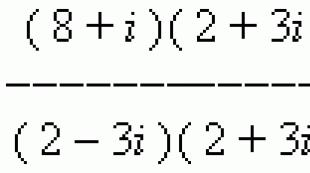សារៈសំខាន់នៃ glycolysis សម្រាប់រាងកាយ។ គ្លីកូលីស។ ខ្ញុំជាជំហានមួយ។ ការបង្កើត isomers triose phosphate
Glycolysis គឺជាដំណើរការសំខាន់នៃ catabolism នៃកាបូអ៊ីដ្រាតផ្សេងៗសម្រាប់សារពាង្គកាយមានជីវិតជាច្រើន។ វាគឺជាគាត់ដែលអនុញ្ញាតឱ្យអ្នកបង្កើតថាមពលក្នុងទម្រង់នៃម៉ូលេគុល ATP នៅក្នុងកោសិកាទាំងនោះដែលដំណើរការរស្មីសំយោគមិនកើតឡើង។ Anaerobic glycolysis កើតឡើងនៅក្នុងវត្តមានឬអវត្តមាននៃអុកស៊ីសែន។
ដំណើរការជាក់លាក់
ការសិក្សាជាច្រើនអំពីខ្លឹមសារគីមីនៃដំណើរការនេះបានបង្ហាញថា ដំណាក់កាលដំបូងនៃការ fermentation និងការដកដង្ហើមមានភាពស្រដៀងគ្នា។ អរគុណចំពោះការរកឃើញនេះ អ្នកវិទ្យាសាស្ត្របានពន្យល់ពីការរួបរួមនៅក្នុងពិភពរស់នៅ។ Anaerobic glycolysis កើតឡើងបន្ទាប់ពីវដ្តនៃអាស៊ីត tricarboxylic បានកន្លងផុតទៅការផ្ទេរអេឡិចត្រុងដោយឥតគិតថ្លៃបានបញ្ចប់។ Pyruvate ចូលទៅក្នុង mitochondria ហើយត្រូវបានកត់សុីទៅជាកាបូនឌីអុកស៊ីត។ នេះនាំឱ្យមានការបញ្ចេញថាមពលដោយឥតគិតថ្លៃពី hexose ។ អង់ស៊ីមដែលបង្កើនល្បឿននៃប្រតិកម្ម glycolytic មាននៅក្នុងកោសិកាជាច្រើនក្នុងទម្រង់រលាយក្នុង cytosol ។ ឧបករណ៍បង្កើនល្បឿននៃដំណើរការអុកស៊ីតកម្ម hexose ដែលកើតឡើងនៅក្នុងវត្តមាននៃអុកស៊ីសែនត្រូវបានប្រមូលផ្តុំនៅក្នុងភ្នាស mitochondrial ។
ដើម្បីបំបែកម៉ូលេគុលកាបូអ៊ីដ្រាតដែលរួមបញ្ចូលអាតូមកាបូនចំនួនប្រាំមួយទៅជាម៉ូលេគុលពីរ វត្តមាននៃអង់ស៊ីមសកម្មដប់គឺចាំបាច់។ អ្នកស្រាវជ្រាវបានគ្រប់គ្រងពួកវាដាច់ដោយឡែកពីគ្នាក្នុងទម្រង់ដ៏បរិសុទ្ធរបស់ពួកគេ ដើម្បីសិក្សាពីលក្ខណៈរូបវន្ត និងគីមីរបស់ពួកគេ។

ជំហានដំណើរការ
ដំណើរការនៃ glycolysis កើតឡើងនៅក្នុងកោសិកានៃសារពាង្គកាយមានជីវិត។ វាត្រូវបានអមដោយការបង្កើតអាស៊ីត pyruvic និងមានដំណាក់កាលជាច្រើន។ សម្រាប់ការវិភាគផ្លូវដង្ហើម គ្លុយកូសត្រូវតែធ្វើឱ្យសកម្ម។ ដំណើរការស្រដៀងគ្នានេះកើតឡើងនៅពេលដែលអាតូមកាបូនត្រូវបាន phosphorylated នៅពេលដែលមានអន្តរកម្មជាមួយ ATP ។
គ្លុយកូស + ATP = គ្លុយកូស -6-phosphate + ADP
ដើម្បីអនុវត្តអន្តរកម្មគីមីនេះ សារធាតុម៉ាញ៉េស្យូមស៊ីស្យូម និងហេកកូណាស (អង់ស៊ីម) ត្រូវបានប្រើ។ បន្ទាប់មកផលិតផលប្រតិកម្មត្រូវបាន isomerized ទៅ fructose-6-phosphate ។ អង់ស៊ីម phosphoglucoisomerase ត្រូវបានគេប្រើជាកាតាលីករ។
Glucolysis នៃជាតិស្ករត្រូវបានកំណត់លក្ខណៈដោយដំណាក់កាលមួយផ្សេងទៀតដែល ATP ត្រូវបានចូលរួម។ អាស៊ីតផូស្វ័រត្រូវបានភ្ជាប់ទៅនឹងអាតូមកាបូនដំបូងនៅក្នុង fructose ។ ដំណាក់កាលបន្តបន្ទាប់នៃ glycolysis ត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការ uncoupling នៃ fructose-1,6-diphosphate លទ្ធផលទៅជា triose, ការបង្កើត PHA (3-phosphoglyceraldehyde) ។
Glycolysis គឺជាដំណើរការពហុដំណាក់កាលដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការបញ្ចេញថាមពល។ នៅពេលដែលម៉ូលេគុលគ្លុយកូសមួយត្រូវបានបំបែក នោះម៉ូលេគុល PHA ពីរត្រូវបានទទួល ដូច្នេះពួកវាត្រូវបានធ្វើម្តងទៀត។
Glycolysis គឺជាដំណើរការដែលត្រូវបានសង្ខេបដោយសមីការ៖
C6H12O6 + 2ATP + 2NAD + 2Pn + 4ADP → 2PVK 2NADH + 2H+ + 4ATP + 2ADP

បទប្បញ្ញត្តិនៃ glycolysis
Glycolysis គឺជាដំណើរការដ៏សំខាន់មួយសម្រាប់សារពាង្គកាយមានជីវិត។ វាមានគោលបំណងអនុវត្តមុខងារពីរ៖
- ការបង្កើត ATP កំឡុងពេលបំបែកម៉ូលេគុល hexose;
- ការដឹកជញ្ជូនប្លុកអគារសម្រាប់ដំណើរការសំយោគជាបន្តបន្ទាប់។
បទប្បញ្ញត្តិនៃដំណើរការនេះគឺសំដៅលើការពេញចិត្តពេញលេញនៃតម្រូវការទាំងនេះនៃកោសិការស់នៅ។ ប្រតិកម្មដែលអង់ស៊ីមដើរតួជាកាតាលីករមិនអាចត្រឡប់វិញបានទេ។
សារៈសំខាន់ជាពិសេសក្នុងការអនុវត្តមុខងារបទប្បញ្ញត្តិជាកម្មសិទ្ធិរបស់ phosphofructokinase ។ អង់ស៊ីមនេះអនុវត្តមុខងារនិយតកម្ម និងកំណត់លក្ខណៈអត្រានៃដំណើរការ។

សារៈសំខាន់សរីរវិទ្យា
Glycolysis អាចត្រូវបានគេចាត់ទុកថាជាវិធីសកលនៃ catabolism ជាតិស្ករ។ វាត្រូវបានប្រើយ៉ាងសកម្មដោយសារពាង្គកាយ pro- និង eukaryotic ។ អង់ស៊ីមដែលបំប្លែង glycolysis រលាយក្នុងទឹក ហើយកកកុញនៅក្នុង cytosol ។ កោសិកា និងជាលិកាមួយចំនួនរបស់សត្វអាច catabolize hexose បានតែដោយ glycolysis ប៉ុណ្ណោះ។ ឧទាហរណ៍ កោសិកានៃប្រឡាយតំរងនោម និងណឺរ៉ូននៃខួរក្បាលមានសមត្ថភាពស្រដៀងគ្នា។
នៅក្នុងជាលិកា adipose និងថ្លើមមានភាពខុសគ្នាមួយចំនួននៅក្នុងតួនាទីសរីរវិទ្យានៃ glycolysis ។ ក្នុងអំឡុងពេលរំលាយអាហារនៅក្នុងជាលិកា adipose និងនៅក្នុងថ្លើម ដំណើរការនេះគឺជាប្រភពនៃស្រទាប់ខាងក្រោមដែលប្រើក្នុងការសំយោគខ្លាញ់។
ជាលិការុក្ខជាតិខ្លះរក្សាទុកម្សៅនៅក្នុងមើម។ រុក្ខជាតិក្នុងទឹកទទួលបានថាមពលតាមរយៈ glycolysis ។
នៅក្រោមលក្ខខណ្ឌ anaerobic, pyruvate ត្រូវបានបំលែងទៅជាអេតាណុលនិង lactate ។ ដំណើរការនេះត្រូវបានអមដោយការបញ្ចេញថាមពលយ៉ាងច្រើន។
Glycolysis មានសារៈសំខាន់ខាងសរីរវិទ្យានៅក្នុង adipocytes ។ ជាមួយនឹងជំនួយរបស់វាជំនួសឱ្យដំណើរការអុកស៊ីតកម្ម lipogenesis កើតឡើងដែលកាត់បន្ថយភាពតានតឹងអុកស៊ីតកម្ម។

សារៈសំខាន់វេជ្ជសាស្ត្រ
ជាមួយនឹងការប្រមូលផ្តុំនៃ lactate ដែលត្រូវបានបង្កើតឡើងក្រោមលក្ខខណ្ឌ anaerobic acidosis lactic មានការរីកចម្រើននៅក្នុងឈាម។ វានាំឱ្យមានការថយចុះនៃប្រតិកម្មនៃបរិយាកាសឈាមដែលត្រូវបានអមដោយការរំខានយ៉ាងខ្លាំងក្នុងការរំលាយអាហារកោសិកា។ ដំណើរការស្រដៀងគ្នានេះកើតឡើងនៅក្នុងដំណើរការ pathological ដែលទាក់ទងនឹងការផ្គត់ផ្គង់អុកស៊ីសែនខ្សោយដល់ជាលិកា។ ឧទាហរណ៍ជាមួយនឹងជំងឺ myocardial infarction, ហូរឈាម, ស្ទះសរសៃឈាមសួត។ ដំណើរការនេះគឺដោយសារតែជំងឺទឹកនោមផ្អែម ដែលដំណើរការ anaerobic កើតឡើងជំនួសឱ្យ aerobic glycolysis ។
ដោយសារអាំងស៊ុយលីនគឺជាអ្នកបង្កើនល្បឿននៃ glycolysis ក្នុងជំងឺទឹកនោមផ្អែមប្រភេទទី 1 glycolysis ត្រូវបានថយចុះ។ នោះហើយជាមូលហេតុដែលថ្នាំទាំងនោះដែលជំរុញអង់ស៊ីមដែលប្រើសម្រាប់ដំណើរការនេះអនុវត្តមុខងារនៃការព្យាបាលជំងឺ។

សេចក្តីសន្និដ្ឋាន
Glycolysis គឺជាដំណើរការដែលចាំបាច់សម្រាប់ជីវិតពេញលេញនៃសារពាង្គកាយ។ នៅក្នុងជំងឺមហារីក ការប្រើប្រាស់គ្លុយកូសកើនឡើង 10 ដង ដូច្នេះលទ្ធភាពនៃកោសិកាដុំសាច់គឺអាស្រ័យលើ glycolysis ។
បន្ទាប់ពីការសិក្សាលម្អិតអំពីលក្ខណៈពិសេសនៃលំហូរនៃដំណើរការនេះអ្នកវិទ្យាសាស្ត្របានគ្រប់គ្រងការប្រើប្រាស់ glycolysis មិនត្រឹមតែចិញ្ចឹមកោសិកាប៉ុណ្ណោះទេប៉ុន្តែថែមទាំងព្យាបាលជំងឺមួយចំនួនផងដែរ។
IN ដំណើរការ anaerobicអាស៊ីត pyruvic ត្រូវបានកាត់បន្ថយទៅជាអាស៊ីតឡាក់ទិក (lactate) ដូច្នេះនៅក្នុងមីក្រូជីវវិទ្យា glycolysis anaerobic ត្រូវបានគេហៅថា fermentation អាស៊ីតឡាក់ទិក។ lactate គឺជាមេតាប៉ូលីស ចុងបញ្ចប់បានស្លាប់ហើយបន្ទាប់មកមិនប្រែទៅជាអ្វីទាំងអស់ វិធីតែមួយគត់ដើម្បីប្រើប្រាស់ lactate គឺដើម្បីកត់សុីវាត្រឡប់ទៅ pyruvate វិញ។
កោសិការាងកាយជាច្រើនមានសមត្ថភាពអុកស៊ីតកម្ម anaerobic នៃជាតិស្ករ។ សម្រាប់ erythrocytesវាគឺជាប្រភពថាមពលតែមួយគត់។ កោសិកា សាច់ដុំគ្រោងដោយសារតែការបំបែកគ្លុយកូសដោយគ្មានអុកស៊ីហ្សែន ពួកគេអាចបំពេញការងារដ៏ខ្លាំងក្លា រហ័ស និងខ្លាំងក្លា ដូចជាការរត់ប្រណាំង ភាពតានតឹងក្នុងកីឡាកម្លាំង។ ក្រៅពីការធ្វើលំហាត់ប្រាណ ការកត់សុីដោយគ្មានអុកស៊ីហ្សែននៃជាតិគ្លុយកូសក្នុងកោសិកាត្រូវបានធ្វើឱ្យប្រសើរឡើងក្នុងអំឡុងពេលដែលមានជំងឺ hypoxia - ជាមួយនឹងប្រភេទផ្សេងៗនៃ ភាពស្លេកស្លាំង, នៅ ជំងឺឈាមរត់ជាលិកាដោយមិនគិតពីមូលហេតុ។
glycolysis
ការបំប្លែងជាតិស្ករ anaerobic កើតឡើង ស៊ីតូសូលនិងរួមបញ្ចូលទាំងដំណាក់កាលពីរនៃប្រតិកម្មអង់ស៊ីមចំនួន 11 ។
ដំណាក់កាលដំបូងនៃ glycolysis
ជំហានដំបូងនៃ glycolysis គឺ រៀបចំនៅទីនេះថាមពល ATP ត្រូវបានប្រើប្រាស់ គ្លុយកូសត្រូវបានធ្វើឱ្យសកម្ម និងការបង្កើតជាតិស្ករពីវា។ ផូស្វ័រ triose.
ប្រតិកម្មដំបូង glycolysis ត្រូវបានកាត់បន្ថយទៅនឹងការបំប្លែងគ្លុយកូសទៅជាសមាសធាតុប្រតិកម្មដោយសារតែ phosphorylation នៃអាតូមកាបូនទី 6 ដែលមិនរួមបញ្ចូលនៅក្នុងសង្វៀន។ ប្រតិកម្មនេះគឺជាការបំប្លែងជាតិគ្លុយកូសដំបូងបង្អស់ ដែលត្រូវបានជំរុញដោយ hexokinase ។
ប្រតិកម្មទីពីរចាំបាច់ដើម្បីយកអាតូមកាបូនមួយបន្ថែមទៀតចេញពីសង្វៀនសម្រាប់ phosphorylation ជាបន្តបន្ទាប់របស់វា (អង់ស៊ីម គ្លុយកូស phosphate isomerase) ជាលទ្ធផល fructose-6-phosphate ត្រូវបានបង្កើតឡើង។
ប្រតិកម្មទីបី- អង់ស៊ីម ផូស្វហ្វ័រតូគីណាសផូស្វ័រលីត fructose-6-phosphate ជាមួយនឹងការបង្កើតម៉ូលេគុល fructose-1,6-diphosphate ស្ទើរតែស៊ីមេទ្រី។ ប្រតិកម្មនេះគឺជាចំណុចកណ្តាលនៃបទប្បញ្ញត្តិនៃអត្រា glycolysis ។
IN ប្រតិកម្មទីបួន fructose 1,6-diphosphate កាត់ពាក់កណ្តាល fructose-1,6-diphosphate aldolase ជាមួយនឹងការបង្កើត phosphorylated isomers ពីរ - aldoses glyceraldehyde(GAF) និង ketoses dihydroxyacetone(DAF) ។
ប្រតិកម្មទីប្រាំដំណាក់កាលត្រៀមរៀបចំ - ការផ្លាស់ប្តូរនៃ glyceraldehyde phosphate និង dihydroxyacetone phosphate ចូលទៅក្នុងគ្នាទៅវិញទៅមកដោយមានការចូលរួម។ triose phosphate isomerase. លំនឹងនៃប្រតិកម្មត្រូវបានផ្លាស់ប្តូរនៅក្នុងការពេញចិត្តនៃ dihydroxyacetone phosphate ចំណែករបស់វាគឺ 97%, ចំណែកនៃ glyceraldehyde phosphate គឺ 3% ។ ប្រតិកម្មនេះសម្រាប់ភាពសាមញ្ញទាំងអស់របស់វាកំណត់ជោគវាសនានៃជាតិស្ករ៖
- ជាមួយនឹងកង្វះថាមពលនៅក្នុងកោសិកា និងការធ្វើឱ្យសកម្មនៃការកត់សុីគ្លុយកូស ផូស្វ័រ dihydroxyacetone ប្រែទៅជា glyceraldehyde phosphate ដែលត្រូវបានកត់សុីបន្ថែមទៀតនៅដំណាក់កាលទីពីរនៃ glycolysis ។
- ជាមួយនឹងបរិមាណគ្រប់គ្រាន់នៃ ATP ផ្ទុយទៅវិញ glyceraldehyde phosphate isomerizes ទៅជា dihydroxyacetone phosphate ហើយក្រោយមកទៀតត្រូវបានបញ្ជូនទៅការសំយោគនៃខ្លាញ់។
ដំណាក់កាលទីពីរនៃ glycolysis

ដំណាក់កាលទីពីរនៃ glycolysis គឺ ការបញ្ចេញថាមពលមាននៅក្នុង glyceraldehyde phosphate ហើយរក្សាទុកវាក្នុងទម្រង់ ATP.
ប្រតិកម្មទីប្រាំមួយ។ glycolysis (អង់ស៊ីម glyceraldehyde phosphate dehydrogenase) - ការកត់សុីនៃ glyceraldehyde phosphate និងការបន្ថែមអាស៊ីត phosphoric ទៅវានាំឱ្យមានការបង្កើតសមាសធាតុថាមពលខ្ពស់នៃអាស៊ីត 1,3-diphosphoglyceric និង NADH ។
IN ប្រតិកម្មទីប្រាំពីរ(អង់ស៊ីម phosphoglycerate kinase) ថាមពលនៃចំណង phosphoester ដែលមាននៅក្នុង 1,3-diphosphoglycerate ត្រូវបានចំណាយលើការបង្កើត ATP ។ ប្រតិកម្មបានទទួលឈ្មោះបន្ថែម - ដែលបញ្ជាក់ពីប្រភពថាមពលសម្រាប់ការទទួលបានចំណងម៉ាក្រូនៅក្នុង ATP (ពីស្រទាប់ខាងក្រោមប្រតិកម្ម) ផ្ទុយទៅនឹង phosphorylation អុកស៊ីតកម្ម (ពីជម្រាលអេឡិចត្រូគីមីនៃអ៊ីយ៉ុងអ៊ីដ្រូសែននៅលើភ្នាស mitochondrial) ។
ប្រតិកម្មទីប្រាំបី- 3-phosphoglycerate សំយោគក្នុងប្រតិកម្មមុនក្រោមឥទ្ធិពលនៃ phosphoglycerate mutase isomerizes ទៅ 2-phosphoglycerate ។
ប្រតិកម្មទីប្រាំបួន- អង់ស៊ីម អ៊ីណូឡាសផ្តាច់ម៉ូលេគុលទឹកចេញពីអាស៊ីត 2-phosphoglyceric ហើយនាំទៅរកការបង្កើតចំណង phosphoester macroergic នៅក្នុងសមាសភាពនៃ phosphoenolpyruvate ។
ប្រតិកម្មទីដប់ glycolysis គឺមួយទៀត ប្រតិកម្ម phosphorylation ស្រទាប់ខាងក្រោម- មាននៅក្នុងការផ្ទេរផូស្វាតថាមពលខ្ពស់ពី phosphoenolpyruvate ទៅ ADP ដោយ pyruvate kinase និងការបង្កើតអាស៊ីត pyruvic ។
ការពិនិត្យទូទៅ
ផ្លូវ glycolytic គឺ 10 ប្រតិកម្មជាប់គ្នាដែលនីមួយៗត្រូវបានជំរុញដោយអង់ស៊ីមផ្សេងគ្នា។
ដំណើរការនៃការ glycolysis អាចត្រូវបានបែងចែកជាពីរដំណាក់កាល។ ដំណាក់កាលទី 1 ដំណើរការជាមួយនឹងការប្រើប្រាស់ថាមពលនៃម៉ូលេគុល ATP 2 គឺការបំបែកម៉ូលេគុលគ្លុយកូសទៅជា 2 ម៉ូលេគុលនៃ glyceraldehyde-3-phosphate ។ នៅដំណាក់កាលទីពីរការកត់សុីដែលពឹងផ្អែកលើ NAD នៃ glyceraldehyde-3-phosphate កើតឡើងដែលអមដោយការសំយោគនៃ ATP ។ Glycolysis ខ្លួនវាគឺជាដំណើរការ anaerobic ទាំងស្រុង ពោលគឺវាមិនតម្រូវឱ្យមានវត្តមានអុកស៊ីសែនសម្រាប់ប្រតិកម្មកើតឡើងនោះទេ។
Glycolysis គឺជាដំណើរការមេតាបូលីសដ៏ចំណាស់បំផុតមួយដែលគេស្គាល់នៅក្នុងសារពាង្គកាយស្ទើរតែទាំងអស់។ សន្មតថា glycolysis បានបង្ហាញខ្លួនជាង 3.5 ពាន់លានឆ្នាំមុននៅក្នុង prokaryotes បឋម។
ការធ្វើមូលដ្ឋានីយកម្ម
នៅក្នុងកោសិកានៃសារពាង្គកាយ eukaryotic អង់ស៊ីមចំនួន 10 ដែលជំរុញការបំបែកជាតិស្ករទៅជា PVC ស្ថិតនៅក្នុង cytosol អង់ស៊ីមផ្សេងទៀតទាំងអស់ដែលទាក់ទងនឹងការរំលាយអាហារថាមពលគឺនៅក្នុង mitochondria និង chloroplasts ។ គ្លុយកូសចូលក្នុងកោសិកាតាមពីរវិធី៖ ស៊ីមផតដែលពឹងផ្អែកលើសូដ្យូម (ជាចម្បងសម្រាប់ enterocytes និង epithelium tubular តំរងនោម) និងសម្របសម្រួលការសាយភាយនៃជាតិស្ករដោយមានជំនួយពីប្រូតេអ៊ីនដឹកជញ្ជូន។ ការងាររបស់ប្រូតេអ៊ីនដឹកជញ្ជូនទាំងនេះត្រូវបានគ្រប់គ្រងដោយអរម៉ូន និងជាដំបូងដោយអាំងស៊ុយលីន។ ភាគច្រើន អាំងស៊ុយលីនជំរុញការដឹកជញ្ជូនគ្លុយកូសនៅក្នុងសាច់ដុំ និងជាលិកា adipose ។
លទ្ធផល
លទ្ធផលនៃ glycolysis គឺការបំប្លែងម៉ូលេគុលនៃគ្លុយកូសមួយទៅជាម៉ូលេគុលពីរនៃអាស៊ីត pyruvic (PVA) និងការបង្កើតនូវសមមូលកាត់បន្ថយពីរក្នុងទម្រង់ coenzyme NAD∙H។
សមីការពេញលេញសម្រាប់ glycolysis គឺ:
គ្លុយកូស + 2NAD + + 2ADP + 2P n \u003d 2NAD ∙ H + 2PVC + 2ATP + 2H 2 O + 2H + ។នៅក្នុងការអវត្ដមានឬកង្វះអុកស៊ីសែននៅក្នុងកោសិកា អាស៊ីត pyruvic ត្រូវបានកាត់បន្ថយទៅជាអាស៊ីតឡាក់ទិក បន្ទាប់មកសមីការទូទៅនៃ glycolysis នឹងមានដូចខាងក្រោម:
គ្លុយកូស + 2ADP + 2F n \u003d 2 lactate + 2ATP + 2H 2 O ។ដូច្នេះក្នុងអំឡុងពេលនៃការវិភាគ anaerobic នៃម៉ូលេគុលគ្លុយកូសមួយ ទិន្នផល ATP សរុបគឺម៉ូលេគុលពីរដែលទទួលបាននៅក្នុងប្រតិកម្មនៃ phosphorylation ស្រទាប់ខាងក្រោម ADP ។
នៅក្នុងសារពាង្គកាយ aerobic ផលិតផលចុងក្រោយនៃ glycolysis ឆ្លងកាត់ការផ្លាស់ប្តូរបន្ថែមទៀតនៅក្នុងវដ្តជីវគីមីដែលទាក់ទងនឹងការដកដង្ហើមកោសិកា។ ជាលទ្ធផលបន្ទាប់ពីការកត់សុីពេញលេញនៃសារធាតុរំលាយអាហារទាំងអស់នៃម៉ូលេគុលគ្លុយកូសមួយនៅដំណាក់កាលចុងក្រោយនៃការដកដង្ហើមកោសិកា - ផូស្វ័រអុកស៊ីតកម្មដែលកើតឡើងនៅលើសង្វាក់ផ្លូវដង្ហើម mitochondrial នៅក្នុងវត្តមាននៃអុកស៊ីសែន - ម៉ូលេគុល ATP បន្ថែមចំនួន 34 ឬ 36 ត្រូវបានសំយោគបន្ថែមសម្រាប់គ្លុយកូសនីមួយៗ។ ម៉ូលេគុល
ផ្លូវ
ប្រតិកម្មដំបូង glycolysis គឺ ផូស្វ័រម៉ូលេគុលគ្លុយកូសដែលកើតឡើងជាមួយនឹងការចូលរួមនៃអង់ស៊ីម hexokinase ជាលិកាជាក់លាក់ជាមួយនឹងការប្រើប្រាស់ថាមពលនៃម៉ូលេគុល 1 ATP; ទម្រង់សកម្មនៃគ្លុយកូសត្រូវបានបង្កើតឡើង - គ្លុយកូស ៦-ផូស្វាត (G-6-F):
សម្រាប់ប្រតិកម្មបន្ត វត្តមានរបស់អ៊ីយ៉ុង Mg 2+ នៅក្នុងឧបករណ៍ផ្ទុកគឺចាំបាច់ ដែលស្មុគស្មាញម៉ូលេគុល ATP ចង។ ប្រតិកម្មនេះគឺមិនអាចត្រឡប់វិញបានហើយជាលើកដំបូង ប្រតិកម្មសំខាន់នៃ glycolysis.
Phosphorylation នៃគ្លុយកូសមានគោលដៅពីរ៖ ទីមួយ ដោយសារភ្នាសប្លាស្មាដែលអាចជ្រាបចូលបានក្នុងម៉ូលេគុលគ្លុយកូសអព្យាក្រឹត មិនអនុញ្ញាតឱ្យម៉ូលេគុល G-6-P ដែលត្រូវបានចោទប្រកាន់អវិជ្ជមានឆ្លងកាត់ គ្លុយកូស phosphorylated ត្រូវបានចាក់សោនៅខាងក្នុងកោសិកា។ ទីពីរក្នុងអំឡុងពេល phosphorylation គ្លុយកូសត្រូវបានបំលែងទៅជាទម្រង់សកម្មដែលអាចចូលរួមក្នុងប្រតិកម្មជីវគីមីនិងត្រូវបានរួមបញ្ចូលនៅក្នុងវដ្តមេតាប៉ូលីស។
isoenzyme ថ្លើមរបស់ hexokinase - glucokinase - មានសារៈសំខាន់ក្នុងការគ្រប់គ្រងកម្រិតជាតិស្ករក្នុងឈាម។
នៅក្នុងប្រតិកម្មបន្ទាប់ ( 2 ) ដោយអង់ស៊ីម phosphoglucoisomerase G-6-P ត្រូវបានបំប្លែងទៅជា fructose-6-phosphate (F-6-F):

ថាមពលមិនត្រូវបានទាមទារសម្រាប់ប្រតិកម្មនេះទេ ហើយប្រតិកម្មគឺអាចបញ្ច្រាស់បានទាំងស្រុង។ នៅដំណាក់កាលនេះ fructose ក៏អាចត្រូវបានរួមបញ្ចូលនៅក្នុងដំណើរការនៃការ glycolysis ដោយ phosphorylation ។
បន្ទាប់មកប្រតិកម្មពីរកើតឡើងស្ទើរតែភ្លាមៗមួយបន្ទាប់ពីមួយផ្សេងទៀត: ផូស្វ័រដែលមិនអាចត្រឡប់វិញបាននៃ fructose-6-phosphate ( 3 ) និងការបំបែក aldol បញ្ច្រាសនៃលទ្ធផល fructose-1,6-bisphosphate (F-1,6-bF) ទៅជាពីរបី ( 4 ).
Phosphorylation នៃ F-6-F ត្រូវបានអនុវត្តដោយ phosphofructokinase ជាមួយនឹងការចំណាយថាមពលនៃម៉ូលេគុល ATP មួយផ្សេងទៀត; នេះជាលើកទីពីរ ប្រតិកម្មសំខាន់ glycolysis, បទប្បញ្ញត្តិរបស់វាកំណត់អាំងតង់ស៊ីតេនៃ glycolysis ទាំងមូល។

ការបំបែក Aldol F-1,6-bFកើតឡើងនៅក្រោមសកម្មភាពនៃ fructose-1,6-bisphosphate aldolase:

ជាលទ្ធផលនៃប្រតិកម្មទីបួន។ ផូស្វ័រ dihydroxyacetoneនិង glyceraldehyde-3-phosphateហើយទីមួយគឺស្ទើរតែភ្លាមៗនៅក្រោមសកម្មភាព phosphotriose isomeraseទៅទីពីរ 5 ) ដែលពាក់ព័ន្ធនឹងការផ្លាស់ប្តូរបន្ថែម៖

ម៉ូលេគុលនីមួយៗនៃ glyceraldehyde phosphate ត្រូវបានកត់សុីដោយ NAD+ នៅក្នុងវត្តមាននៃ glyceraldehyde phosphate dehydrogenaseពីមុន 1,3-diphosphoglycerate (6 ):

មកពី 1,3-diphosphoglycerateដោយមានចំណងម៉ាក្រូក្នុង 1 ទីតាំង អង់ស៊ីម phosphoglycerate kinase ផ្ទេរសំណល់អាស៊ីតផូស្វ័រទៅម៉ូលេគុល ADP (ប្រតិកម្ម 7 ) - ម៉ូលេគុល ATP ត្រូវបានបង្កើតឡើង៖

នេះគឺជាប្រតិកម្មដំបូងនៃ phosphorylation ស្រទាប់ខាងក្រោម។ ចាប់ពីពេលនេះតទៅ ដំណើរការនៃការបំបែកជាតិស្ករលែងមានផលចំណេញក្នុងលក្ខខណ្ឌនៃថាមពល ដោយសារតម្លៃថាមពលនៃដំណាក់កាលទី 1 ត្រូវបានផ្តល់សំណង: ម៉ូលេគុល ATP ចំនួន 2 ត្រូវបានសំយោគ (មួយសម្រាប់ 1,3-diphosphoglycerate នីមួយៗ) ជំនួសឱ្យការចំណាយពីរនៅក្នុង ប្រតិកម្ម 1 និង 3 . ដើម្បីឱ្យប្រតិកម្មនេះកើតឡើង វត្តមានរបស់ ADP នៅក្នុង cytosol ត្រូវបានទាមទារ ពោលគឺជាមួយនឹង ATP លើសនៅក្នុងកោសិកា (និងកង្វះ ADP) អត្រារបស់វាថយចុះ។ ដោយសារ ATP ដែលមិនត្រូវបានរំលាយ មិនត្រូវបានដាក់ក្នុងកោសិកា ប៉ុន្តែត្រូវបានបំផ្លាញយ៉ាងសាមញ្ញ ប្រតិកម្មនេះគឺជានិយតករដ៏សំខាន់នៃ glycolysis ។
បន្ទាប់មកតាមលំដាប់លំដោយ៖ ទម្រង់ phosphoglycerol mutase 2-phosphoglycerate (8 ):

ទម្រង់អេណូឡាស phosphoenolpyruvate (9 ):

ហើយជាចុងក្រោយ ប្រតិកម្មទីពីរនៃស្រទាប់ខាងក្រោម phosphorylation នៃ ADP កើតឡើងជាមួយនឹងការបង្កើតទម្រង់ enol នៃ pyruvate និង ATP ( 10 ):

ប្រតិកម្មកើតឡើងក្រោមសកម្មភាពរបស់ pyruvate kinase ។ នេះគឺជាប្រតិកម្មសំខាន់ចុងក្រោយនៃ glycolysis ។ Isomerization នៃទម្រង់ enol នៃ pyruvate ទៅ pyruvate កើតឡើងដោយមិនមានអង់ស៊ីម។
ចាប់តាំងពីការចាប់ផ្តើមរបស់ខ្លួន។ F-1,6-bFមានតែប្រតិកម្មបន្តជាមួយនឹងការបញ្ចេញថាមពល 7 និង 10 ដែលជាកន្លែងដែល phosphorylation ស្រទាប់ខាងក្រោមនៃ ADP កើតឡើង។
ការអភិវឌ្ឍន៍បន្ថែមទៀត
ជោគវាសនាចុងក្រោយនៃ pyruvate និង NAD∙H បង្កើតឡើងកំឡុងពេល glycolysis អាស្រ័យលើសារពាង្គកាយ និងលក្ខខណ្ឌនៅក្នុងកោសិកា ជាពិសេសវត្តមាន ឬអវត្តមាននៃអុកស៊ីសែន ឬអ្នកទទួលអេឡិចត្រុងផ្សេងទៀត។
នៅក្នុងសារពាង្គកាយ anaerobic, pyruvate និង NAD∙H ត្រូវបាន fermented បន្ថែមទៀត។ ក្នុងអំឡុងពេល fermentation អាស៊ីតឡាក់ទិក ឧទាហរណ៍នៅក្នុងបាក់តេរី pyruvate ត្រូវបានកាត់បន្ថយទៅជាអាស៊ីតឡាក់ទិកដោយសកម្មភាពនៃអង់ស៊ីម lactate dehydrogenase ។ នៅក្នុងផ្សិត ដំណើរការស្រដៀងគ្នាមួយគឺការ fermentation ជាតិអាល់កុល ដែលផលិតផលចុងក្រោយនឹងជាអេតាណុល និងកាបូនឌីអុកស៊ីត។ Butyric និង citrate fermentation ត្រូវបានគេស្គាល់ផងដែរ។
ការ fermentation Butyric៖
គ្លុយកូស → អាស៊ីត butyric + 2 CO 2 + 2 H 2 O ។
ការ fermentation ជាតិអាល់កុល:
គ្លុយកូស → 2 អេតាណុល + 2 CO 2 ។
ការ fermentation នៃក្រូចឆ្មា៖
គ្លុយកូស → អាស៊ីតនៃក្រូចឆ្មា + 2 H 2 O ។
ការ fermentation គឺចាំបាច់នៅក្នុងឧស្សាហកម្មម្ហូបអាហារ។
នៅក្នុង aerobes, pyruvate ជាធម្មតាចូលទៅក្នុងវដ្តអាស៊ីត tricarboxylic (វដ្ត Krebs) ហើយ NAD∙H ត្រូវបានកត់សុីដោយអុកស៊ីសែននៅលើសង្វាក់ផ្លូវដង្ហើមនៅក្នុង mitochondria តាមរយៈដំណើរការនៃ phosphorylation អុកស៊ីតកម្ម។
ទោះបីជាការពិតដែលថាការរំលាយអាហាររបស់មនុស្សគឺភាគច្រើនលើសលប់ aerobic អុកស៊ីតកម្ម anaerobic ត្រូវបានគេសង្កេតឃើញនៅក្នុងសាច់ដុំគ្រោងឆ្អឹងដែលធ្វើការយ៉ាងខ្លាំង។ នៅក្រោមលក្ខខណ្ឌនៃការចូលប្រើអុកស៊ីសែនមានកម្រិត pyruvate ត្រូវបានបំប្លែងទៅជាអាស៊ីតឡាក់ទិក ដូចដែលកើតឡើងក្នុងអំឡុងពេលនៃការ fermentation អាស៊ីតឡាក់ទិកនៅក្នុងអតិសុខុមប្រាណជាច្រើន៖
PVC + NAD∙H + H + → lactate + NAD + ។
ការឈឺចាប់សាច់ដុំដែលកើតឡើងមួយរយៈបន្ទាប់ពីសកម្មភាពរាងកាយខ្លាំងមិនធម្មតាត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការប្រមូលផ្តុំអាស៊ីតឡាក់ទិកនៅក្នុងពួកគេ។
ការបង្កើតអាស៊ីតឡាក់ទិកគឺជាសាខាចុងក្រោយនៃការរំលាយអាហារ ប៉ុន្តែមិនមែនជាផលិតផលចុងក្រោយនៃការរំលាយអាហារនោះទេ។ នៅក្រោមសកម្មភាពរបស់ lactate dehydrogenase អាស៊ីត lactic ត្រូវបានកត់សុីម្តងទៀត បង្កើតជា pyruvate ដែលចូលរួមក្នុងការបំប្លែងបន្ថែម។
បទប្បញ្ញត្តិនៃ glycolysis
បែងចែករវាងបទប្បញ្ញត្តិក្នុងស្រុក និងទូទៅ។
បទបញ្ជាក្នុងតំបន់ត្រូវបានអនុវត្តដោយការផ្លាស់ប្តូរសកម្មភាពនៃអង់ស៊ីមដែលស្ថិតនៅក្រោមឥទ្ធិពលនៃសារធាតុរំលាយអាហារផ្សេងៗនៅក្នុងកោសិកា។
បទប្បញ្ញត្តិនៃ glycolysis ទាំងមូលភ្លាមៗសម្រាប់សារពាង្គកាយទាំងមូលកើតឡើងក្រោមសកម្មភាពនៃអរម៉ូនដែលឥទ្ធិពលតាមរយៈម៉ូលេគុលនៃអ្នកនាំសារបន្ទាប់បន្សំផ្លាស់ប្តូរការរំលាយអាហារក្នុងកោសិកា។
អាំងស៊ុយលីនដើរតួនាទីយ៉ាងសំខាន់ក្នុងការជំរុញ glycolysis ។ Glucagon និង adrenaline គឺជាថ្នាំទប់ស្កាត់អរម៉ូនដ៏សំខាន់បំផុតនៃ glycolysis ។
អាំងស៊ុយលីនជំរុញ glycolysis តាមរយៈ៖
- ការធ្វើឱ្យសកម្មនៃប្រតិកម្ម hexokinase;
- ការរំញោចនៃ phosphofructokinase;
- ការរំញោចនៃ pyruvate kinase ។
អ័រម៉ូនផ្សេងទៀតក៏មានឥទ្ធិពលលើ glycolysis ផងដែរ។ ឧទាហរណ៍ somatotropin រារាំងអង់ស៊ីម glycolysis ហើយអរម៉ូនទីរ៉ូអ៊ីតគឺជាភ្នាក់ងាររំញោច។
Glycolysis ត្រូវបានគ្រប់គ្រងតាមជំហានសំខាន់ៗជាច្រើន។ ប្រតិកម្មដែលជំរុញដោយ hexokinase ( 1 ) ផូស្វហ្វ័រតូគីណេស ( 3 ) និង pyruvate kinase ( 10 ) ត្រូវបានកំណត់លក្ខណៈដោយការថយចុះយ៉ាងខ្លាំងនៃថាមពលដោយឥតគិតថ្លៃ និងមិនអាចត្រឡប់វិញបាន ដែលអនុញ្ញាតឱ្យពួកគេក្លាយជាចំណុចមានប្រសិទ្ធភាពនៃបទប្បញ្ញត្តិនៃ glycolysis ។
បទប្បញ្ញត្តិនៃ hexokinase
Hexokinaseរារាំងដោយផលិតផលប្រតិកម្ម - គ្លុយកូស-៦-ផូស្វាតដែលភ្ជាប់ទៅនឹងអង់ស៊ីមដោយផ្លាស់ប្តូរសកម្មភាពរបស់វា។
ដោយសារតែការពិតដែលថាភាគច្រើននៃ G-6-P នៅក្នុងកោសិកាត្រូវបានផលិតដោយការបំបែក glycogen ប្រតិកម្ម hexokinase តាមការពិតមិនចាំបាច់សម្រាប់ការកើតឡើងនៃ glycolysis ហើយ phosphorylation គ្លុយកូសនៅក្នុងបទប្បញ្ញត្តិនៃ glycolysis គឺមិនមានទេ។ សារៈសំខាន់ពិសេស។ ប្រតិកម្ម hexokinase គឺជាជំហានសំខាន់មួយក្នុងការគ្រប់គ្រងកំហាប់គ្លុយកូសក្នុងឈាម និងក្នុងកោសិកា។
ក្នុងអំឡុងពេល phosphorylation ជាតិគ្លុយកូសបាត់បង់សមត្ថភាពក្នុងការដឹកជញ្ជូនតាមរយៈភ្នាសដោយម៉ូលេគុលក្រុមហ៊ុនដឹកជញ្ជូន ដែលបង្កើតលក្ខខណ្ឌសម្រាប់ការប្រមូលផ្តុំរបស់វានៅក្នុងកោសិកា។ ការទប់ស្កាត់ hexokinase G-6-P កំណត់ការបញ្ចូលជាតិគ្លុយកូសទៅក្នុងកោសិកាការពារការប្រមូលផ្តុំលើសរបស់វា។
Glucokinase (IV isotype នៃ hexokinase) នៃថ្លើមមិនត្រូវបានរារាំងដោយគ្លុយកូស-6-ផូស្វាតទេហើយកោសិកាថ្លើមបន្តប្រមូលផ្តុំជាតិគ្លុយកូសសូម្បីតែនៅក្នុងមាតិកាខ្ពស់នៃ G-6-P ដែល glycogen ត្រូវបានសំយោគជាបន្តបន្ទាប់។ បើប្រៀបធៀបទៅនឹង isotypes ផ្សេងទៀត glucokinase មានតម្លៃខ្ពស់នៃថេរ Michaelis ពោលគឺអង់ស៊ីមដំណើរការពេញសមត្ថភាពតែក្រោមលក្ខខណ្ឌនៃកំហាប់គ្លុយកូសខ្ពស់ដែលកើតឡើងស្ទើរតែគ្រប់ពេលបន្ទាប់ពីអាហារ។
គ្លុយកូស-៦-ផូស្វាត អាចត្រូវបានបំប្លែងទៅជាគ្លុយកូសវិញដោយសកម្មភាពរបស់គ្លុយកូស-៦-ផូស្វាត។ អង់ស៊ីម glucokinase និង glucose-6-phosphatase ចូលរួមក្នុងការរក្សាកម្រិតជាតិស្ករក្នុងឈាមធម្មតា។
បទប្បញ្ញត្តិ Phosphofructokinase
អាំងតង់ស៊ីតេនៃប្រតិកម្ម phosphofructokinase មានឥទ្ធិពលសម្រេចចិត្តលើការបញ្ជូនទាំងមូលនៃ glycolysis ហើយការរំញោចនៃ phosphofructokinase ត្រូវបានចាត់ទុកថាជាជំហានសំខាន់បំផុតនៅក្នុងបទប្បញ្ញត្តិ។
Phosphofructokinase (PFK) គឺជាអង់ស៊ីម tetrameric ដែលមានជម្មើសជំនួសក្នុងស្ថានភាពអនុលោមភាពពីរ (R និង T) ដែលស្ថិតក្នុងលំនឹង ហើយឆ្លងពីមួយទៅមួយទៀត។ ATP គឺជាស្រទាប់ខាងក្រោម និងសារធាតុរារាំង allosteric នៃ PFK ។
អនុផ្នែក FFK នីមួយៗមានកន្លែងចង ATP ពីរ៖ កន្លែងដាក់ស្រទាប់ខាងក្រោម និងកន្លែងរារាំង។ បណ្តាញស្រទាប់ខាងក្រោមមានសមត្ថភាពស្មើគ្នាក្នុងការភ្ជាប់ ATP នៅក្នុងការអនុលោមតាម tetramer ណាមួយ។ ចំណែកឯទីតាំងនៃការរារាំងភ្ជាប់ ATP ទាំងស្រុងនៅពេលអង់ស៊ីមស្ថិតក្នុងស្ថានភាពអនុលោមតាម T ស្រទាប់ខាងក្រោមមួយទៀតសម្រាប់ FPA គឺ fructose 6-phosphate ដែលភ្ជាប់ទៅនឹងអង់ស៊ីមនិយមក្នុងស្ថានភាព R ។ នៅកំហាប់ខ្ពស់នៃ ATP កន្លែងរារាំងត្រូវបានកាន់កាប់ ការផ្លាស់ប្តូររវាងការអនុលោមតាមអង់ស៊ីមមិនអាចទៅរួច ហើយម៉ូលេគុលអង់ស៊ីមភាគច្រើនមានស្ថេរភាពនៅក្នុងរដ្ឋ T មិនអាចភ្ជាប់ P-6-P បានទេ។ ទោះជាយ៉ាងណាក៏ដោយការទប់ស្កាត់ ATP phosphofructokinase ត្រូវបានបង្ក្រាបដោយ AMP ដែលភ្ជាប់ទៅនឹងការអនុលោមតាម R នៃអង់ស៊ីមដូច្នេះវាធ្វើឱ្យមានស្ថេរភាពនៃស្ថានភាពនៃអង់ស៊ីមសម្រាប់ការភ្ជាប់ F-6-P ។
និយតករ allosteric ដ៏សំខាន់បំផុតនៃ glycolysis និង gluconeogenesis គឺ fructose 2,6-bisphosphateដែលមិនមែនជាតំណមធ្យមនៃវដ្តទាំងនេះ។ Fructose-2,6-bisphosphate allosterically ធ្វើឱ្យសកម្ម phosphofructokinase ។
ការសំយោគ fructose-2,6-biphosphate ត្រូវបានជំរុញដោយអង់ស៊ីមដែលមានមុខងារពិសេស - phosphofructokinase-2 / fructose-2,6-biphosphatase (FFK-2 / F-2,6-BPase) ។ នៅក្នុងទម្រង់ unphosphorylated របស់វា ប្រូតេអ៊ីនត្រូវបានគេស្គាល់ថា phosphofructokinase-2 និងមានសកម្មភាពកាតាលីករលើ fructose 6-phosphate ដែលផលិត fructose 2-6-bisphosphate ។ ជាលទ្ធផលសកម្មភាពរបស់ FFK ត្រូវបានជំរុញយ៉ាងខ្លាំងហើយសកម្មភាពរបស់ fructose-1,6-biphosphatase ត្រូវបានរារាំងយ៉ាងខ្លាំង។ នោះគឺនៅក្រោមលក្ខខណ្ឌនៃសកម្មភាព FFK-2 តុល្យភាពនៃប្រតិកម្មនេះរវាង glycolysis និង gluconeogenesis ត្រូវបានផ្លាស់ប្តូរឆ្ពោះទៅរកដំបូង - fructose-1,6-bisphosphate ត្រូវបានសំយោគ។
នៅក្នុងទម្រង់ phosphorylated អង់ស៊ីម bifunctional មិនមានសកម្មភាព kinase ទេ ផ្ទុយទៅវិញ គេហទំព័រមួយត្រូវបានធ្វើឱ្យសកម្មនៅក្នុងម៉ូលេគុលរបស់វាដែល hydrolyzes P2,6BP ទៅជា P6P និង phosphate inorganic ។ ឥទ្ធិពលមេតាប៉ូលីសនៃ phosphorylation នៃអង់ស៊ីម bifunctional គឺថាការរំញោច allosteric នៃ PFK ឈប់, allosteric inhibition នៃ F-1,6-BPase ត្រូវបានលុបចោល ហើយលំនឹងផ្លាស់ប្តូរឆ្ពោះទៅរក gluconeogenesis ។ F6F ត្រូវបានផលិតហើយបន្ទាប់មកគ្លុយកូស។
ការបំប្លែងអង់ស៊ីមទ្វេមុខងារត្រូវបានអនុវត្តដោយប្រូតេអ៊ីន kinase ដែលពឹងផ្អែកលើ cAMP (PC) ដែលតាមលំដាប់លំដោយត្រូវបានគ្រប់គ្រងដោយអរម៉ូន peptide ដែលចរាចរក្នុងឈាម។
នៅពេលដែលកំហាប់គ្លុយកូសក្នុងឈាមថយចុះ ការបង្កើតអាំងស៊ុយលីនក៏ត្រូវបានរារាំង ហើយការបញ្ចេញជាតិ glucagon ផ្ទុយទៅវិញត្រូវបានជំរុញ ហើយកំហាប់របស់វានៅក្នុងឈាមកើនឡើងយ៉ាងខ្លាំង។ Glucagon (និងអរម៉ូន contrainsular ផ្សេងទៀត) ភ្ជាប់ទៅនឹងអ្នកទទួលនៅលើភ្នាសប្លាស្មានៃកោសិកាថ្លើមដែលបណ្តាលឱ្យមានការធ្វើឱ្យសកម្មនៃភ្នាស adenylate cyclase ។ Adenylate cyclase ជំរុញការបំប្លែង ATP ទៅជា AMP ស៊ីក្លូ។ cAMP ភ្ជាប់ទៅផ្នែករងនិយតកម្មនៃប្រូតេអ៊ីន kinase ដែលបណ្តាលឱ្យមានការចេញផ្សាយ និងការធ្វើឱ្យសកម្មនៃអនុកាតាលីកររបស់វា ដែល phosphorylate អង់ស៊ីមមួយចំនួន រួមទាំង FFK-2/P-2,6-BPase ដែលមានមុខងារ។ នៅពេលដំណាលគ្នានោះ ការប្រើប្រាស់គ្លុយកូសក្នុងថ្លើមឈប់ ហើយ gluconeogenesis និង glycogenolysis ត្រូវបានធ្វើឱ្យសកម្ម ស្ដារឡើងវិញនូវ normoglycemia ។
pyruvate kinase
ជំហានបន្ទាប់ដែលបទប្បញ្ញត្តិនៃ glycolysis ត្រូវបានអនុវត្តគឺជាប្រតិកម្មចុងក្រោយ - ដំណាក់កាលនៃសកម្មភាពរបស់ pyruvate kinase ។ ចំពោះ pyruvate kinase សារធាតុ isoenzymes មួយចំនួនក៏ត្រូវបានពិពណ៌នាផងដែរ ដែលមានលក្ខណៈបទប្បញ្ញត្តិ។
ថ្លើម pyruvate kinase(L-type) ត្រូវបានគ្រប់គ្រងដោយ phosphorylation ដោយ allsteric effectors និងដោយបទប្បញ្ញត្តិនៃការបញ្ចេញហ្សែន។ អង់ស៊ីមនេះត្រូវបានរារាំងដោយ ATP និង acetyl-CoA ហើយត្រូវបានធ្វើឱ្យសកម្មដោយ fructose-1,6-bisphosphate ។ ការទប់ស្កាត់ ATP pyruvate kinase កើតឡើងស្រដៀងគ្នាទៅនឹងសកម្មភាពរបស់ ATP នៅលើ PFK ។ ការផ្សារភ្ជាប់ ATP ទៅនឹងទីតាំងនៃការរារាំងអង់ស៊ីមកាត់បន្ថយភាពស្និទ្ធស្នាលរបស់វាចំពោះ phosphoenolpyruvate ។ Hepatic pyruvate kinase ត្រូវបាន phosphorylated និងរារាំងដោយប្រូតេអ៊ីន kinase ហើយដូច្នេះក៏ស្ថិតនៅក្រោមការគ្រប់គ្រងអ័រម៉ូនផងដែរ។ លើសពីនេះទៀតសកម្មភាពនៃ pyruvate kinase ថ្លើមត្រូវបានគ្រប់គ្រងក្នុងបរិមាណផងដែរពោលគឺដោយការផ្លាស់ប្តូរកម្រិតនៃការសំយោគរបស់វា។ នេះគឺជាបទប្បញ្ញត្តិរយៈពេលវែង និងយឺត។ ការកើនឡើងនៃកាបូអ៊ីដ្រាតនៅក្នុងរបបអាហារជំរុញការបញ្ចេញមតិនៃហ្សែនដែលបានអ៊ិនកូដ pyruvate kinase ជាលទ្ធផលកម្រិតនៃអង់ស៊ីមនៅក្នុងកោសិកាកើនឡើង។
ប្រភេទ M-pyruvate kinaseត្រូវបានរកឃើញនៅក្នុងខួរក្បាល សាច់ដុំ និងជាលិកាដែលទាមទារជាតិស្ករផ្សេងទៀត មិនត្រូវបានគ្រប់គ្រងដោយប្រូតេអ៊ីន kinase ទេ។ នេះគឺជាមូលដ្ឋានគ្រឹះដែលការរំលាយអាហារនៃជាលិកាទាំងនេះត្រូវបានកំណត់ដោយតម្រូវការខាងក្នុងប៉ុណ្ណោះហើយមិនអាស្រ័យលើកម្រិតនៃជាតិស្ករក្នុងឈាមនោះទេ។
សាច់ដុំ pyruvate kinase មិនស្ថិតនៅក្រោមឥទ្ធិពលខាងក្រៅ ដូចជាការបន្ថយកម្រិតជាតិស្ករក្នុងឈាម ឬការបញ្ចេញអរម៉ូន។ លក្ខខណ្ឌក្រៅកោសិកាដែលនាំទៅរក phosphorylation និងការរារាំងនៃ isoenzyme ថ្លើមមិនផ្លាស់ប្តូរសកម្មភាពរបស់ M-type pyruvate kinase ទេ។ នោះគឺអាំងតង់ស៊ីតេនៃ glycolysis នៅក្នុងសាច់ដុំ striated ត្រូវបានកំណត់តែដោយលក្ខខណ្ឌខាងក្នុងកោសិកានិងមិនអាស្រ័យលើបទប្បញ្ញត្តិទូទៅ។
អត្ថន័យ
Glycolysis គឺជាផ្លូវ catabolic ដែលមានសារៈសំខាន់ពិសេស។ វាផ្តល់ថាមពលសម្រាប់ប្រតិកម្មកោសិកា រួមទាំងការសំយោគប្រូតេអ៊ីន។ ផលិតផលកម្រិតមធ្យមនៃ glycolysis ត្រូវបានប្រើក្នុងការសំយោគខ្លាញ់។ Pyruvate ក៏អាចត្រូវបានប្រើដើម្បីសំយោគ alanine, aspartate និងសមាសធាតុផ្សេងទៀត។ សូមអរគុណដល់ glycolysis ការអនុវត្ត mitochondrial និងអុកស៊ីសែនមិនកំណត់ថាមពលសាច់ដុំកំឡុងពេលផ្ទុកខ្លាំងរយៈពេលខ្លីនោះទេ។
សូមមើលផងដែរ
តំណភ្ជាប់
- glycolysis (ភាសាអង់គ្លេស)
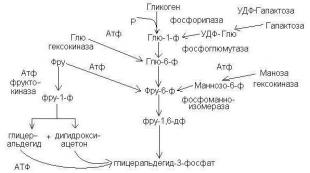
Anaerobic glycolysis គឺជាដំណើរការអង់ស៊ីមស្មុគ្រស្មាញនៃការបំប្លែងជាតិគ្លុយកូសជាបន្តបន្ទាប់ដែលកើតឡើងនៅក្នុងជាលិកាមនុស្ស និងសត្វដោយមិនប្រើប្រាស់អុកស៊ីសែន (រូបភាព 28)។
ការបំប្លែងអាស៊ីត pyruvic ទៅជាអាស៊ីតឡាក់ទិកត្រូវបានជំរុញដោយ lactate dehydrogenase៖
លទ្ធផលសរុបនៃ glycolysis ត្រូវបានបង្ហាញដោយសមីការដូចខាងក្រោមៈ C 6 H 12 O 6 + 2H 3 RO 4 + 2ADP \u003d 2C 3 H 6 O 3 + 2ATP + 2H 2 O
ដូច្នេះទិន្នផលសុទ្ធនៃ ATP ក្នុងអំឡុងពេល glycolysis anaerobic គឺ 2 mol នៃ ATP ក្នុង 1 mol នៃជាតិស្ករ។ វាគឺជាអរគុណចំពោះ glycolysis anaerobic ដែលសារពាង្គកាយមនុស្សនិងសត្វអាចអនុវត្តមុខងារសរីរវិទ្យាមួយចំនួនក្នុងរយៈពេលជាក់លាក់មួយក្រោមលក្ខខណ្ឌនៃកង្វះអុកស៊ីសែន។
ដំណើរការនេះនៅក្នុងបាក់តេរីត្រូវបានគេហៅថា fermentation អាស៊ីតឡាក់ទិក៖ វាបញ្ជាក់ពីការរៀបចំផលិតផលទឹកដោះគោដែលមានជាតិ fermented ។ Anaerobic glycolysis កើតឡើងនៅក្នុង cytosol នៃកោសិកាដែលមានអង់ស៊ីមទាំងអស់ដែលចាំបាច់សម្រាប់ការនេះ ហើយមិនត្រូវការខ្សែសង្វាក់ផ្លូវដង្ហើម mitochondrial ទេ។ ATP នៅក្នុងដំណើរការនៃការ glycolysis anaerobic ត្រូវបានបង្កើតឡើងដោយសារតែប្រតិកម្មនៃ phosphorylation ស្រទាប់ខាងក្រោម។
នៅក្នុងផ្សិតនៅក្រោមលក្ខខណ្ឌ anaerobic ដំណើរការស្រដៀងគ្នាមួយកើតឡើង - ជាតិអាល់កុល fermentation ក្នុងករណីនេះអាស៊ីត pyruvic ត្រូវបាន decarboxylated ដើម្បីបង្កើត acetaldehyde ដែលបន្ទាប់មកត្រូវបានកាត់បន្ថយទៅជាជាតិអាល់កុល ethyl:
CH 3 -CO-COOH → CH 3 -CHO + CO 2;
CH 3 -CHO + OVERHAND + H + → CH 3 -CH 2 -OH + OVER + ។

Fig.28 ។ ដ្យាក្រាមនៃជាតិស្ករ glycolysis anaerobic
១០.៦. ការបំបែកគ្លុយកូសតាមអាកាស
ការបំបែកគ្លុយកូសតាមបែប Aerobic មានបីដំណាក់កាល៖
1) ការបំប្លែងជាតិស្ករទៅជាអាស៊ីត pyruvic (pyruvate) - aerobic glycolysis ។ ផ្នែកនេះគឺស្រដៀងទៅនឹងដំណើរការនៃ glycolysis anaerobic ដែលបានពិភាក្សាខាងលើ លើកលែងតែជំហានចុងក្រោយរបស់វា (ការបំប្លែង pyruvate ទៅជាអាស៊ីតឡាក់ទិក);
2) ផ្លូវទូទៅនៃ catabolism;
3) ខ្សែសង្វាក់ដឹកជញ្ជូនអេឡិចត្រុង mitochondrial - ដំណើរការនៃការដកដង្ហើមជាលិកា។
ផ្លូវទូទៅនៃ catabolism
ផ្លូវទូទៅនៃ catabolism មានពីរដំណាក់កាល។
ដំណាក់កាលទី 1 - decarboxylation អុកស៊ីតកម្មនៃអាស៊ីត pyruvic ។ នេះគឺជាដំណើរការពហុដំណាក់កាលស្មុគ្រស្មាញដែលជំរុញដោយប្រព័ន្ធពហុអង់ស៊ីម - ស្មុគស្មាញ pyruvate dehydrogenase; បានធ្វើមូលដ្ឋានីយកម្មនៅក្នុង mitochondria (ភ្នាសខាងក្នុងនិងម៉ាទ្រីស) ហើយអាចត្រូវបានបញ្ជាក់ដោយគ្រោងការណ៍ទូទៅសរុប:
CH 3 -CO-COOH + HS-KoA + OVER + → CH 3 -CO-SkoA + OVERHAND + H + + CO 2 ។
ដំណាក់កាលទី 2 - វដ្ត Krebs (វដ្ត citrate ឬវដ្តនៃអាស៊ីត tricarboxylic និង dicarboxylic) (រូបភាព 29); បានធ្វើមូលដ្ឋានីយកម្មនៅក្នុង mitochondria (ម៉ាទ្រីស) ។ នៅក្នុងវដ្ដនេះ សំណល់អាសេទីលដែលរួមបញ្ចូលក្នុងអាសេទីល-CoA បង្កើតបានជាម្ចាស់ជំនួយអ៊ីដ្រូសែនបឋមមួយចំនួន។ លើសពីនេះ អ៊ីដ្រូសែន ដោយមានការចូលរួមពី dehydrogenases ចូលទៅក្នុងសង្វាក់ផ្លូវដង្ហើម។ ជាលទ្ធផលនៃសកម្មភាពរួមគ្នានៃវដ្ត citrate និងខ្សែសង្វាក់ផ្លូវដង្ហើម សំណល់អាសេទីលត្រូវបានកត់សុីទៅជា CO 2 និង H 2 O។ សមីការរួមសម្រាប់លំដាប់ទាំងមូលនៃការផ្លាស់ប្តូរជាតិស្ករអំឡុងពេលការពុកផុយតាមអាកាសមានដូចខាងក្រោម៖
C 6 H 12 O 2 + 6O 2 → 6CO 2 + 6H 2 O
ឥទ្ធិពលថាមពលនៃការបំបែកតាមអាកាសគឺជាការសំយោគនៃម៉ូលេគុល ATP ចំនួន ៣៨ កំឡុងពេលបំបែកម៉ូលេគុលគ្លុយកូស ១ ។ ដូច្នេះនៅក្នុងលក្ខខណ្ឌនៃថាមពលការកត់សុីពេញលេញនៃជាតិស្ករទៅកាបូនឌីអុកស៊ីតនិងទឹកគឺជាដំណើរការដ៏មានប្រសិទ្ធិភាពជាង glycolysis anaerobic ។ អុកស៊ីសែនរារាំង glycolysis anaerobic ដូច្នេះនៅក្នុងវត្តមាននៃអុកស៊ីសែនលើសការផ្លាស់ប្តូរត្រូវបានអង្កេតនៅក្នុងជាលិការុក្ខជាតិនិងសត្វពី glycolysis anaerobic (fermentation) ទៅផ្លូវដង្ហើម (aerobic glycolysis) ពោលគឺឧ។ ការផ្លាស់ប្តូរកោសិកាទៅជាវិធីដែលមានប្រសិទ្ធភាព និងសន្សំសំចៃជាងក្នុងការទទួលបានថាមពល (បែបផែន Pasteur)។ តួនាទីនៃ glycolysis anaerobic ក្នុងការផ្តល់ឱ្យរាងកាយនូវថាមពលគឺអស្ចារ្យជាពិសេសក្នុងអំឡុងពេលការងារដែលពឹងផ្អែករយៈពេលខ្លីនៅពេលដែលថាមពលនៃយន្តការនៃការដឹកជញ្ជូនអុកស៊ីសែនទៅ mitochondria មិនគ្រប់គ្រាន់ដើម្បីផ្តល់នូវ aerobic glycolysis ។ ដូច្នេះការរត់សម្រាប់ ~ 30 វិនាទី (សម្រាប់ 200 ម៉ែត្រ) ត្រូវបានផ្តល់ទាំងស្រុងដោយ glycolysis anaerobic ខណៈពេលដែលអត្រានៃ glycolysis anaerobic ថយចុះជាមួយនឹងការកើនឡើងនៃការដកដង្ហើមហើយអត្រានៃការពុកផុយ aerobic កើនឡើង។ បន្ទាប់ពី 4-5 នាទី។ ការរត់ (1.5 គីឡូម៉ែត្រ) - ពាក់កណ្តាលនៃថាមពលត្រូវបានផ្តល់ដោយ anaerobic ពាក់កណ្តាលដោយដំណើរការ aerobic ។ បន្ទាប់ពី 30 នាទី។ (រត់ 10 គីឡូម៉ែត្រ) - ថាមពលត្រូវបានផ្គត់ផ្គង់ស្ទើរតែទាំងស្រុងដោយដំណើរការ aerobic ។
កោសិកាឈាមក្រហមមិនមាន mitochondria ទាល់តែសោះ ហើយតម្រូវការរបស់ពួកគេសម្រាប់ ATP ត្រូវបានពេញចិត្តទាំងស្រុងដោយ anaerobic glycolysis ។
(ពីភាសាក្រិច glykys - ផ្អែមនិង lysis - រលួយការរលួយ) - មួយក្នុងចំណោមបីសំខាន់ (glycolysis, វដ្ត Krebs និងផ្លូវ Entner-Doudorov) នៃការបង្កើតថាមពលនៅក្នុងសារពាង្គកាយមានជីវិត។ នេះគឺជាដំណើរការនៃ anaerobic (ពោលគឺមិនតម្រូវឱ្យមានការចូលរួមនៃ O 2 ដោយឥតគិតថ្លៃ) ការបំបែកអង់ស៊ីមដែលមិនមែនជាអ៊ីដ្រូលីកនៃកាបូអ៊ីដ្រាត (ជាចម្បងគ្លុយកូសនិង glycogen) នៅក្នុងជាលិកាសត្វដែលអមដោយការសំយោគនៃអាស៊ីត adenosine triphosphoric (ATP) និងបញ្ចប់ដោយ ការបង្កើតអាស៊ីតឡាក់ទិក។ Glycolysis មានសារៈសំខាន់សម្រាប់កោសិកាសាច់ដុំ មេជីវិតឈ្មោល ជាលិកាលូតលាស់ (រួមទាំងដុំសាច់) ដោយសារតែ ផ្តល់នូវការផ្ទុកថាមពលនៅពេលអវត្ដមាននៃអុកស៊ីសែន។ ប៉ុន្តែ glycolysis ត្រូវបានគេស្គាល់ផងដែរនៅក្នុងវត្តមាននៃ O 2 (aerobic glycolysis) - នៅក្នុង erythrocytes, retina, ជាលិកាទារកភ្លាមៗបន្ទាប់ពីកំណើតនិងនៅក្នុង mucosa ពោះវៀន។ G. និង K. Corey ក៏ដូចជាអ្នកត្រួសត្រាយនៃជីវគីមីដូចជា O. Meyerhof និង G. Embden បានរួមចំណែកយ៉ាងធំធេងក្នុងការសិក្សាអំពី glycolysis ។ Glycolysis គឺជាលំដាប់ដំបូងដែលត្រូវបានបកស្រាយយ៉ាងពេញលេញនៃប្រតិកម្មជីវគីមី (ពីចុងសតវត្សទី 19 ដល់ឆ្នាំ 1940) ។ ផ្លូវ hexomonophosphate shunt ឬ pentose phosphate pathway នៅក្នុងកោសិកាមួយចំនួន (erythrocytes, adipose tissue) ក៏អាចដើរតួជាអ្នកផ្គត់ផ្គង់ថាមពលផងដែរ។
បន្ថែមពីលើជាតិស្ករ glycerol អាស៊ីតអាមីណូមួយចំនួន និងស្រទាប់ខាងក្រោមផ្សេងទៀតអាចចូលរួមក្នុងដំណើរការនៃ glycolysis ។ នៅក្នុងជាលិកាសាច់ដុំដែលស្រទាប់ខាងក្រោមសំខាន់នៃ glycolysis គឺ glycogen ដំណើរការចាប់ផ្តើមដោយប្រតិកម្ម 2 និង 3 ( សង់ទីម៉ែត. គ្រោងការណ៍) និងត្រូវបានគេហៅថា glycogenolysis ។ កម្រិតមធ្យមទូទៅសម្រាប់ glycogenolysis និង glycolysis គឺគ្លុយកូស-6-phosphate ។ ផ្លូវបញ្ច្រាសនៃការបង្កើត glycogen ត្រូវបានគេហៅថា glycogenesis ។
ផលិតផលដែលបានបង្កើតឡើងកំឡុងពេល glycolysis គឺជាស្រទាប់ខាងក្រោមសម្រាប់ការបំលែងអុកស៊ីតកម្មជាបន្តបន្ទាប់ ( សង់ទីម៉ែត. វដ្តអាស៊ីត Tricarboxylic ឬវដ្ត Krebs) ។ ដំណើរការស្រដៀងទៅនឹង glycolysis គឺអាស៊ីត lactic, butyric, ជាតិអាល់កុល, glycerol fermentation កើតឡើងនៅក្នុងកោសិការុក្ខជាតិ ផ្សិត និងបាក់តេរី។ អាំងតង់ស៊ីតេនៃដំណាក់កាលនីមួយៗនៃ glycolysis អាស្រ័យលើអាស៊ីត - pH - pH (pH ល្អបំផុត 7-8) សីតុណ្ហភាពនិងសមាសធាតុអ៊ីយ៉ុងនៃឧបករណ៍ផ្ទុក។ លំដាប់នៃប្រតិកម្ម glycolysis ( សង់ទីម៉ែត។គ្រោងការណ៍) ត្រូវបានសិក្សាយ៉ាងល្អហើយផលិតផលកម្រិតមធ្យមត្រូវបានកំណត់។ អង់ស៊ីម glycolysis រលាយដែលមាននៅក្នុងបឹងទន្លេសាបត្រូវបានញែកដាច់ពីគ្នាក្នុងទម្រង់ជាគ្រីស្តាល់ ឬបន្សុត។
អង់ស៊ីមដែលអនុវត្តជំហាននីមួយៗនៃ glycolysis៖
1. Hexokinase CF2.7.1.1 (ឬ glucokinase CF2.7.1.2)
2. Glycogen phosphorylase CP2.4.1.1
3. Phosphoglucomutase CP2.7.5.1
4. គ្លុយកូស phosphate isomerase CP5.3.1.9
5. Phosphofructokinase KF2.7.1.11
6. Fructose bisphosphate aldolase EC4.1.2.13
7. Triose phosphate isomerase CP5.3.1.1
8, 9. Glyceraldehyde phosphate dehydrogenase CP1.2.1.12
10. Phosphoglycerate kinase CP2.7.2.3
11. Phosphoglyceromutase EC2.7.5.3
12. Enolase KF4.2.1.11
13. Pyruvate kinase KF2.7.1.40
14. Lactate dehydrogenase CP1.1.1.27
Glycolysis ចាប់ផ្តើមជាមួយនឹងការបង្កើតដេរីវេនៃផូស្វ័រនៃជាតិស្ករ ដែលរួមចំណែកដល់ការបំប្លែងទម្រង់រង្វិលនៃស្រទាប់ខាងក្រោមទៅជា acyclic ដែលមានប្រតិកម្មច្រើនជាងមុន។ ប្រតិកម្មមួយក្នុងចំណោមប្រតិកម្មដែលគ្រប់គ្រងអត្រានៃ glycolysis គឺប្រតិកម្ម 2 ជំរុញដោយអង់ស៊ីម phosphorylase ។ តួនាទីគ្រប់គ្រងកណ្តាលនៅក្នុង glycolysis ជាកម្មសិទ្ធិរបស់អង់ស៊ីម phosphofructokinase (ប្រតិកម្មទី 5) ដែលសកម្មភាពរបស់វាត្រូវបានរារាំងដោយ ATP និង citrate ប៉ុន្តែត្រូវបានជំរុញដោយផលិតផលពុកផុយរបស់វា។ តំណភ្ជាប់កណ្តាលនៃ glycolysis គឺ glycolytic oxidoreduction (ប្រតិកម្ម 8-10) ដែលជាដំណើរការ redox ដែលដំណើរការជាមួយនឹងការកត់សុីនៃ 3-phosphoglyceraldehyde ទៅអាស៊ីត 3-phosphoglyceric និងការថយចុះនៃ coenzyme nicotinamide adenine dinucleotide (NAD) ។ ការផ្លាស់ប្តូរទាំងនេះត្រូវបានអនុវត្តដោយ 3-phosphoglyceraldehyde dehydrogenase (DPHA) ដោយមានការចូលរួមពី phosphoglycerate kinase ។ នេះគឺជាជំហានអុកស៊ីតកម្មតែមួយគត់នៅក្នុង glycolysis ប៉ុន្តែវាមិនត្រូវការអុកស៊ីសែនដោយឥតគិតថ្លៃទេមានតែវត្តមានរបស់ NAD + ប៉ុណ្ណោះដែលចាំបាច់ដែលបន្ទាប់មកត្រូវបានកាត់បន្ថយទៅជា NAD-H 2 ។
ជាលទ្ធផលនៃ oxidoreduction (ដំណើរការ redox) ថាមពលត្រូវបានបញ្ចេញដែលត្រូវបានបង្គរ (ក្នុងទម្រង់ជាសមាសធាតុ ATP ដែលសំបូរទៅដោយថាមពល) នៅក្នុងដំណើរការនៃការ phosphorylation ស្រទាប់ខាងក្រោម។ ប្រតិកម្មទីពីរដែលផ្តល់នូវការបង្កើត ATP គឺប្រតិកម្ម 13 - ការបង្កើតអាស៊ីត pyruvic ។ នៅក្រោមលក្ខខណ្ឌ anaerobic glycolysis បញ្ចប់ដោយការបង្កើតអាស៊ីតឡាក់ទិក (ប្រតិកម្ម 14) នៅក្រោមសកម្មភាពនៃ lactate dehydrogenase និងដោយមានការចូលរួមពី NAD ដែលត្រូវបានកាត់បន្ថយដែលត្រូវបានកត់សុីទៅ NAD (NAD-H 2) ហើយអាចប្រើម្តងទៀតនៅដំណាក់កាលអុកស៊ីតកម្ម។ . នៅក្រោមលក្ខខណ្ឌ aerobic អាស៊ីត pyruvic ត្រូវបានកត់សុីនៅក្នុង mitochondria ក្នុងអំឡុងពេលវដ្ត Krebs ។
ដូច្នេះនៅពេលដែល 1 ម៉ូលេគុលនៃជាតិស្ករត្រូវបានបំបែក 2 ម៉ូលេគុលនៃអាស៊ីតឡាក់ទិក និង 4 ម៉ូលេគុលនៃ ATP ត្រូវបានបង្កើតឡើង។ ក្នុងពេលជាមួយគ្នានេះក្នុងដំណាក់កាលដំបូងនៃ glycolysis (សូមមើលប្រតិកម្ម 1, 5) 2 ម៉ូលេគុល ATP ត្រូវបានប្រើប្រាស់ក្នុង 1 ម៉ូលេគុលគ្លុយកូស។ នៅក្នុងដំណើរការនៃការ glycogenolysis ម៉ូលេគុល ATP 3 ត្រូវបានបង្កើតឡើងដោយសារតែ។ មិនចាំបាច់ចំណាយ ATP ដើម្បីទទួលបានគ្លុយកូស-6-ផូស្វាត។ ប្រតិកម្មប្រាំបួនដំបូងនៃ glycolysis គឺជាដំណាក់កាល endergonic (ជាមួយនឹងការស្រូបយកថាមពល) ហើយប្រតិកម្មចុងក្រោយគឺដំណាក់កាល exergonic (ជាមួយនឹងការបញ្ចេញថាមពល) ។ នៅក្នុងដំណើរការនៃការ glycolysis មានតែប្រហែល 7% នៃថាមពលទ្រឹស្តីត្រូវបានបញ្ចេញដែលអាចទទួលបានជាមួយនឹងការកត់សុីពេញលេញនៃជាតិស្ករ (ទៅ CO 2 និង H 2 O) ។ ទោះជាយ៉ាងណាក៏ដោយ ប្រសិទ្ធភាពនៃការផ្ទុកថាមពលសរុបក្នុងទម្រង់ ATP គឺ 35-40% ហើយនៅក្រោមលក្ខខណ្ឌជាក់ស្តែង កោសិកាអាចខ្ពស់ជាងនេះ។
Glyceraldehyde phosphate dehydrogenase និង lactate dehydrogenase ត្រូវបានផ្សំពីខាងក្នុង (មួយត្រូវការ NAD + ទម្រង់ផ្សេងទៀត NAD +) ដែលធានាចរាចរនៃ coenzyme នេះ។ នេះប្រហែលជាសារៈសំខាន់ជីវគីមីសំខាន់នៃស្ថានីយ dehydrogenase ។
ប្រតិកម្ម glycolysis ទាំងអស់គឺអាចបញ្ច្រាស់បាន លើកលែងតែ 1, 5, និង 13។ ទោះជាយ៉ាងណាក៏ដោយ គ្លុយកូស (ប្រតិកម្ម 1) ឬ fructose monophosphate (ប្រតិកម្ម 5) អាចទទួលបានពីដេរីវេនៃផូស្វ័ររបស់ពួកគេដោយការបំបែក hydrolytic នៃអាស៊ីតផូស្វ័រនៅក្នុងវត្តមាននៃអង់ស៊ីមសមស្រប។ ប្រតិកម្ម 13 គឺមិនអាចត្រឡប់វិញបានជាក់ស្តែងដោយសារតែថាមពលខ្ពស់នៃអ៊ីដ្រូលីលីសនៃក្រុមផូស្វ័រ (ប្រហែល 13 kcal / mol) ។ ដូច្នេះការបង្កើតជាតិស្ករពីផលិតផល glycolysis ដំណើរការតាមរបៀបផ្សេង។
នៅក្នុងវត្តមាននៃ O 2 អត្រានៃ glycolysis ថយចុះ (ឥទ្ធិពល Pasteur) ។ មានឧទាហរណ៍នៃការទប់ស្កាត់ការដកដង្ហើមជាលិកាដោយ glycolysis (ឥទ្ធិពល Crabtree) នៅក្នុងជាលិកា glycolyzing ខ្លាំងមួយចំនួន។ យន្តការនៃទំនាក់ទំនងរវាងដំណើរការអុកស៊ីតកម្ម anaerobic និង aerobic មិនត្រូវបានយល់ច្បាស់នោះទេ។ បទប្បញ្ញត្តិដំណាលគ្នានៃដំណើរការនៃ glycolysis និង glycogenesis កំណត់លំហូរនៃកាបូននៅតាមបណ្តោយផ្លូវនីមួយៗ អាស្រ័យលើតម្រូវការរបស់រាងកាយ។ ការគ្រប់គ្រងត្រូវបានអនុវត្តជាពីរកម្រិត - អ័រម៉ូន (នៅក្នុងសត្វខ្ពស់តាមរយៈបទប្បញ្ញត្តិដែលមានការចូលរួមពីអ្នកនាំសារទីពីរ) និងការរំលាយអាហារ (នៅក្នុងគ្រប់សារពាង្គកាយទាំងអស់) ។
Igor Rapanovich