa, santykiniai atstumai tarp jų yra toli gražu ne visada, ir tam yra gana išsamūs paaiškinimai, kurie bus aptarti vėliau.
Kaip informacijos ir kontrolės mokslas, klasikinė genetika netgi turi panašią struktūrą į matematiką. Jis visiškai remiasi spekuliatyvių apriorinių sąvokų sistema, su kuria koreliuoja stebimi reiškiniai (priešingai, pavyzdžiui, citologijai, kurios koncepcinis aparatas pristatomas remiantis akiai matomais empiriniais faktais). Deja, šias sąvokas atitinkančioje terminijoje (o sąvokos ir terminai nėra tas pats) genetikos egzistavimo metu susikaupė tam tikras neatitikimas, į kurį atkreipsiu ypatingą dėmesį, kad jūsų nesuklaidintų įvairios žodžių vartosenos genetinė literatūra. Žinoma, genetinės sąvokos įvedamos remiantis pastebėtais faktais. Tačiau pagrindinės iš jų pristatomos kaip spekuliacinės matematinės sąvokos. Genetikoje yra daug sąvokų ir atitinkamų terminų. Bet jie tikrai reikalingi, o įvedę praktiškai išsemia temą. Daugeliu atvejų pakanka palyginti stebimą reiškinį su tinkama koncepcija, ir viskas tampa aišku. Galbūt gera knyga galėtų pasitarnauti kaip genetikos vadovėlis. žodynas genetiniai terminai. Pedagogiškai teisingiau būtų įvesti konceptualųjį aparatą ir terminiją, kai prireikia. Tačiau nepakenks nuo pat pradžių supažindinti ir aptarti pagrindines sąvokas, o paskui pasižymėti vietas, kur jų reikia. Remsimės tuo, kad kai kurias sąvokas jau esate susipažinę bent jau iš mokyklos kurso, o kartais jas vartojate net prieš jas išsamiai aptardami.
1.2. organizmų požymių. Fenotipas ir genotipas.
Galbūt svarbiausia genetinė sąvoka ženklas. Genetika kaip mokslas prasidėjo būtent tuo metu, kai Gregoras Mendelis pradėjo analizuoti individualius bruožus, o ne visą paveldimumą. Ar galite man pasakyti, kas yra ženklas? O kiek jų gali būti? Ženklas yra viskas, kas siejama su asmeniu, jei tik yra būdas jį kažkaip užregistruoti. Ūgis, svoris, spalva, šauksmo ūgis, pusė uodegos ilgio, pridėta prie trečdalio nosies ilgio kvadratinės šaknies, plaukų skaičius barzdoje, urvelės ar skruzdėlyno forma, patinų, vejančių vieną patelę, skaičius, laikas, per kurį jūs negali kvėpuoti po vandeniu, kiek meilužių turi tiriamojo motina ar dukra. Nejuokauju – tarp vieno iš dopamino receptorių tam tikro varianto nešiotojų požymių yra didelis žymens „užaugo be tėvo“ dažnis (aišku, kad čia daugiau apie ženklą vienas iš tėvų, o ne aptariamas subjektas, kuris vis dėlto gali paveldėti polinkį ).
Pasirinkimas didžiulis, tačiau kuo sėkmingesnį, išmintingesnį ar šmaikštesnį ženklą pasirinksite, tuo daugiau informacijos sužinosite iš patirties. Akivaizdu, kad prie uodegos ilgio nereikėtų pridėti nosies ilgio kvadratinės šaknies, nes abu ilgiai turi tą patį matmenį ir dėl to gausite matematinę abrakadabra. Bet jei prie uodegos ilgio pridėsime kūno masės kubo šaknį, tai prasmingiau, nes masė priklauso nuo linijinių matmenų kubo ir, ištraukę kubo šaknį, gauname ilgį proporcingą reikšmę. uodegos, o pridėjus du minėtus dydžius, gauname tam tikrus linijinius matmenis.
Nesunku suprasti, kad ne visi ženklai iš begalinės įvairovės yra vienodai informatyvūs. Kai kurie yra vienodai informatyvūs, tačiau vienas kitam nieko neprideda. Pavyzdžiui, jei paimtume du tokius požymius: dešinės kojos ilgį ir kairės kojos ilgį, tai net intuityviai aišku, kad nors abiejų kojos ilgis gali šiek tiek skirtis, antroji mažai ką pridės prie pirmosios. Paimkite šiuos ženklus: kairės kojos ilgį ir aukštį. Ką apie juos galime pasakyti? Kuo didesnis ūgis, tuo ilgesnis kojų ilgis – tai gana akivaizdu. Kojų aukštis ir ilgis koreliuoja – ne daugiau, bet ne mažiau. Iš tiesų, jei paimsime žmonių pavyzdį, išmatuosime kojų aukštį ir ilgį bei apskaičiuosime koreliacijos koeficientą, tada jis bus gana artimas vienybei ir labai patikimas. Tačiau mes žinome, kad žmonės dažniausiai yra trumpakojai ir ilgakojai. O jei imtume ūgį ir kojų ilgio santykį su ūgiu, gautume du visiškai nepriklausomus požymius – linijinius matmenis ir ilgas kojas, kurias galima paveldėti savarankiškai.
Turime dviejų išmatuotų verčių santykį. Paprastai dirbant su daugeliu funkcijų nedelsiant reikia teisingo statistinio apdorojimo. Tokiam apdorojimui nėra labai patogu tvarkyti santykius. Tačiau yra matematinių metodų rinkinys, vadinamas daugiamatė statistika(ypač Pagrindinio komponento metodas kiekybiniams požymiams), leidžianti gauti N naujų požymių iš N bet kurių išmatuotų požymių, kurie yra tiesinės pirminių (jų sumos su skirtingais koeficientais) kombinacijos, kurios tarpusavyje nekoreliuos. Tai reiškia, kad kiekvienas iš jų turės nepriklausomą informaciją. Ir jei pažiūrėtume, kaip sudarytas N šių naujų bruožų, pamatysime, kad vienas iš jų atspindi, pavyzdžiui, linijinius matmenis (tai apims visus kūno, rankų, kojų ir kt. ilgius), kitas - storis, trečias – storio netolygumas (ryškus juosmuo, klubai, krūtinė), ketvirtas – santykiniai galvos dydžiai, penktas – tamsi oda ir tt Tokios savybės yra labiausiai informatyvios ir turi skirtingą indėlį į bendras objektų kintamumas, kurį taip pat galima įvertinti. Tačiau daugiamatės analizės metodai neišsprendžia ypatybių dubliavimo problemos, nes dubliavimas turi įtakos minėtam santykiniam indėliui į bendrą naujos savybės, kuriai jie priklauso, kintamumą. Ši problema matematinėje statistikoje iki šiol nebuvo išspręsta.
Ženklai gali būti labai skirtingi, tačiau jie skirstomi į dvi dideles klases - kokybės, arba alternatyva, ir kiekybinis, arba tęstinis. Požymis yra kokybinis tuo atveju, kai kintamumas pasireiškia kelių alternatyvių požymio variantų egzistavimu, tai yra individo priklausymu tam tikrai aiškiai klasei, o jo priskyrimas vienai iš klasių nekelia abejonių. Pavyzdžiui, galime išskirti dvi tokias žmonių klases kaip vyrai ir moterys. Moterys taip pat gali būti suskirstytos į keletą alternatyvių klasių. Tarkime, mergina yra apsirengusi kelnėmis arba jos kojos yra apsivilkusios vieną cilindrinį daiktą – suknelę ar sijoną. Gauname dvi klases. Paskutinį atvejį galima suskirstyti į dvi klases – apsirengusi suknele arba sijonu. Mes gauname tris moterų klases. Moterys tikrai gali išskirti daugybę alternatyvių drabužių klasių ir tuo pačiu nepatirs nė menkiausio sunkumo klasifikuodamos. Klasikiniai pavyzdžiai: žirnių žiedai balti arba violetiniai, vaisinės muselės akys vėl baltos arba violetinės spalvos; juokinga, bet abu organai gali būti ir rausvi, o tai dar viena kokybinės savybės būsena, atskira klasė. Tais atvejais, kai galima išskirti kokybinius (alternatyvius) požymius, o gamtoje nuolat aptinkami skirtingoms klasėms (variantams) priklausantys asmenys, įprasta kalbėti apie polimorfizmas, o šių požymių klasės (variantai) paprastai vadinamos morfai, arba formos Iš pradžių tai tas pats žodis, graikų ir lotynų kalbomis, tačiau antrojo reikšmė per daug dviprasmiška, todėl geriau jo vengti. Etimologiškai abu žodžiai žymi formą, tačiau kaip terminai vartojami bet kokiems požymiams, pavyzdžiui, susijusiems su spalva. Žemiau yra du Altajaus violetinės spalvos morfai - atitinkamai geltonomis ir violetinėmis gėlėmis, gamtoje sutinkami maždaug vienodu dažnumu.
https://pandia.ru/text/78/138/images/image002_73.jpg" width="283" height="311 src=">.jpg" width="347" height="453 src=">
Kadangi visi lankėme mokyklą, galime įtarti, kad baltoji ir violetinė rainelė yra homozigotinės kai kurių alelių atžvilgiu, o alyvinė – heterozigotinė šiems alelams. Bet mes (ypač aš) tokios informacijos dar neturime ir bet kuriuo atveju turime pradėti nuo trijų spalvų morfų nustatymo.
Paminėjome tris aiškias žirnių žiedų spalvos klases – baltą, violetinę ir rožinę. Tačiau Zolotodolinskaya gatvėje auga obelys purpuriniais žiedlapiais. Ir yra obelų su rausvais, šiek tiek rausvais ir baltais žiedlapiais. Kalbant apie kioskuose parduodamus gvazdikus, mums atrodo, kad žiedų spalva yra kokybės ženklas – yra raudonų, baltų, rožinių ir baltų žiedlapių su raudonu apvadu. O gėlių selekcininkai turbūt turi tokią gvazdikų įvairovę, kad požymis virsta kiekybine. Galite paimti spektrofotometrą, išskirti antocianino pigmentą iš standartinio žiedlapių mėginio ir išmatuoti purpurinės antocianino spalvos intensyvumą, išreiškiant ją skaičiumi. Ir tada gauname kiekybinis ženklas- tai ženklas, kurį galima išreikšti tikru skaičiumi. Vienas ir tas pats ženklas skirtingose situacijose gali veikti ir kiekybiškai, ir kokybiškai. Beveik bet kokio kokybinio požymio atveju galite rasti būdą, kaip jį išmatuoti ir laikyti kiekybiniu. Priešingai, dauguma kiekybinių charakteristikų negali būti laikomos kokybinėmis, nes išmatuoto parametro reikšmės retai sugrupuojamos į aiškiai atskiriamas klases.
Žmogaus ūgis (jei neįtrauksime akivaizdaus nykštukiškumo) yra tipiškas kiekybinis požymis. Kiek augimo galimybių yra normaliam žmogui? Teisingai, pasakyti neįmanoma – tai yra teigiamas realusis skaičius, o „parinkčių“ skaičius priklauso nuo to, kokiu tikslumu matuojame ir kokios yra šio dydžio fizinės ribos. Daugelio žmonių ūgį galima apibūdinti jo vidutine verte. Tačiau mums taip pat reikia kai kurių jo kintamumo savybių. Norėdami tai padaryti, turėsime ištirti kiekybinio požymio dažnio pasiskirstymą. Kitas vadovėlinis pavyzdys: jei paimsite daug žmonių, išmatuosite jų ūgį centimetro tikslumu ir pastatysite pagal ūgį taip, kad vienodo ūgio žmonės stovėtų vienoje stulpelyje, gautume tokį vaizdą: stulpelių ilgis sudaro natūras. varpo formos kreivė. Pakankamai išmatuojant ūgį ir žmonių skaičių, jis atkurs gerai žinomas tikimybių teorijoje - normalus arba Gauso skirstinys.
Dispersija" href="/text/category/dispersiya/" rel="bookmark">dispersija – vidutinis atskirų verčių nuokrypių nuo vidurkio kvadratas. Šios vertės kvadratinė šaknis suteikia standartinis nuokrypis, jo matmuo sutampa su išmatuoto dydžio matmeniu ir gali būti naudojamas kaip ypatybės paplitimo matas. Apie 70% visų normaliai paskirstytų objektų, nesvarbu, kiek jų matuojame, yra verčių diapazone nuo vidurkio atėmus standartinį nuokrypį iki vidurkio plius standartinis nuokrypis. Jei šis intervalas aplink vidurkį padvigubinamas, tada objektų bus apie 90%, jei tris kartus, tai apie 99%.
Centrinė matematinės statistikos ribinė teorema teigia, kad didelio skaičiaus nepriklausomų atsitiktinių dydžių sumos skirstinys artėja prie normalaus. Ir beveik bet koks kiekybinis bruožas susidaro veikiant daugybei daugiakrypčių ir skirtingų stiprumo veiksnių (tai ypač pasakytina apie kūno dydį). Štai kodėl dauguma kiekybinių charakteristikų paklūsta normaliajam pasiskirstymui.
Tačiau šis teiginys teisingas tik pirmuoju apytiksliu variantu. Kaip žinoma, norint įvertinti modelio priimtinumą, būtina atkreipti dėmesį į ribines sąlygas. Normalusis skirstinys yra simetriškas ir pateikiamas visai realiųjų skaičių rinkiniui nuo - iki +, nors tikimybių tankis gana greitai krenta tolstant nuo vidurkio. Kaip pavyzdį grįžkime prie ženklo „žmogaus ūgis“. Iš tiesų, mes neturime griežtos viršutinės žmogaus ūgio ribos ir kad ir kokį rekordininką rastume, niekada nėra garantijos, kad anksčiau ar vėliau aukštesnio subjekto nepavyks rasti. Tačiau yra net teorinė apatinė riba - juk žmogaus ūgis pagal apibrėžimą negali būti mažesnis už nulį. Tai reiškia, kad ribinės sąlygos neleidžia žmogui augti Gauso modelio. Be to, jei paimtume daug žmonių, pamatytume, kad jų ūgio pasiskirstymas yra šiek tiek asimetriškas ir pasviręs į dešinę - fizinė apatinė riba ties nuliu jaučiasi! Kokį modelį galime pasiūlyti vietoj Gauso, kaip adekvatesnį biologinių objektų kiekybinėms savybėms?
Pagalvokime apie tai. Ženklai formuojasi individualaus organizmo vystymosi eigoje, o tai iš tikrųjų yra labai sudėtinga cheminė reakcija, kuri vyksta kontroliuojant genams, kurie tam tikrais momentais suteikia tam tikras tam tikrų medžiagų koncentracijas. Šios koncentracijos veikia kaip faktoriai greičių lygtyse, kurios sudaro individualų reakcijų vystymąsi (pavyzdžiui, Michaelio lygtys), o ženklų reikšmės tiesiogiai priklauso nuo kai kurių iš šių (ar net visų) greičių. Todėl individualus atskirų genų indėlis į kiekybinį požymį dažniausiai nesumuojamas, o yra dauginamas, tai yra, kiekvienas genas požymio vertę padidina arba sumažina kelis kartus. Daugelio nepriklausomų atsitiktinių dydžių sandauga linkusi lognormalus pasiskirstymas. Dėl to realūs organizmų kiekybinių požymių pasiskirstymai yra ne normalūs, o log-normalūs. Jie tikrai labai panašūs, bet vis tiek kiek asimetriški – švelnesni į dešinę.
https://pandia.ru/text/78/138/images/image007_23.jpg" width="304" height="416 src=">
Įprasti (A ir B) ir žemaūgiai (C) žirniai
Būtent šis bruožas – santykinis tarpubamblio ilgis – čia yra alternatyvus požymis, o augalų augimas labai retai elgiasi kaip tikras alternatyvus požymis.
Yra dar viena sąlyginai išskiriama savybių klasė, kurią reikia aiškiai suvokti. Paimkime tokį ženklą kaip procesų skaičius ant elnio ragų. Smulkiausi rageliai nešakoti. Maksimaliu atveju turime 10 procesų abiejuose raguose. Mes nepatirsime jokių sunkumų priskirdami tą ar kitą ragą klasei su tam tikru procesų skaičiumi ir tuo remdamiesi galime manyti, kad tai yra kokybinė savybė. Tačiau kokybė čia koreliuoja su sveikuoju skaičiumi, o klasių skaičius, kaip ir sveikųjų skaičių serija, yra neribotas (niekas negali garantuoti, kad anksčiau ar vėliau nesusidursime su elniu su 11 ar daugiau procesų). Tokie ženklai vadinami skaičiuojamas; jie taip pat vadinami meristinė, kuris gali būti painus, nes čia reikia ne matuoti, o skaičiuoti. Tiesą sakant, čia yra paprastas modelis – kuo didesnis ragas, tuo daugiau procesų jis turi; vien tam, kad būtų pridėtas priedas, rago užuomazgas turi įgyti tam tikrą kritinę masę. Taigi suskaičiuojamas procesų skaičius yra tik rago dydžio matas. Kalbant apie laumžirgio sparno ląstelių skaičių, tai tampa dar akivaizdžiau. Tą patį matą gauname matuodami, kai sustojame ties tam tikru jo tikslumu. Įsivaizduokite, jei skaičiuotume ne elnio ragų procesus, o plaukelius ant jaunų ragų. Tiesą sakant, mes turime įvairios priemonės rago dydžio, bet kitokiu žingsniu (apvalinimu).
Jie veikia su skaičiuojamais ženklais, naudojant tuos pačius metodus kaip ir su kiekybiniais ženklais, su kai kuriomis matematinio apdorojimo ypatybėmis. Ir būtų klaida jiems taikyti tuos pačius metodus, kurie taikomi alternatyvioms savybėms. Pavyzdžiui, viena Maskvos mokslininkų grupė tyrė ląstelių skaičių tam tikrose laumžirgio sparnų srityse. Jie suskaičiavo vidutinį ląstelių skaičių, nustatė vidutinį ir standartinį nuokrypį ir, pavyzdžiui, nustatė, kad dviejuose skirtinguose vandens telkiniuose šie vidurkiai statistiškai reikšmingai skiriasi. Jie padarė išvadą, kad šių dviejų ežerų populiacijos yra genetiškai specifinės, nes alternatyvius bruožus būtinai turi nulemti paveldimi veiksniai – vienas ar keli. Bet tada jie veikė su savo ženklu kaip su kiekybiniu! Labiausiai tikėtina, kad viename iš rezervuarų laumžirgiai išsivystė mažiau palankiomis sąlygomis ir turėjo mažesnį sparnų plotą, kuriame buvo mažiau ląstelių, kurių dydis yra gana standartizuotas pagal ontogeniškumą.
Galiausiai dažnai išskiriama trečioji didelė požymių klasė – ranginiai požymiai. Kalbame apie tuos atvejus, kai objektus galime reitinguoti pagal principą „daugiau“ / „mažiau“ („geriau“ / „blogiau“), tačiau neturime tiesioginės galimybės išreikšti šios kai kurių pranašumo savybę. kiti skaičiais. Situacijos, kuriose atsiranda reitingo ženklai, yra gana įvairios. Parado aikštelėje nesunkiai galime pastatyti karius pagal ūgį, nematuodami jų ūgio, toje pačioje vietoje pagal pečių diržus nesunkiai atpažįstame karinius laipsnius, iš anksto žinodami, kokia tvarka jie yra išdėstyti vienas kito atžvilgiu. Kai kuriais atvejais esame priversti subjektyviai vertinti kai kuriuos sudėtingus integralinius parametrus, pavyzdžiui, atskirų augalų „stiprumą“, skirstydami juos į „stiprus“, „vidutinį“ ir „silpną“.
Įdomu tai, kad kai tik turime rangus, mes jau turime apytikslį skaitinį bruožo matavimą, nors ir labai apytikslį ar subjektyvų. Taigi, eilės skaičiai yra sveikieji skaičiai. Ir su jais jau galima operuoti kaip su išmatuojamais ženklais. Su visomis tokio „matavimo“ sutartimis sukurtas matematiniai metodai, leidžianti jų pagrindu padaryti labai patikimas išvadas. Be to, net ir neabejotinos kokybinės savybės gali būti gana apytiksliai traktuojamos kaip kiekybinės. Tarkime, jei turime keturias spalvų morfas, galime jas laikyti ne vienu kokybiniu požymiu, o keturiais kiekybiniais požymiais, kurių kiekvienas gali turėti dvi reikšmes - 0 (asmuo nepriklauso šiai morfijai) ir 1 ( individas priklauso tam tikram morfui). Patirtis rodo, kad tokius panašius dirbtinius „kiekybinius požymius“ galima sėkmingai apdoroti.
Kaip rodo žirnių augimo pavyzdžiai, ta pati savybė gali būti ir kiekybinė, ir kokybinė. Bet kokia mūsų išskirtinė savybė visada gali būti kažkaip išmatuota (net priklausymas vyriškai ir moteriškai lyčiai gali būti išmatuotas kaip tam tikrų hormonų santykis). Pasirinkimą, kaip veikti su ženklu – kaip skaitinio parametro reikšme arba kaip priklausymo klasei rodikliu – lemia konkrečios užduoties specifika. Bimodalinio skirstinio atveju naudinga visus individus suskirstyti į dvi klases, bent jau kaip pirmą aproksimaciją, net jei dvi skirstinio kupros susilieja ir negalime vienareikšmiškai klasifikuoti tarp jų patenkančių individų, nebent formaliai. įvedant slenkstinę vertę.
Tiek kokybiniai, tiek kiekybiniai bruožai gali būti tam tikru mastu paveldimi, todėl patenka į genetikos akiratį. Kiekybiniams ir kokybiniams požymiams analizuoti genetika naudoja skirtingus modelius. Kokybinių bruožų paveldėjimas (būtent su jais dirbo Mendelis) kombinatorikos ir tikimybių teorijos terminais aprašomas paprasčiau ir tiksliau, ir mes daugiausiai nagrinėsime jį. Kiekybinių požymių paveldėjimas aprašomas matematinės statistikos požiūriu ir daugiausia pagrįstas koreliacijų ir skaidymo į dispersinius komponentus analize. Kaip minėta pirmiau, kokybinių savybių paveldėjimas taip pat gali būti traktuojamas kaip kiekybinių požymių paveldėjimas, o tai kai kuriais atvejais yra labai vaisingas požiūris. Tikiuosi, turėsime laiko trumpai apžvelgti kiekybinių bruožų genetikos užuomazgas. Tuo tarpu šiek tiek daugiau terminų.
Dvi ne mažiau plačios sąvokos nei ženklas, be kurių negalima apsieiti - genotipas ir fenotipas. Patys šie terminai, kaip ir terminas " genas" 1909 metais pristatė danų genetikas Wilhelmas Ludwig Johansen. Fenotipas yra viskas, kas susiję su aptariamų organizmų savybėmis, genotipas yra viskas, kas susiję su jų genais. Aišku, kad ženklų gali būti be galo daug, o genų – dešimtys tūkstančių. Be to, niekas neregistruoja didžiosios daugumos požymių ir niekas nežino didžiosios daugumos genų. Tačiau fenotipas ir genotipas yra darbinės sąvokos, kurių turinį kiekvienu atveju padiktuoja genetinis eksperimentas. Genetinis eksperimentas dažniausiai susideda iš to, kad kažkas su kuo nors sukryžminta, dažnai per daugelį kartų, ir jie seka palikuonių požymius, kuriuos pagal šiuos požymius galima atrinkti, sukryžminti ir pan. pašalintas iš gamtos, registruoti jų požymius, išsiaiškinti, kuriuos variantus reprezentuoja kai kurie genai, stebėti jų dažnių dinamiką. Kiekvienu atveju mes ieškome aiškiai apibrėžtų bruožų ir genų, dažnai kelių. Ir kai mes kalbame apie fenotipą, turime omenyje būtent šių požymių reikšmes ar būsenas, o kai kalbame apie genotipą, tada šių genų rinkinį. Yra pirmojo priklausomybė nuo antrojo, bet, kaip matysime, ne pati tiesiausia. Genetika iš esmės yra šios priklausomybės išaiškinimas. Ir tik tuo atveju, jei pati DNR seka pasirodo kaip požymis, fenotipas sutampa su genotipu.
Tik neseniai tapo įmanoma atlikti aukštųjų technologijų eksperimentus, skirtus sekti visus žinomus genus tų objektų, kuriuose jie yra žinomi (pavyzdžiui, žmonių), pavyzdžiui, pagal visos pasiuntinio RNR arba visų baltymų buvimą ar nebuvimą konkrečiame objekte. audinių. Atitinkamos sritys buvo atitinkamai pavadintos „proteomika“ ir „transkriptomika“, o visų baltymų arba pasiuntinio RNR, esančių konkrečiame objekte, visuma, atitinkamai - proteomas ir transkriptas.
1.3. Sąvokos „genas“, „lokusas“, „alelis“, „ortologas“, „paralogas“, „mutacija“.
Remdamiesi mūsų išankstiniu tvirtinimu, kad genetikoje yra daug matematikos, turėtume tikėtis terminologinio griežtumo. Deja, tai taip pat yra empirinis mokslas, egzistuojantis didžiulėje ir nevienalytėje eksperimentinėje medžiagoje, kurią atliko daugybė skirtingų specializacijų (ir skirtingo išsilavinimo!) mokslininkų. Dėl to genetikoje atsirado įvairių terminologinių „tarmių“, įskaitant labai svarbūs dalykai. Pereikime prie koncepcijos, kuri gali atrodyti esminė genetikai, bet kuri iš tikrųjų pasirodė pernelyg miglota. Pasakyk man, kas yra genas? Iš tikrųjų tai labai nesėkminga sąvoka, todėl dabar ji turi keletą reikšmių. Klasikinėje genetikoje genas yra paveldimas veiksnys, turintis įtakos organizmo savybėms. Kadaise tai buvo laikoma nedaloma paveldimumo vienetu. Po DNR struktūros atradimo greitai paaiškėjo, kad daugelis klasikinių genų yra DNR dalys, koduojančios tam tikrą baltymą, pavyzdžiui, fermentą, kuris lemia paveldimą požymį. Tai buvo didžiulis mokslo laimėjimas, ir ant šios bangos iš pradžių taip atrodė visi klasikinės genetikos genai yra būtent tokie. Buvo sukurta tokia formulė: Vienas genas – viena polipeptidinė grandinė“. Originalioje formuluotėje „vienas genas – vienas fermentas“ 1941 m. (ty 12 metų prieš Watsono ir Cricko DNR struktūros dekodavimą) pasiūlė George'as Beadle'as ir Edwardas Tathamas (rasite šių ir daugelis kitų mokslininkų vadovėlyje), kurie dirbo su neurosporų pelėsių štamais, kurie skyrėsi savo gebėjimu atlikti tam tikras biochemines reakcijas ir nustatė, kad kiekvienas genas yra atsakingas už vieną specifinę biocheminę reakciją, tai yra už tam tikrą pelėsių metabolizmo etapą. Už šiuos darbus jie gavo Nobelio premiją 1948 m. Atkreipkite dėmesį, kad tuo metu genas dar buvo suprantamas gana klasikiniu būdu, tačiau buvo atliekami aktyvūs tyrimai, siekiant išsiaiškinti, ką jis fiziškai reprezentuoja. O atradus DNR struktūrą, viskas tarsi stojo į savo vietas ir genomą imta vadinti DNR segmentu, koduojančiu polipeptidinę grandinę.
Tačiau laikui bėgant buvo nustatyta, kad šalia koduojančios sekos visada yra reguliuojančios DNR sekos, kurios pačios nieko nekoduoja, bet turi įtakos šio geno transkripcijos įjungimui-išjungimui ir intensyvumui. Jūs juos gerai žinote: tai yra promotorius - RNR polimerazės nusileidimo vieta, operatoriai - reguliuojančių baltymų nusileidimo vietos, taip pat stiprintuvai- taip pat reguliuojamųjų baltymų, kurie skatina transkripciją, bet yra tam tikru, kartais reikšmingu atstumu nuo koduojančios sekos, vietas ir duslintuvai- sekos, kurios neleidžia transkripcijai ir tt Kartais jos yra šimtuose ir tūkstančiuose nukleotidų (chromosomos skalėje tai nėra tiek daug), bet vis tiek veikia kaip cis-veiksniai (t. y. netoliese), fiziškai išsidėstę netoliese dėl tam tikros DNR krūvos. Visa ši ekonomika pradėta laikyti priklausančia genui, kuris kažką koduoja. Taigi eukariotų molekulinėje genetikoje genas yra koduojanti DNR sritis kartu su gretimomis DNR sritimis, turinčiomis įtakos jo transkripcijai.
Tokiai vietai 1957 metais S. Benzeris pasiūlė patikslinantį terminą cistronas, kas taip pat nepasisekė, nes šiuo terminu pradėta žymėti tik koduojanti DNR sritis (vadinamasis atviras skaitymo rėmas), o kartais ir DNR sritis tarp promotoriaus ir terminatoriaus, iš kurios nuskaitoma viena RNR molekulė. Prisimenate, kad prokariotuose, kuriuose molekuliniai genetiniai mechanizmai buvo pradėti aiškintis anksčiau, genų operoninė organizacija yra plačiai paplitusi, kai sekos, koduojančios kelias polipeptidines grandines, turi bendrą reguliavimą ir yra skaitomos kaip vienos mRNR dalis. Tai neleidžia vartoti anksčiau pateikto termino „genas“ apibrėžimo. Kita vertus, terminas „cistronas“ čia mažai naudingas: apibrėžiamas kaip DNR sritis, iš kurios skaitoma viena RNR, ji apims regionus, koduojančius kelis skirtingus baltymus, kurie, kita vertus, kadaise buvo vadinami "policistroninis genetinės medžiagos organizavimo principas". Dėl to terminų „genas“ ir „cistronas“ vartojimas be paaiškinimo (bent jau apie kokią karalystę kalbama) šiuo metu kyla nesusipratimų.
Atkreipkite dėmesį, kad molekuline biologine prasme genas pasirodė suskirstytas į dalis – egzonus, intronus, operatorius, stiprintuvus ir galiausiai – atskirus nukleotidus. O DNR reguliavimo seka, paimta kaip tokia, prarado teisę vadintis genu, nes pati nieko nekoduoja. Tačiau dėl poveikio geno transkripcijai ši seka taip pat gali turėti įtakos tam tikram požymiui (t. y. fenotipui), kuris bus paveldimas kartu su šia seka. Ir jis gali būti atskirtas rekombinacija nuo koduojančios sekos, ypač jei tai yra nuotolinis stipriklis. Kitaip tariant, reguliavimo seka taip pat yra ypatingas paveldimas veiksnys, kuris taip pat turi savo vietą chromosomoje. Kai kurios reguliavimo sekos, pavyzdžiui, stiprintuvai, gali paveikti kelių genų transkripciją vienu metu, t.y., užimti specifinę jų vietą reguliavimo tinkle, kuris kontroliuoja organizmo vystymąsi ir funkcionavimą. Klasikinės genetikos supratimu yra visi geno požymiai.
Šis prieštaravimas tarp klasikinės ir molekulinės biologinės geno sampratos, atsiradęs tuo metu, kai atrodė, kad visi klasikiniai genai yra transkribuotos DNR dalys, koduojančios baltymą arba RNR, neįveiktas iki šiol, o tai nėra ypač aktualu. svarbu, nes žodis „genas“ ilgą laiką nebuvo vartojamas kaip griežtas terminas. Sparčiai vystantis molekulinei biologijai, laimi molekulinė biologija: genas yra transkribuota DNR dalis kartu su reguliuojančiomis DNR sekomis. Tačiau klasikinė samprata, kad genas yra paveldimas veiksnys (nesvarbu, kaip jis funkcionuotų, koks jis yra ir iš ko susideda), buvo istoriškai pirmoji, tęsėsi daugiau nei pusę amžiaus ir pasirodė esanti itin vaisinga. Turite suvokti šį prieštaravimą ir išmokti suprasti, kas sakoma iš konteksto.
Praktikoje šis prieštaravimas sprendžiamas dviem būdais: arba prieš vartojant žodį „genas“ jo reikšmė preliminariai patikslinama, arba jis nevartojamas kaip terminas. Pirmojo atvejo pavyzdys: straipsnio, skirto genų skaičiavimui genome, skiltyje „medžiagos ir metodai“ bus parašyta, kokiu kriterijumi buvo nustatytas genas – pavyzdžiui, suskaičiuotas atvirų skaitymo kadrų skaičius. Kitame straipsnyje jie parašys: išanalizavome išraišką ir parodėme, kad kai kurie galimi rastų skaitymo rėmeliai niekada nėra transkribuojami ir, matyt, yra ne genai, o pseudogenai. Antros situacijos pavyzdys: tiriamas lokusas, iš kurio susidaro keli tūkstančiai baltymų dėl to, kad yra trys alternatyvūs promotoriai, trys alternatyvūs terminatoriai ir keliolika intronų, kuriems taikomas alternatyvus splaisingas. Kur čia yra genas ir kiek genų yra šiame lokuse? Šiuo atveju žodis „genas“ bus paminėtas tik įžangoje, kaip žodžio „locus“ sinonimas. Jei frazę, kurioje yra žodis „genas“, paimsime iš populiacijos genetinio konteksto ir įterpsime į molekulinį biologinį kontekstą, prarasime prasmę.
Skirtingi to paties geno variantai bet kuria prasme žymimi terminu aleliai. Šia forma terminą pasiūlė W. Johansenas 1926 m., remdamasis terminu „alomorfinė pora“, kurį W. Batsonas įvedė 1902 m.). „Alelio“ sąvoka atsirado tada, kai nieko nebuvo žinoma apie DNR struktūrą ir ji buvo pristatyta būtent kaip alternatyvi geno versija. Ši koncepcija ypač svarbi diploidiniams organizmams, kurie iš tėvo ir motinos gauna tą patį genų rinkinį ir dėl to kiekvienas iš jų genome yra dviem kopijomis, kurios gali būti identiškos arba skirtis, bet ne tokiai. tiek, kad negalima sakyti, kad tai „tas pats genas“. Šios dvi kopijos vadinamos aleliais.
Juokinga, bet kalbant apie terminą „alelis“, tokio paprasto klausimo, kaip šio žodžio gramatinė lytis rusų kalba, vienareikšmiško sprendimo nėra. Maskva, taip pat Kijevo ir Novosibirsko mokyklos mano, kad alelis yra vyriškas, Leningradas (Sankt Peterburgas) - moteriškas. Matote, kad net dviejuose rekomenduojamuose vadovėliuose šis žodis vartojamas įvairiai.
Terminas „aleliai“ iš pradžių buvo įvestas siekiant apibūdinti geno, atsakingo už tam tikrą požymį, variantus, susijusius su tos savybės būsena. Tačiau paaiškėjo, kad vienas nuo kito nepriklausomi genai gali vienodai paveikti tą patį požymį. Dėl to kyla problemų, kaip atskirti tų pačių ar skirtingų genų alelius. Laimei, dar anksčiau paaiškėjo, kad genai išsidėstę griežtai apibrėžta seka linijinėse struktūrose – kaip paaiškėjo, chromosomose – taip, kad kiekvienas genas vienoje iš chromosomų užima griežtai apibrėžtą vietą. Todėl kiekvieną geną buvo galima identifikuoti ne tik pagal jo įtaką požymiui, bet ir pagal vietą konkrečioje chromosomoje. Pasirodo, kiekviena chromosomos vieta yra atsakinga už tam tikrą bruožą - lokusas– užima vienas iš alelių – atskiri geno variantai. Diploidiniame branduolyje yra du kiekvieno lokuso aleliai, gauti iš motinos ir tėvo, skirtingi arba identiški. Lokusą galima apibrėžti kaip padėtis chromosomoje, kurią užima tam tikras paveldimas veiksnys, a alelis- kaip tam tikro paveldimo veiksnio variantas, ir kadangi būtent lokusas suteikia tikrumo paveldimajam veiksniui, bet alelis yra paveldimo faktoriaus, esančio tam tikrame lokuse, variantas. Akivaizdu, kad šis apibrėžimas pateiktas klasikinės genetikos požiūriu. Šiuo atveju geriau sakyti „locusas chromosomoje“. o ne „chromosomos lokusas“, nes antruoju atveju gali atrodyti, kad chromosoma susideda tik iš tokių lokusų, kurie turi genetinę reikšmę. Nors genas klasikine prasme tikrai atitinka tam tikrą chromosomos DNR segmentą ir nors labai dažnai koduojantys DNR segmentai gali kažką paveikti bent netiesiogiai (pavyzdžiui, pasikartojimų bloko buvimas gali prisidėti prie sutankinimo chromatino ir taip paveikti koduojančių DNR segmentų, esančių net dideliu atstumu nuo jos, transkripcijos intensyvumą), vis dėlto tikrai egzistuoja ilgų DNR atkarpų, kurios neturi jokio genetinio turinio, tai yra nieko neveikia ir yra ne genai jokia prasme.
Tačiau terminai „lokusas“ ir „alelis“ taip pat turi juokingą plačią reikšmę. Jeigu tirtume pačią DNR seką, kuri šiuo atveju yra ir mūsų bruožas, ir genas, kadangi ji pažodžiui koduoja save, bet kurią jos dalį, kurią galima bet kokiu būdu atpažinti, galime vadinti lokusu, o jos variantą – aleliu. . Pavyzdžiui, genome yra vadinamieji „mikrosatellitai“ – labai trumpų, iš dviejų ar trijų raidžių sudarytų, tandemų (išdėsčių viena po kitos) sekos. Šių pakartojimų skaičius labai lengvai keičiasi dėl mechanizmų, susijusių su slydimu replikacijos metu arba neteisinga rekombinacija. Tiesą sakant, dėl šių mechanizmų jie „paleidžiami“ genome, o neturi savo funkcijos ir nėra genai molekuline prasme. Dėl didelio kintamumo mikropalydovai mėgsta tyrinėti evoliucinę genetiką, nes pakartojimų kopijų skaičius gali būti naudojamas su tam tikru tikrumo laipsniu vertinant ryšį. Taigi šiuo atveju taip pat įprasta kalbėti apie alelius, šiuo žodžiu žyminčius skirtingo ilgio mikrosatellitų sekas (tai yra su skirtingu pasikartojimų kopijų skaičiumi).
Paaiškėjo, kad žodžio „genas“ klasikinėje genetikoje galima visai atsisakyti. Yra lokusas – vieta chromosomoje, kurią visada užima vienas iš alelių. Ryšys tarp lokuso ir alelio yra toks pat kaip ryšys tarp kintamojo ir jo reikšmės. Be to, pagal klasikinį apibrėžimą ir lokusas yra genas (kaip bendrinė sąvoka), ir alelis yra genas (kaip individuali sąvoka). Dažnai galite išgirsti „šie genai yra nealeliniai vienas kitam“, tai yra, jie kalba apie alelinius ir nealelinius genus, tai yra apie vieno lokuso alelius ir skirtingų lokusų alelius. Genetikos praktikoje susiformavo ne itin griežta tradicija žodį „genas“ vartoti kaip žodžio „locus“ sinonimą, tokių pavyzdžių bus ir mūsų tekste.
Tačiau pasitaiko situacijų, kai žodžio „genas“ išvengti sunku. Pavyzdžiui, raudonžiedžius žirnius apdorojo cheminiu mutagenu, o žirnius gavo baltažiedžiais. Nustatyta, kad požymis „gėlės spalva“ paveldimas taip, kaip lemia vienas lokusas – tokiais atvejais įprasta kalbėti apie monogeninisženklas (nors tikslesnis būtų neegzistuojantis terminas „monolokalinis“). Tačiau baltažiedžiai žirniai jau buvo žinomi ir šį požymį lemia gerai žinomo lokuso alelis. Kyla klausimas, ar tame pačiame lokuse gavome tą patį alelį, ar skirtingą (DNR sekos lygmeniu) alelį tame pačiame lokuse, kuris vis dėlto veda prie baltų gėlių? Arba naujo, anksčiau nežinomo lokuso alelis – kuris, tarkime, gali būti visiškai kitoks pigmento sintezės etapas? Kol tai nenustatyta, tenka tingiai sakyti: „Gavome baltai žydintį geną“. Beje, aprašoma tikra situacija iš mūsų laboratorijos gyvenimo – gavome baltąjį žydėjimą lemiantį geną, kuris pasirodė esąs alelinis mažai žinomam lokusui, atsakingam už žiedo antocianininę spalvą. a, bet į mažai žinomą vietą a2 .
Terminai lokusas ir alelis taip pat gali būti taikomi genui molekuline genetine prasme – būtent tam tikrai nukleotidų sekai. Čia terminų „lokusas“ ir „genas“ reikšmė yra ta pati, o alelis reikš tam tikro geno specifinė nukleotidų seka. Tačiau molekulinės genetikos kontekste šių terminų poreikis iškyla nedažnai, nes molekulinis biologinis svarstymas paprastai nukrypsta nuo antrojo tokio geno egzistavimo diploidiniame organizme, turinčio identišką arba šiek tiek skirtingą seką. homologinė chromosoma.
Apie egzistavimą tikriausiai žinote iš molekulinės biologijos daugiageninės šeimos: kai genome yra keli genai molekuline prasme, kurie koduoja to paties tipo baltyminį produktą – pvz., tą patį fermentą. Be to, jie gali šiek tiek skirtis pirmine struktūra: tiek DNR, tiek baltymų produktu, tiek kai kuriomis fizikinėmis ir cheminėmis baltymo produkto savybėmis – molekulinės funkcijos intensyvumu, taip pat raiškos ypatybėmis – tai yra sintezės vieta, laikas ir intensyvumas. Tas pats žirnis turi septynis (molekuline prasme) histono H1 genus, kurių kiekvienas koduoja specialų molekulės variantą, vienas iš kurių yra tik aktyviai besidalijančiose ląstelėse ir išnyksta iš dalijimąsi baigusių ląstelių chromatino. Bet kuri bet kurio iš šių genų seka būtų H1 histono geno variantas. Tačiau tame pačiame genome šie septyni genai užima skirtingus lokusus, todėl tik skirtingi konkretaus lokuso variantai bus aleliai. Jūs turite būti susipažinę su koncepcija homologija- panašumas, pagrįstas bendra kilme, ir homologai- objektai, kurie turi tokį panašumą. Molekulinėje genetikoje išskiriami du genų homologijos tipai. Homologiniai, bet nealeliniai genai tame pačiame haploidiniame genome, užimantys skirtingus lokusus, vadinami paralogai(iš graikų „para“ – šalia, šalia). Vadinami atskiri to paties lokuso variantai skirtinguose individuose ortologai(iš graikų kalbos „ortho“ – tiesiogiai, priešingai; prisiminkime orto-para izomerus organinėse medžiagose). Iš esmės ortologai yra aleliai. Tačiau terminą „ortologas“ molekuliniai biologai dažniausiai vartoja tirdami skirtingų rūšių genus – tais atvejais, kai galima vienareikšmiškai nustatyti, kad jie turi tą patį lokusą, o terminas „alelis“ vartojamas tik genų variantui. tos pačios rūšies arba glaudžiai susijusių rūšių, kurios vis dėlto gali kryžmintis (pavyzdžiui, kviečiai ir jų laukiniai giminaičiai). Taigi alelis yra genetinė sąvoka; apie alelius kalbama tada, kai iš esmės jie gali dalyvauti kryžminant.
Užduokime sau klausimą – iš kur atsirado paralogai? Logiška ir teisinga manyti, kad jie atsirado dėl genų dubliavimosi – tai yra retų geno „atsidauginimo“ atvejų genome. Natūralu, kad bet koks toks įvykis, kad ir koks retas būtų, įvyksta bet kurios rūšies ribose. Dėl to turime situaciją, kai vienų tos pačios rūšies individų genome yra du lokusai, kurie savo pirmine struktūra yra identiški (laikui bėgant gali kauptis skirtumai), o kiti turi tik vieną. Tarkime, kad dvi padauginto geno kopijos yra viena šalia kitos, todėl abu nauji lokusai yra toje pačioje vietoje, kur buvo vienas senasis. Ir taip jie pradeda kaupti skirtumus. Kur ir kokie čia aleliai? Mes apsvarstėme situaciją, kai „alelio“ sąvoka nepavyksta, ir tai yra labai gerai, nes tai darydami atsekėme jos taikymo ribą.
Beje, netikėtai nebanalus klausimas – kas yra skirtingi ir identiški aleliai. Ankstyvosiose genetikos vystymosi stadijose aleliai buvo atpažįstami tik pagal fenotipą, o skirtingais aleliais buvo laikomi tik tie, kurie lemia skirtingus fenotipus. Dažniausiai buvo du aleliai - normalus ir defektinis (mutantas), todėl ankstyvosiose genetikos vystymosi stadijose buvo populiari „buvimo-nebuvimo teorija“ (tam tikros funkcijos). Tačiau vystantis genetikai tapo žinoma vis daugiau atvejų, kai tas pats požymis turi keletą paveldimų variantų, dėl kurių galiausiai atsirado garsus aforizmas Thomas Morganas: „Vienas buvimas negali atitikti kelių nebuvimų“. O kalbant apie kiekybinius požymius, nulemtus daugelio genų vienu metu, ypatingo fenotipinio vieno alelio pasireiškimo apskritai nėra. Dėl to jie apsistojo prie to, kad aleliai buvo laikomi akivaizdžiai skirtingais, jei šiame eksperimente jie nebuvo akivaizdžiai paveldėti iš to paties individo, tai yra, jie nebuvo identiški savo kilme arba toks tapatumas nebuvo nustatytas. Pavyzdžiui, gamtoje gaudome šimtą iš pažiūros identiškų individų, siekdami ištirti nedidelius tam tikro geno fenotipinio pasireiškimo niuansus, sukryžminti juos specialiomis testerių linijomis, iš jų gautą tirtą geną perkelti į identišką genų foną, išmatuoti mus dominanti savybė – ir tuo pačiu manome, kad eksperimente dalyvauja šimtas skirtingų (pagal kilmę) normalių (!) alelių (visi jie gauti iš gamtos iš gyvybingų individų).
Jūs suprantate, kad kai tapo įmanoma iššifruoti pirminę tiriamų genų struktūrą, alelių tapatumo klausimas nustojo teorinis ir buvo sumažintas iki jų pirminės struktūros (nukleotidų sekos) tapatumo. Jei yra bent vienas pakaitalas, aleliai yra skirtingi, jei ne, jie yra vienodi, nes tai yra visiškai identiškos molekulės. Atsižvelgiant į tai, kad gali kauptis nukleotidų pakaitalai, kurių daugelis neturi įtakos lokuso funkcijai, praktiškai šis metodas mažai skiriasi nuo a priori, nes visi aleliai, nepriklausomai gauti iš skirtingų asmenų, yra skirtingi. Tačiau pakeitimų dažnis labai skiriasi priklausomai nuo lokuso – pavyzdžiui, kai kuriuose lokusuose pastebėjome identišką nukleotidų seką net ir iš skirtingų žirnių porūšių (laukinių ir auginamų) gautuose aleliuose.
Palieskime tokius negriežtus, populiarius terminus kaip „laukinio tipo aleliai“, „mutantiniai aleliai“ ir „nuliniai aleliai“. Aukščiau pateikta „buvimo-nebuvimo teorija“ daugeliu atvejų yra gana tinkama. Paimkime, pavyzdžiui, tuos pačius žirnius. Žirnių žiedai turi pigmentą – antocianiną, kuris nuspalvina juos rausvai raudona (violetinė) spalva. Jei kuris nors iš baltymų, dalyvaujančių biocheminėje antocianinų sintezės grandinėje, yra sugedęs arba jo nėra, antocianinas nesusintetinamas ir žiedai lieka balti. Tarkime, kad tam tikroje chromosomoje yra lokusas, pažymėkime jį a, kuriame yra DNR seka, koduojanti vieną iš šių baltymų. Dažniausiai sakoma ne taip griežtai, bet paprasčiau – tam tikroje chromosomoje yra genas a, kuris koduoja vieną iš šių baltymų (Žirniai turi tokį geną tokiu pavadinimu ir koduoja reguliuojantį baltymą, kuris jungiasi prie DNR, o ne fermentą, dalyvaujantį antocianino sintezėje). Tegul šis genas turi du alelius, pažymėkime juos A ir a. alelis BET koduoja normalų funkcinį baltymą. alelis a nekoduoja funkcinio baltymo. Kaip tai įmanoma – pakalbėsime vėliau, mums dabar svarbu, kad šis alelis paprasčiausiai „neveikia“ – neatliktų savo molekulinės funkcijos, net jei jis mums nežinomas. Tokiais atvejais vadinamas normalus alelis laukinis tipas/ Žirnių pavyzdyje šis terminas yra dvigubai teisingas. Žirniai yra ir auginami, ir laukiniai (gamtoje ir toliau egzistuoja tos pačios rūšies atstovai). Ir visi laukiniai žirniai yra purpuriniais žiedais, o kultūriniai – ir violetiniais, ir baltais žiedais, tačiau europinės selekcijos daržovių ir grūdų veislėse vyrauja baltieji. Šis terminas taip pat dažnai vartojamas aleliui, kuris negali sudaryti funkcinio baltyminio produkto. nulinis alelis.
Pasitaiko atvejų, kai „laukinio tipo“ arba „nulinio alelio“ sąvoka netaikytina. Pavyzdžiui, dvismailėje boružėje Adalia bipunctata Yra dvi formos – raudona su juodomis dėmėmis ir juoda su raudona. (Beje, tai vienas iš klasikinių populiacijos genetikos objektų, į šį mokslą įtrauktas Timofejevas-Resovskis.) Abu yra atstovaujami europinėje Rusijos dalyje, nė vienas nėra geresnis už kitą (tačiau Novosibirske tik rastas antrasis). Nė vienas iš jų negali būti vadinamas laukiniu tipu, priešingai nei kiti. Tačiau gali būti, kad vienas iš šių alelių yra susijęs su šio lokuso baltyminio produkto molekulinės funkcijos praradimu, kuris, kaip ir kiti individualaus vystymosi genai, greičiausiai yra veiksnys, turintis įtakos kitų genų raiškai.
Tada genetikoje yra populiarus terminas - mutacija. Istoriškai šią koncepciją pristatė Hugo De Vriesas, tam tikra prasme priartėjęs prie to, kas dabar egzistuoja siaubo filmuose – staigus paveldimų polinkių pasikeitimas, lemiantis radikalų fenotipo pasikeitimą. De Vries dirbo su viena iš raktažolės rūšių ( Oenothera), kuri, kaip vėliau paaiškėjo, pasižymi itin originalia citogenetika: dėl kelių chromosomų persitvarkymų visas genomas paveldimas kaip vienas alelis. Tačiau šis žodis tapo plačiai vartojamu terminu ir ne tik Holivude. Sergejus Sergejevičius Četverikovas, vienas iš populiacijos genetikos įkūrėjų, vartojo terminą „genovariacija“, kuris yra teisingesnis, bet neįsitvirtino (nors Četverikovas buvo vienas iš naminių genetikų, padariusių didelę įtaką pasaulio genetikai, faktiškai įkūręs populiaciją genetika). Šiuo metu pagal mutacija Supratau bet kokie pirminės DNR struktūros pokyčiai- nuo vieno nukleotido pakeitimo iki didžiulių chromosomų dalių praradimo. Noriu atkreipti jūsų dėmesį į tai, kad žodis „mutacija“ reiškia patį pasikeitimo įvykį. Tačiau negriežtoje, bet atkaklioje genetinėje praktikoje tas pats žodis „mutacija“ dažnai taikomas jos rezultatui, tai yra aleliui, atsiradusiam dėl mutacijos. Jie sako: „Eksperimente dalyvauja Drosophila - mutacijos nešiotojai baltas“. Niekas neužfiksavo paties mutacijos įvykio, dėl kurio atsirado ši klasikinė mutacija – beje, ji siejama su mobilaus genetinio elemento įterpimu į fermento geną kopijavimas, kuris juda išskirtinai retai – bet visi vietoje „mutantinio alelio“ vis sako „mutacija“. Suprantama, kad kartą įvyko mutacija, kuri sugadino normalų alelį ir susidarė mutantas. Nesunku suprasti, kad „mutantinis alelis“ taip pat yra posakio „laukinio tipo alelis“ antonimas, bet platesnis nei „nulinis alelis“, nes leidžia įvairiai nukrypti nuo laukinio tipo alelio, nes tai lemia visišką alelį. molekulinės funkcijos praradimas (tas pats "keletas nebuvimų!), o ne vedantis.
Yra dar viena labai bjauri terminologinė situacija, su kuria kai kuriems iš jūsų teks susidurti žmogaus genetikos srityje. Kaip matysime vėliau, žmogaus genetika apskritai, terminologiškai, gana smarkiai nukrypo nuo bendrosios genetikos. Priežastis ta, kad, viena vertus, ši specializuota mokslo sritis priklauso ir biologijai, ir medicinai ir yra grynai instituciškai izoliuota nuo visos kitos genetikos, ir šia prasme ji verda savo sultyse. Kita vertus, dėl savo praktinės reikšmės ši sritis yra labai didelė savo apimtimi – tyrėjų ir jų studijų, žurnalų, straipsnių skaičiumi, todėl jos vidinės tradicijos yra atsparios išoriniams poveikiams, taip pat ir iš „motinos“ bendroji genetika. Šiuolaikinė žmogaus genetika pažengė taip toli, kad daugeliu atvejų įgyvendino seną genetikų svajonę, t. y. paaiškėjo, kad tam tikrus požymius (taip pat ir patologinius) pavyko susieti su tam tikrų nukleotidų buvimu konkrečiose konkrečios pozicijose. genai. Tačiau čia įvyko apgailėtinas terminų pakeitimas. Kai palyginame daugybę alelių pirminės DNR struktūros atžvilgiu, paaiškėja, kad kai kuriose padėtyse visada yra tas pats specifinis nukleotidas, o kai kuriose pozicijose galimi nukleotidų pakeitimai. (Kyla įtarimas, kad visų žmonijos žmonių genomuose galima rasti bet kurį nukleotidą bet kurioje padėtyje, todėl kyla juokingas filosofinis klausimas – kas yra žmogaus genomas). Jie buvo teisingai pavadinti. polimorfinės padėtys– ir iš tiesų, kiekviena tokia padėtis pasižymi alternatyviu kintamumu – tai yra polimorfizmu – atsižvelgiant į tai, kuris iš keturių nukleotidų gali būti užimtas. Bet čia kažkaip buvo pakeistos sąvokos. „Polimorfizmas“ pradėtas vadinti specifiniu nukleotidu tam tikroje polimorfinėje padėtyje (tai turėtų būti vadinama „morfu“). Jie pradėjo šnekėti maždaug taip: „Mes susekvenavome tokį ir tokį geną tiek daug žmonių ir radome dvylika polimorfizmų: du – tokiose ir tokiose, šešiose tokiose ir tokiose, keturiose – tokiose ir tokiose. Du polimorfizmai, esantys tokioje ir tokioje padėtyje, parodė reikšmingą ryšį su tokio ir tokio sindromu. Greičiausiai toks pakeitimas įvyko laboratorinio žargono lygiu, kuris egzistuoja bet kuriame moksliniame darbe ir susideda iš terminijos supaprastinimo, dažnai neraštingos. Studentai, kurie ateina į laboratoriją, kartais klaidingai supranta slengo terminus ir pradeda vartoti jį visiškai rimtai. Kažkuriuo momentu nutinka taip, kad ir straipsnio autorius, ir recenzentai mokslo žurnale yra pripratę prie to paties žargono, tada jis prasiskverbia į mokslinę spaudą ir, su tam tikra tikimybe, užsifiksuoja. (Paveikslas, beje, daugiau nei pažįstamas iš populiacijos genetikos ir visiškai atkartoja specifikacijos procesą – kai atsitiktiniai įvykiai įvyksta izoliuotoje populiacijoje, jie sutampa skirtingoms lytims ir fiksuojamos tinkamų seksualinių partnerių atpažinimo sistemos anomalijos, kurie tampa norma naujoje rūšyje ir veda prie jos nekryžminimo su senąja .) Be etimologinio prieštaravimo (vienas morfas vadinamas žodžiu, rodančiu, kad yra daug morfų) ir blogo skonio, toks pakeitimas taip pat turi. pasekmė, kad tyrinėtojai, vartoję šį žargoną, atėmė iš savęs terminą „polimorfizmas“ teisingą reikšmę. O kai atsiranda būtinybė išreikšti atitinkamą sąvoką (kuri niekur nedingo), vietoj vienareikšmio termino tenka griebtis daugžodžių apibūdinimų. Tarkime, situacijose, kuriose egzistuoja „subalansuoto polimorfizmo“ terminas – kai vienas iš morfų turi pranašumą tam tikromis sąlygomis, kitas – kitomis, todėl jie egzistuoja kartu ir neišstumia vienas kito – jie visada turi griebtis ilgo. aprašymai, tokie kaip aukščiau.
Kalbant apie supažindinimą su tradicine ir ne visada nuoseklia genetine terminologija, būtina paminėti gana juokingą terminą žymeklis. Šis terminas buvo įvestas lokusams, kurie mums svarbūs ne patys, o tiek, kiek jie žymi tam tikrą chromosomos sritį. Tokio termino atsiradimas buvo siejamas su ilgu laikotarpiu, kai nebuvo žinoma labai daug genetinių lokusų. Jo prireikė situacijose, kai reikėjo įkalti naujai atrastą geną arba, kad ir kaip paradoksaliai tai skambėtų, dirbti su dar neatrastais genais. Pavyzdžiui, genų, valdančių ekonomiškai vertingas kiekybines augalų ir gyvūnų savybes, prigimtis ilgą laiką buvo visiškai nežinoma, o ir dabar apie juos mažai žinoma. Tuo pačiu metu nebuvo jokių abejonių, kad šie genai egzistuoja ir yra chromosomose. Manipuliuojant žinomais lokusais – žymenimis – pavyko nustatyti chromosomų sritis, su kuriomis siejamas tam tikras poveikis kiekybinėms savybėms, ir panaudoti juos veisimo darbe. Iš pradžių tai dažniausiai buvo „matomi žymenys“ – lokusai, turintys alelius su matomu poveikiu. Tačiau ateityje šis požiūris buvo rimtai plėtojamas dėl įsitraukimo į biocheminių požymių genetinę analizę (dažniausiai taip pat funkciškai nesusijusių su ekonomiškai vertingomis savybėmis), o vėliau atsiradus galimybei dirbti su polimorfizmu. pačios chromosomos DNR. Tai paskatino „molekulinio žymeklio“ sąvokos atsiradimą. Taigi, terminas „žymeklis“ yra tik termino „lokusas“ sinonimas, tačiau pabrėžiama, kad šis lokusas mus domina ne kaip toks, o tik kaip orientyras chromosomoje. Tačiau terminas taip priprato, kad buvo pradėtas vartoti tais atvejais, kai lokusas yra tiesiogiai tiriamas objektas. Paradoksalu, bet molekulinės filogenijos tyrimuose pačios analizuojamos sekos taip pat dažnai vadinamos žymenimis. Čia būtų galima numanyti, kad jie yra tik orientyrai laike, o nukleotidų pakaitalai juose žymi evoliucinius įvykius, kurie, žinoma, neapsiriboja tik analizuojamų sekų pokyčiais.
Genai (tiksliau, lokusai) dažniausiai žymimi santrumpos, susidedančios iš lotyniškų raidžių, taip pat iš skaičių. Tačiau už šių pavadinimų slypi pilni genų pavadinimai, lotyniški arba, dažniau, angliški. Tiek pilni vardai, tiek genų santrumpos visada rašomi kursyvu. Genams su matoma išraiška tai paprastai yra žodis, apibūdinantis mutantinį fenotipą: w – baltas(baltos musės akys), y – geltona(geltonas kūnas musėje), a – antocianinas slopinimas(žirniams) op – kiaušialąstė pistiloidas(žirniams) bth – bithoraksas- nelabai geras pavadinimas Drosophila mutacijai, kai ant metatorakso (metothorakso) atsiranda antroji sparnų pora (kaip ir ant mezotorakso) - bet parašyta taip, lyg krūtinės ląstos tagma būtų padvigubėjusi. Yra net Drosophila mutacija su oficialiu pavadinimu fuši tarazu(sutrumpintas simbolis - ftz) – japonų. Linksmi amerikiečiai pavadino vieną iš genų motinos prieš dekapentaplegija, pagal analogiją su tokiomis organizacijomis kaip „motinos prieš karą Irake“ – vaisinių muselių patelėse šios mutacijos nešiotojai, palikuonys, turintys geną, neišgyvens. dekapentaplegija. Šio geno santrumpa skamba taip pat gerai: Piktas. Kartais, o ne populiariausiuose objektuose, oficialus geno pavadinimas ir jo santrumpa nesusiję vienas su kitu: mutacija, paverčianti žirnių ūselius lapais, turi pavadinimą tl(nuo be ūselių), o pavadinimas yra raktikaulis. Jei genas yra žinomas pagal jo molekulinį produktą (baltymą arba RNR), tada pats genas bus pavadintas jo produkto vardu: mtTrnK – mitochondrijų transportavimas RNR dėl lizinas, Rbcl – ribulis bifosfatas karboksilazės didelis subvienetas. Svarbu, kad kiekviena rūšis turėtų visiškai nepriklausomą oficialią genų simbolių nomenklatūrą, o tai kelia tam tikrų sunkumų šiuo metu, kai padaugėjo objektų su išvystyta privačia genetika, o objektų, kuriuose genai tiriami ne genetinių eksperimentų, bet tiesiogiai nuskaitant DNR sekas – auga kaip lavina (pvz., jau veikia projektas „10 000 stuburinių genomų“).
Genetika prasidėjo nuo atvejų, kai kiekviename lokuse buvo žinomi tik du aleliai ir juos buvo galima atskirti rašant didžiąja arba mažąja raide, kurią inicijavo Mendelis. Dominuojančiam aleliui buvo vartojama didžioji raidė (ką tai reiškia, žinote iš mokyklos laikų, dominavimo reiškinį plačiau paliesime vėliau) – dažniausiai tai yra laukinio tipo alelis; kaip dabar sakytume – alelis su normalia, nepažeista molekuline funkcija. Tuo pačiu metu lokusas buvo pažymėtas maža raide, tai yra, jo žymėjimas sutapo su recesyviniu, tai yra, mutantiniu, nefunkciniu aleliu, nes mokslininkai pirmą kartą sužinojo apie tokio alelio egzistavimą. apie lokuso egzistavimą. Retais atvejais, kai paaiškėjo, kad mutantinis alelis yra dominuojantis, tiek jis, tiek pats lokusas buvo žymimi didžiąja raide.
Kai ir labai greitai paaiškėjo, kad lokuse yra daug alelių (dabar žinome, kad jų daug), buvo įvesti alelių žymėjimai, kurie rašomi viršutiniame indekse po lokusų žymėjimo. „+“ simbolis dažnai naudojamas kaip toks laukinio tipo alelio indeksas, kartais jo nėra. Tarkime, pačiame pirmame žinomame Drosophila lokuse baltas (w) žymimas laukinio tipo alelis w+ , alelis, atsakingas už akių baltumą w, ir atsakingas už abrikosą - wa (pilnas vardas - baltasapricot).
Atkreipiu jūsų dėmesį į tai, kad tradiciniams genetiniams objektams su išvystyta privačia genetika vis dar egzistuoja skirtingos tradicijos rašant lokusų ir jų alelių pavadinimus. Iki šiol radau tris iš jų:
Lokusai su matomu pasireiškimu rašomi mažąja arba didžiąja raide, priklausomai nuo to, ar lokusas apibūdinamas recesyviniu ar dominuojančiu aleliu laukinio tipo atžvilgiu; ir didžiosiomis raidėmis, jei lokusas žinomas iš molekulinės funkcijos. Tuo pačiu metu išlaikoma tradicija recesyvinius alelius rašyti mažąja raide, o dominuojančius – didžiąja raide. Tokia yra genetinė nomenklatūra, pavyzdžiui, žirnių ir pelių. Pavyzdžiui, žirnių lokusas a, atsakinga už žiedų spalvą, turi alelių A ir a.
Kaip ir ankstesniu atveju, tačiau didžiosios ir mažosios raidės žymint lokusą ir jo alelius yra griežtai fiksuotos. Tokia sistema naudojama Drosofiloje. Čia pavadinimai w ir W priklauso visiškai skirtingiems lokusams. baltas ir Susiraukšlėjęs. Laukinio tipo alelis čia visada žymimas indeksu „+“. (Įdomu, kad Drosophila ir pelių genetikai, kurie yra pripratę prie savo tiriamųjų priimtos sistemos, paprastai net nežino, kad egzistuoja kita lokusų pavadinimo sistema.)
Visos lokusų žymėjimo raidės visada yra didžiosios. Tokia sistema dabar naudojama žmogaus genetikoje ir buvo priimta visai neseniai.
Tie patys alelių pavadinimai naudojami fenotipams, bet visada be kursyvo. Taigi, jei aprašysite eksperimento, kurio metu stebėjote tiek daug žirnių augalų su purpuriniais žiedais ir tiek daug su baltais žiedais, rezultatus ir žinote, kad baltažiedis eksperimente yra susijęs su lokusu. a, tada atsiradimo lentelėje A ir a raidėmis pažymėsite violetinius ir baltažiedžius augalus, net jei nežinote jų genotipo. Lygiai taip pat daroma ir nustačius, kad yra kokio nors izofermento elektroforezinių variantų: ten fenotipo atitikimas genotipui yra didesnis, bet net ir ne visada vienareikšmis.
1.4. Sąvokos „homozigotas“, „heterozigotas“, „hemizigotas“.
Kiekviename diploidiniame organizme kiekviena chromosoma (išskyrus lytines chromosomas) yra pavaizduota dviem egzemplioriais – homologais, gautais atitinkamai iš tėvo ir motinos. Kiekvienas iš homologų turi tą patį lokusų rinkinį ir kiekviename iš homologų kiekvienas lokusas yra užimtas tam tikru aleliu. Todėl kiekvienas diploidinis organizmas turi po du kiekvieno lokuso alelius. Registruojant jo genotipą, iš eilės rašomi dviejų mus dominančiame lokuse (lokusuose) esančių alelių pavadinimai, pavyzdžiui, jei yra ažirnių aleliai A ir a Yra trys galimi genotipai: A A, A a ir a a.
Jei abiejuose homologuose lokusas yra vaizduojamas tuo pačiu aleliu, vadinasi, individas yra homozigotinisšiame alelyje arba šiame lokuse. Be to, kai jie sako, kad jie yra homozigotiniai lokusui, akcentuojama tai, kad abiejuose homologuose nėra skirtumų, kai sakoma, kad jie yra homozigotiniai aleliui, akcentuojama, kuriam aleliui. Jei abiejuose homologuose lokusą vaizduoja skirtingi aleliai, tai individas heterozigotinisšiam lokusui. Paprastumo dėlei homozigotiniai ir heterozigotiniai asmenys vadinami atitinkamai homozigotinis ir heterozigotas. Atsižvelgiant į tai, kas buvo pasakyta aukščiau apie alelių tapatumą / skirtumus, tikrosios homozigotos gamtoje nėra labai paplitusios. Tačiau konkrečiame eksperimente niekas nesivargina ignoruoti skirtumų, kurie nėra aptikti arba negali būti aptikti šiame eksperimente, ir laikyti homozigotais asmenis, kuriuose abi lokuso kopijos turi identišką fenotipinį pasireiškimą. Atliekant tyrimus, kuriuose dalyvauja giminingi asmenys, sutinkami žinomi homozigotai – individai, kurių abu kurio nors lokuso aleliai yra identiškos kilmės. Tokiuose tyrimuose dažnai vartojama sąvoka vidutinis heterozigotiškumas yra heterozigotinių lokusų dalis tarp visų lokusų.
Pridėkime dar vieną terminą hemizigotas- tai individas, kuriame akivaizdžiai yra ne du, o tik vienas alelis. Na, pavyzdžiui, tikriausiai žinote, kad vyrai turi tik vieną lytinė X chromosoma, o antroji lytinė chromosoma, Y chromosoma, nėra jai homologiška (išskyrus mažas sritis), nes joje nėra daugumos genetinės informacijos prisotintų regionų. Todėl aleliai iš tų X chromosomos regionų, kurie nėra atstovaujami Y chromosomoje, neturi homologų branduolyje, tai yra, jie yra hemizigote. Kartais chromosoma praranda dalį savo fragmento kartu su joje esančiais genais (arba vienu genu). Šiuo atveju šių genų aleliai homologinėje chromosomoje yra ir hemizigotoje. Tačiau genetinio eksperimento metu dažnai nežinome, kas atsitiko chromosomose, o apie genus sprendžiame tik pagal fenotipą. Šiuo atveju geno nebuvimas gali nesiskirti nuo jo „suirimo“ – jo funkcijos praradimo. Ir kol nežinosime, tarkime, molekulinio fono, bet kažkokiu būdu nepadarysim išvados, kad molekulinė funkcija prarasta, kalbėsime tik apie alelį, arba „nulinį alelį“.
Diploidiniuose organizmuose gali būti svarbu atskirti homozigotą, heterozigotą ir hemizigotą, nes dozę atitinkamas alelis genome šiuo atveju skiriasi per pusę (pavyzdžiui, esant lokusui X chromosomoje, po dvi kopijas viename genome moterims ir po vieną vyrų), o tai gali būti svarbu. Molekulinė genetika paprastai nukrypsta nuo savo subjektų homozigotiškumo / heterozigotiškumo. Tačiau ši sąvoka čia dažnai naudojama. genų dozės, tai yra, alelių, turinčių nepažeistą molekulinę funkciją, skaičius genome – dažniausiai jis svyruoja nuo 0 iki 2, bet gali būti padidintas modifikuojant geną, tai yra dirbtinai įvedant papildomas kopijas į genomą.
Kalbant apie haploidinius organizmus, įprasta sakyti, kad apskritai visi visų genų aleliai yra hemizigotoje. Kokius haploidinius organizmus turime? Prokariotai, žemesni grybai ir ascomicetai, augalų gametofitai. Atkreipkime dėmesį į vieną detalę – haploidai nėra tie, kurių ląstelėje yra griežtai vienas haploidinis genomas. Daugumoje bakterijų ląstelių yra keletas nukleoidų, kurie dar nespėjo atsiskirti – tačiau jie visi yra identiški (iki de novo mutacijų). Apatiniuose grybuose hifai dažnai visai neskirstomi į atskiras ląsteles. Svarbu, kad haploidinis organizmas savo ląstelėse turėtų vieną haploidinio genomo variantą. Galiausiai, kai kurie gyvūnai, pavyzdžiui, Hymenoptera, turi haploidinį lytį – tikriausiai žinote, kad bičių tranai yra haploidiniai. Tuo pačiu metu somatinėse ląstelėse chromosomų rinkinys padvigubėja, todėl jos nenustoja būti haploidais. Mitochondrijos ir plastidės dažniau paveldimos tik iš motinos, todėl ląstelės yra hemizigotinės šių organelių genomuose esantiems genams. Tačiau daugelyje augalų plastidės kartais turi dviejų tėvų paveldėjimą, kituose taip nutinka retkarčiais, o tėvo mitochondrijos taip pat itin retai prasiskverbia pro zigotą. Tokiais atvejais palikuonis iš abiejų tėvų gauna tam tikrą skirtingą šių organelių dalį, nebūtinai lygią 1/2. Tokiais atvejais įprasta kalbėti apie heteroplazmija.
05.05.2015 13.10.2015
Šiuolaikiniame genetikos moksle plačiai vartojami terminai aleliai, lokusai, žymenys. Tuo tarpu vaiko likimas dažnai priklauso nuo tokių siaurų terminų supratimo, nes tėvystės diagnozė tiesiogiai susijusi su šiomis sąvokomis.
Žmogaus genetinė savybė
Kiekvienas žmogus turi savo unikalų genų rinkinį, kurį gauna iš savo tėvų. Dėl tėvų genų rinkinio derinio gaunamas visiškai naujas, unikalus vaiko organizmas, turintis savo genų rinkinį.
Genetikos moksle šiuolaikiniai mokslininkai nustatė tam tikras žmogaus genų sritis, kurios turi didžiausią kintamumą – lokusus (antrasis jų pavadinimas – DNR žymenys) diagnostikai.
Bet kuris iš šių lokusų turi daugybę genetinių variacijų – alelių (alelių variantų), kurių sudėtis yra visiškai unikali ir grynai individuali kiekvienam žmogui. Pavyzdžiui, plaukų spalvos lokusas turi du galimus alelius – tamsius arba šviesius. Kiekvienas žymeklis turi savo individualų alelių skaičių. Vienų žymenų yra 7-8, kituose daugiau nei 20. Visų tirtų lokusų alelių derinys vadinamas konkretaus žmogaus DNR profiliu.
Būtent šių genų sekcijų kintamumas leidžia atlikti genetinį žmonių giminystės tyrimą, nes vaikas iš savo tėvų gauna po vieną lokusą iš kiekvieno iš tėvų.
Genetinio tyrimo principas
Genetinė biologinės tėvystės nustatymo procedūra padeda nustatyti, ar tam tikro vaiko tėvu save laikantis vyras yra tikrasis tėtis, ar šis faktas atmetamas. Biologinei tėvystei tirti analizės metu lyginami lokusai tarp tėvų ir jų vaiko.
Šiuolaikiniai DNR analizės metodai gali vienu metu tirti žmogaus genomą keliuose lokusuose. Pavyzdžiui, standartizuotas genų tyrimas apėmė 16 žymenų tyrimą vienu metu. Tačiau šiandien šiuolaikinėse laboratorijose ekspertiniai tyrimai atliekami beveik 40 lokusų.
Analizės atliekamos naudojant šiuolaikinius genų analizatorius – sekvenatorius. Išvestyje tyrėjas gauna elektroforegramą, kurioje nurodomi tiriamo mėginio lokusai ir aleliai. Taigi, atlikus DNR analizę, analizuojamas tam tikrų alelių buvimas analizuojamame DNR mėginyje.
Santykių tikimybės nustatymas
Ryšio lygiui nustatyti statistiškai apdorojami konkrečiam tyrimo dalyviui gauti DNR profiliai, kurių rezultatais ekspertas padaro išvadą apie ryšio procentinę tikimybę.
Kad būtų galima apskaičiuoti ryšio lygį, tam tikra statistinė programa lygina tų pačių alelinių variantų buvimą visuose tirtuose lokusuose iš analizuotų lokusų. Skaičiavimas atliekamas tarp visų analizės dalyvių. Skaičiavimo rezultatas – jungtinio tėvystės indekso nustatymas. Antrasis rodiklis – tėvystės tikimybė. Didelė kiekvienos iš nustatytų verčių vertė yra tiriamo vyro biologinės tėvystės įrodymas. Paprastai giminystės rodikliams apskaičiuoti naudojama alelinių dažnių duomenų bazė, gauta Rusijos gyventojams.
Teigiamas 16 skirtingų, atsitiktinai parinktų DNR žymenų palyginimas leidžia, remiantis statistika, nustatyti tėvystės tikimybę. Tačiau jei 3 ir daugiau žymenų iš 16 alelių rezultatai nesutampa, biologinės tėvystės tyrimo rezultatas laikomas neigiamu.
Tyrimo rezultatų tikslumas
Genetinių tyrimų rezultatų tikslumui įtakos turi keli veiksniai:
analizuojamų genetinių lokusų skaičius;
lokuso prigimtis.
Kuo daugiau konkrečiam asmeniui būdingų lokusų genetinė analizė leidžia tiksliau nustatyti (arba, atvirkščiai, paneigti) tėvystės tikimybės laipsnį.
Taigi pasiekiamas tikimybės laipsnis vienu metu tiriant iki 40 skirtingų lokusų yra iki 99,9%, patvirtinant biologinės tėvystės tikimybę, ir iki 100%, jei gaunamas neigiamas rezultatas.
Biologinės tėvystės nustatymas su 100% tikimybės laipsniu neįmanomas dėl teorinės galimybės egzistuoti vyrą, turintį tą patį DNR žymenų rinkinį kaip ir vaiko tėvas. Tačiau esant 99,9% tikimybės lygiui, tyrimas laikomas teigiamu, o tėvystė įrodyta.
Kokie DNR šaltiniai tinka analizei?
DNR tyrimas yra labai jautri procedūra, kurios nereikia dideli kiekiai mėginys DNR ekstrakcijai. Dėl šiuolaikinių mokslo pasiekimų, genetiniai tyrimai tėvystės tikimybei nustatyti gali būti atliekami naudojant tiek biologinę medžiagą, gautą iš konkretaus asmens (tepinėlis iš burnos, plaukų, kraujo), tiek nebiologinė medžiaga, tai yra tik kontaktuojant. su žmogumi (pavyzdžiui, jo dantų šepetėliu, drabužiu, kūdikio čiulptuku, virtuvės reikmenimis). Tai įmanoma dėl to, kad visose žmogaus ląstelėse, neatsižvelgiant į jų kilmę, DNR molekulės yra visiškai vienodos, todėl galima palyginti iš paciento burnos gautus DNR mėginius su kraujo mėginiu arba iš gauto DNR mėginio. nuo dantų šepetėlio ar drabužių.
Nauji pažanga nustatant tėvystę
Naujas žodis tėvystės apibrėžime buvo mikroschemos diagnostikos plėtra. Mikroschemoje (mažoje plokštelėje) nurodant beveik visus žmogaus genus, nustatyti tėvystę nebus sunku. Ši technologija panaši į genetinį „pasą“. Paėmus iš vaisiaus kraujo ar vaisiaus vandenų mėginį, iš jo bus galima nesunkiai išskirti DNR ir atlikti hibridizaciją ant tėvų mikrogardelių. Tyrėjai planuoja naudoti šią technologiją ir aptikti paveldimas ligas.
Gene- gyvų organizmų paveldimumo struktūrinis ir funkcinis vienetas. Genas yra DNR dalis, nurodanti tam tikro polipeptido arba funkcinės RNR seką.
Peptidai- medžiagų šeima, kurios molekulės sudarytos iš dviejų ar daugiau aminorūgščių liekanų, sujungtų į grandinę peptidiniais (amidiniais) ryšiais -C (O) NH -. Paprastai tai reiškia peptidus, susidedančius iš aminorūgščių. Peptidai, kurių seka yra trumpesnė nei 10-20 aminorūgščių liekanų, taip pat gali būti vadinami oligopeptidai, su didesniu sekos ilgiu jie vadinami polipeptidai.
baltymai paprastai vadinami polipeptidais, kuriuose yra apie 50 aminorūgščių liekanų.
Genomas- paveldimos medžiagos, esančios kūno ląstelėje, visuma. Genome yra biologinė informacija, reikalinga organizmui sukurti ir palaikyti. Dauguma genomų, įskaitant žmogaus genomą ir visų kitų ląstelių gyvybės formų genomus, yra sukurti iš DNR, tačiau kai kurie virusai turi RNR genomus. Žmonių (Homo sapiens) genomą sudaro 23 poros chromosomų, esančių branduolyje, taip pat mitochondrijų DNR. Dvidešimt dviejose autosomose, dviejose lytinėse chromosomose X ir Y, ir žmogaus mitochondrijų DNR kartu yra apie 3,1 milijardo bazinių porų.
Kartu su aplinkos veiksniais genomas lemia fenotipas organizmas.
Genotipas- tam tikro organizmo genų rinkinys, apibūdinantis individą. Terminą „genotipas“ kartu su terminais „genas“ ir „fenotipas“ 1909 m. įvedė genetikas V. L. Johansenas savo darbe „Tikslios paveldimumo doktrinos elementai“. Paprastai apie genotipą kalbama konkretaus geno kontekste; poliploidiniams individams tai reiškia tam tikro geno alelių derinį. Dauguma genų atsiranda organizmo fenotipe, tačiau fenotipas ir genotipas skiriasi šiais būdais:
- Pagal informacijos šaltinį (genotipas nustatomas tiriant individo DNR, fenotipas fiksuojamas stebint organizmo išvaizdą)
- Genotipas ne visada atitinka tą patį fenotipą. Kai kurie genai fenotipe atsiranda tik tam tikromis sąlygomis. Kita vertus, kai kurie fenotipai, pavyzdžiui, gyvūnų kailio spalva, yra kelių genų sąveikos rezultatas pagal komplementarumo tipą.
aleliai- skirtingos to paties geno formos, esančios tuose pačiuose homologinių chromosomų regionuose (lokusuose) ir lemiančios alternatyvius to paties požymio išsivystymo variantus. Diploidiniame organizme gali būti du identiški to paties geno aleliai, tokiu atveju organizmas vadinamas homozigotiniu, arba du skirtingi, todėl susidaro heterozigotinis organizmas. Terminą „alelis“ pasiūlė ir W. Johansenas (1909).
Locus- genetikoje reiškia konkretaus geno vietą genetiniame arba citologiniame chromosomos žemėlapyje. DNR sekos variantas tam tikrame lokuse vadinamas aleliu. Vadinamas sutvarkytas bet kurio genomo lokusų sąrašas genetinis žemėlapis.
Genų kartografavimas yra konkretaus biologinio požymio lokuso apibrėžimas.
Chromosomos- eukariotinės ląstelės branduolyje esančios nukleoproteinų struktūros, kuriose sutelkta didžioji dalis paveldimos informacijos ir kurios yra skirtos jai saugoti, įgyvendinti ir perduoti. Šviesos mikroskopu chromosomos aiškiai matomos tik mitozinio ar mejozinio ląstelių dalijimosi laikotarpiu. Visų ląstelės chromosomų rinkinys, vadinamas kariotipu, yra rūšiai būdingas bruožas, kuriam būdingas palyginti mažas individualaus kintamumo lygis.
Iš pradžių šis terminas buvo pasiūlytas reikšti struktūras, esančias eukariotinėse ląstelėse, tačiau pastaraisiais dešimtmečiais vis dažniau kalbama apie bakterines ar virusines chromosomas. Todėl platesnis apibrėžimas yra chromosomos, kaip struktūros, kurioje yra, apibrėžimas nukleino rūgštis ir kurių funkcija yra saugoti, įgyvendinti ir perduoti paveldimą informaciją. Eukariotinės chromosomos yra DNR turinčios struktūros branduolyje, mitochondrijose ir plastiduose. Prokariotinės chromosomos yra DNR turinčios struktūros ląstelėje be branduolio.
Virusinės chromosomos yra DNR arba RNR molekulė kapsidėje.
Locus (iš lot. locus - vieta)
chromosoma Linijinė chromosomos dalis, kurią užima vienas genas. Genetiniais ir citologiniais metodais galima nustatyti geno lokalizaciją, t.y., nustatyti, kurioje konkrečioje chromosomoje šis genas yra, taip pat jo L. padėtį kitų genų L. atžvilgiu. gulintys toje pačioje chromosomoje (žr. Genetiniai chromosomų žemėlapiai). Kaip parodyta kai kuriuose mikroorganizmuose, genai, valdantys tam tikrą biocheminių reakcijų seką, yra kaimyninėse L., o L. yra ta pačia tvarka, kuria vyksta biosintezės reakcijos; aukštesniems organizmams ši taisyklė nenustatyta. Terminas "L." genetinėje literatūroje jis kartais vartojamas kaip terminų Gene ir Cistron sinonimas.
Didžioji sovietinė enciklopedija. - M.: Tarybinė enciklopedija. 1969-1978 .
Pažiūrėkite, kas yra „Locus“ kituose žodynuose:
Vieta (-os)- * lokusas (-iai) * lokusas (-ai) 1. Tam tikro geno (jo specifinių alelių) vieta chromosomoje arba genominės DNR segmente. 2. Tam tikros mutacijos ar geno vieta genetiniame žemėlapyje. Dažnai vartojamas vietoj terminų "mutacija" ...... Genetika. enciklopedinis žodynas
- (lot. lokusas) konkretaus geno vieta chromosomos genetiniame žemėlapyje ... Didysis enciklopedinis žodynas
- (iš lot. locus place), konkretaus geno (jo alelių) vieta genetinėje. arba citologija. chromosomų žemėlapis. Kartais terminas „L“. nepagrįstai vartojamas kaip termino „genas“ sinonimas. .(Šaltinis: "Biologinis enciklopedinis žodynas". Sk. ... ... Biologinis enciklopedinis žodynas
Ak, m. (… Rusų kalbos svetimžodžių žodynas
LOCUS- (iš lot. locus place), šio geno vieta chromosomoje. Ekologinis enciklopedinis žodynas. Kišiniovas: pagrindinis moldavų leidimas Sovietinė enciklopedija. I.I. Senelis. 1989... Ekologijos žodynas
Locus- tam tikro geno (jo alelių) išsidėstymas chromosomoje ... Šaltinis: METODINĖS REKOMENDACIJOS PROGNOZAVIMAS, ANKSTYNĖS IKIKLINIKINĖS DIAGNOSTIKOS NUO INULINO PRIKLAUSOMO MELLITO (N 15) PREVENCIJA (patvirtinta Komiteto pirmininko .... .. Oficiali terminija
Egzistuoja., Sinonimų skaičius: 1 vieta (170) ASIS Sinonimų žodynas. V.N. Trišinas. 2013... Sinonimų žodynas
lokusas- Alelio geno vieta chromosomoje Biotechnologijos temos EN lokusas ... Techninis vertėjo vadovas
Šis terminas turi kitas reikšmes, žr. Locus (reikšmės). Scheminis chromosomos vaizdas: (1) Chromatid, viena iš dviejų identiškų chromosomos dalių po S fazės. (2) Centromeras, vieta, kur chromatidės yra kartu... Vikipedija
- (lot. locus), konkretaus geno vieta chromosomos genetiniame žemėlapyje. * * * LOCUS LOCUS (lot. locus), tam tikro geno vieta chromosomos genetiniame žemėlapyje... enciklopedinis žodynas
Locus locus. Geno (arba jo specifinių alelių) vieta organizmo chromosomų žemėlapyje; dažnai terminas „L“. netinkamai vartojamas vietoj termino „genas“
Knygos
- Nepilnamečių nusikaltėlių kontrolės vieta, Elena Smoleva. Straipsnyje išsamiai aptariami nepilnamečių kontrolės lokuso (subjektyvios kontrolės lygio) diagnostikos ir korekcijos klausimai. Ypatingas dėmesys skiriamas empiriniams tyrimams ...