See ei ole komplemendisüsteemi funktsioon. Komplemendisüsteemi valgud: omadused ja bioloogiline aktiivsus. Komplemendisüsteemi aktiveerimise klassikaline viis
Komplement on keha üks olulisemaid polüfunktsionaalseid süsteeme. Ühest küljest võib seda pidada antikehast sõltuvate reaktsioonide peamiseks efektoriks. See osaleb mitte ainult lüütilistes ja bakteritsiidsetes reaktsioonides, vaid ka muudes antikehadest sõltuvates toimetes, mille hulgas on fagotsütoosi suurenemine üks selle olulisemaid funktsioone in vivo. Teisest küljest toimib komplement põhisüsteemina - põletikuliste reaktsioonide võimendajana. Võimalik, et evolutsioonilisest aspektist on see tema peamine (esmane) funktsioon ja seda pole üldse vaja seostada antikehade ja muude immunoloogiliste mehhanismidega.
Komplemendi aktiveerimise protsessi keskne sündmus on C3 komponendi lõhustumine mööda klassikalist (nii nimetatakse ainult seetõttu, et see avastati esimesena, mitte selle erakordse tähtsuse tõttu) ja alternatiivset rada. Teine põhipunkt on protsessi võimalik sügavus: see peatub
kas see on SZ lõhustumise staadiumis, pakkudes samal ajal mitmeid bioloogilisi mõjusid, või süveneb see veelgi (C5-lt C9-le). Aktiveerimise viimast etappi nimetatakse sageli terminaalseks, lõplikuks (membraani rünnak), see on tavaline, identne klassikalise ja alternatiivse radade jaoks ning sellega on seotud komplemendi lüütiline funktsioon.
Praegu on komplemendisüsteemi ühendatud vähemalt 20 plasmavalku. Põhimõtteliselt jagunevad nad 3 rühma. Klassikalises aktivatsioonirajas ja viimases (membraanirünnaku) etapis osalevad komponendid on tähistatud kui Clq, Clr, C1, C4, C2, C3, C5, C6, C7, C8 ja C9. Alternatiivses aktivatsioonirajas osalevaid valke nimetatakse faktoriteks ja neid tähistatakse kui B, D, P. Lõpuks eristatakse reaktsiooni intensiivsust reguleerivate valkude rühma või kontrollvalkude rühma: nende hulka kuuluvad C1-inhibiitor. (C1INH), C3b-inaktivaator (C3bINa ), pIH faktor - C4 - BP, anafülotoksiini inhibiitor. Põhikomponentide ensümaatilisel lõhustamisel tekkinud fragmendid tähistatakse väikeste tähtedega (näiteks C3a, C3b, C3d, C5a jne). Ensümaatilise aktiivsusega komponentide või fragmentide tähistamiseks asetatakse nende sümbolite kohale joon, näiteks Cl, C42, C3bBb.
Komplemendi üksikute komponentide sisaldus seerumis on järgmine:
Komponendi kontsentratsioon, μg / ml
Klassikaline viis
C1 70
C1 34
C1 31
C4 600
C2 25
SZ 1200
Alternatiivne viis
Properdin 25
Tegur B 200
Tegur D 1
Membraanirünnaku kompleks
C5 85
C6 75
C7 55
C8 55
C9 60
Reguleerivad valgud
C1 inhibiitor 180
Tegur H 500
Tegur I 34
Komplemendi süsteem on üks "käivitavatest" ensüümidest
keemilised süsteemid, samuti vere hüübimissüsteem, fibrinolüüs, kiniinide moodustumine. Seda iseloomustab kiire ja kiiresti kasvav reaktsioon stimulatsioonile. Seda võimendust (amplifikatsiooni) põhjustab kaskaadnähtus, mille puhul on iseloomulik, et ühe reaktsiooni saadused on järgmise katalüsaatoriks. Selline kaskaad võib olla lineaarne, ühesuunaline (näiteks klassikaline komplemendi aktiveerimise rada) või see hõlmab tagasisideahelaid (alternatiivne rada). Seega toimuvad mõlemad variandid komplemendisüsteemis (skeem 1).
Klassikalist rada aktiveerivad immuunkompleksid
antigeen – antikeha, mille hulka kuuluvad antigeenidena IgM, IgG (alaklassid 3, 1, 2; need paiknevad väheneva aktiivsusega). Lisaks võivad IgG, CRP, DNA ja plasmiini agregaadid aktiveerida klassikalise raja. Protsess algab C1 aktiveerimisega, mis koosneb 3 komponendist Clq, Clr, Cls. Clq (suhteline molekulmass 400), on omapärase struktuuriga: 6 alaühikut kollageenipulga ja mittekollageenipeaga, molekuli pea vastas olevas otsas on ühendatud 6 varrast. Peadel on kohad antikehamolekulide külge kinnitamiseks, samas kui C1G ja Cls kinnituskohad asuvad kollageenivarrastel. Pärast Clq kinnitumist AT-ga muutub C1r konformatiivsete transformatsioonide kaudu aktiivseks proteaasiks C1r. lõikab Cls, muutes kogu kompleksi C1 serinesteraasiks. Viimane lõikab C4 kaheks fragmendiks - C4a ja C4b ning C2 C2a ja C2b. Saadud C4b2b kompleks (a) on aktiivne ensüüm, mis lagundab C3 komponendi (klassikalise raja C3 konvertaas); mõnikord on see tähistatud C42.
Klassikalise raja regulaatoriks on C1 inhibiitor (C1INH), mis pärsib C1r ja Cls aktiivsust, seondudes pöördumatult nende ensüümidega. Leiti, et C1INH vähendab ka kallikreiini, plasmiini ja Hagemani faktori aktiivsust. Selle inhibiitori kaasasündinud puudulikkus põhjustab C4 ja C2 kontrollimatut aktiveerumist, mis väljendub kaasasündinud tursevastases vormis.
Alternatiivne (properdiniline) rada koosneb järjestikustest reaktsioonidest, mis ei sisalda Cl-, C4- ja C2-komponente ning viivad sellegipoolest C3 aktiveerimiseni. Lisaks põhjustavad need reaktsioonid lõpliku membraanirünnaku mehhanismi aktiveerimist. Selle raja aktiveerimise algatab gramnegatiivsete bakterite endotoksiin, mõned polüsahhariidid nagu inuliin ja sümosaan, IgA või IgG sisaldavad immuunkompleksid (IC) ning mõned bakterid ja seened (nt Staf. Epidermis, Candida albicans). Reaktsioon hõlmab 4 komponenti: tegurid D ja B, C3 ja propediin (P). Sel juhul on faktor D (ensüüm) sarnane klassikalise raja Cls-iga, C3 ja faktor B on sarnased C4 ja C2 komponentidega. Selle tulemusena moodustub alternatiivse raja C3Bb konvertaas. Saadud kompleks on äärmiselt ebastabiilne ja oma funktsiooni täitmiseks stabiliseerib seda propediin, moodustades keerukama CZvVlR kompleksi. Alternatiivse raja reguleerivad valgud on piH ja C3J-inaktivaator (C3JNA). Esimene seostub C3b-ga ja moodustab inaktivaatori (C3bINA) sidumissaidi. Nende tegurite või nende geneetilise puudulikkuse kunstlik eemaldamine, mille olemasolu inimestel on hiljuti kindlaks tehtud, viib alternatiivse raja ohjeldamatu aktiveerimiseni, mis võib potentsiaalselt põhjustada CZ või faktori B täielikku ammendumist.
Terminalmembraani rünnaku mehhanism. Nagu juba mainitud, koonduvad mõlemad teed C3 komponendile, mille aktiveerib mis tahes moodustunud C42 või C3bBb konvertaas. Sest
C5-konvertaasi moodustumine nõuab täiendava koguse C3 lõhustamist. C3, mis on seotud raku pinnal, ja vaba B, P või p1H moodustavad saidi C5 sidumiseks ja muudavad viimase vastuvõtlikuks mis tahes C3 konvertaasi proteolüüsi suhtes. Sel juhul lõigatakse C5-st väike peptiid C5a ja ülejäänud suur C5b kinnitub rakumembraanile ja sellel on Cb kinnituskoht. Lisaks on komponendid C7, C8, C9 järjestikku ühendatud. Selle tulemusena moodustub stabiilne transmembraanne kanal, mis tagab ioonide ja vee kahesuunalise liikumise läbi raku bilipiidkihi. Membraan on kahjustatud ja rakk sureb. Seega tapetakse eelkõige võõraid mikroorganisme.
Komplemendi aktiveerimise käigus moodustub hulk fragmente, peptiide, mis mängivad olulist rolli põletiku-, fagotsütoosi- ja allergiliste reaktsioonide protsessides.
Seega põhjustab C4 ja C2 lõhustamine Cls-iga veresoonte läbilaskvuse suurenemist ja on C1 inhibiitori puudulikkusega seotud kaasasündinud tursevastase patogeneesi aluseks. Peptiididel C3a ja C5a on anafülotoksiini omadused. Seoses nuumrakkude ja basofiilidega, kutsuvad nad esile histamiini vabanemise. Trombotsüütidega seondudes põhjustab SZa serotoniini sekretsiooni. C3a ja C5a anafülotoksiline toime hävib kergesti karboksüpeptidaasi B toimel, mis lõikab nendest peptiididest arginiini. Saadud tooted omandavad polümorfonukleaarsete rakkude, eosinofiilide ja monotsüütide suhtes kemoatraktantide omadused. C5i67 kompleks, millel puuduvad hemolüütilised omadused, ja Bb fragment põhjustavad kemotaksist ainult polümorfonukleaarsetes leukotsüütides. Tavaline inimese seerum sisaldab CFi-d, mis pärsib C5a aktiivsust polümorfonukleaarsete rakkude suhtes, kõrvaldades selle võime stimuleerida lüsosomaalsete ensüümide vabanemist. Sarkoidoosi ja Hodgkini tõvega patsientidel on CFi liigne. See võib seletada nende rakkude talitlushäireid. Teine peptiid, C3b, on polümorfonukleaarsete rakkude (PMN) ja makrofaagide jaoks tugev opsoniin. Selle peptiidi retseptoreid on leitud ka teistelt rakkudelt (monotsüüdid ja B-lümfotsüüdid), kuid nende tähtsus nende rakkude funktsioneerimisele on siiani ebaselge. Komplemendi sidumine lümfotsüütidega, mis on osa immuunkompleksist, võib mängida rolli esmase immuunvastuse kujunemisel.
Komplemendisüsteemi uuringut kliinilises praktikas saab kasutada haiguse diagnoosimiseks, protsessi aktiivsuse määramiseks ja teraapia efektiivsuse hindamiseks. Vadaku komplemendi tase igal ajahetkel sõltub selle komponentide sünteesi, katabolismi ja tarbimise tasakaalust.
Komplemendi hemolüütilise aktiivsuse madalad väärtused võivad kajastada üksikute komponentide puudulikkust või selle lõhustumisproduktide esinemist vereringes. Seda tuleks ka meeles pidada
et komplemendi intensiivset lokaalset tarbimist sellistes piirkondades nagu pleura, liigeseõõnsused ei tohi kombineerida komplemendi ja seerumi taseme muutusega. Näiteks mõnel reumatoidartriidiga patsiendil võib seerumi komplemendi tase olla normaalne, samas kui sünoviaalvedelikus võib see selle aktiivse tarbimise tõttu järsult väheneda. Komplemendi määramine sünoviaalvedelikus on diagnoosimiseks väga oluline.
Kaasasündinud komplemendi puudused. Komplemendi puudulikkuse pärand on autosoomne retsessiivne või kodominantne, seega on heterosügootidel umbes 50% komplemendi komponentide normaalsest tasemest. Enamikul juhtudel seostatakse süsteemse erütematoosluupusega kaasasündinud varajase alguse komponentide (C1, C4, C2) puudusi. C-komponendi puudulikkusega isikud on altid korduvatele püogeensetele infektsioonidele. Terminaalsete komponentide puudulikkusega kaasneb tundlikkuse suurenemine gonokokkide ja meningokokkide infektsioonide suhtes. Süsteemne erütematoosluupus esineb ka nende komplemendi puudulikkusega, kuid harvemini. Kõige sagedamini täheldatakse kaasasündinud C2 puudulikkust. Selle tunnuse homosügootset puudulikkust leitakse mõnede autoimmuunhaiguste, sealhulgas luupuselaadsete haiguste, Shenleini-Henochi tõve, glomerulonefriidi ja dermatomüosiidi korral. Selle tunnuse suhtes homosügootsetel isikutel ei ole suurenenud vastuvõtlikkus infektsioonidele, kui alternatiivne aktiveerimisrada toimib normaalselt. C2 puudulikkusega homosügoote on leitud praktiliselt tervete inimeste seas.
Heterosügootne C2 puudulikkus võib olla seotud juveniilse reumatoidartriidi ja süsteemse erütematoosluupusega. Pereuuringud on leidnud, et C2 ja C4 puudused on seotud teatud HLA haplotüüpidega.
Komplemendisüsteemi regulatoorsete valkude puudulikkusel võivad olla ka kliinilised ilmingud. Seega täheldatakse C3INA kaasasündinud puudulikkuse korral kliinilist pilti, mis sarnaneb C3 puudulikkusega, kuna viimase tarbimine alternatiivse tee kaudu muutub kontrollimatuks.
8381 0
Komplemendi süsteem, mis koosneb umbes 30 valgust, mis nii ringlevad kui ka ekspresseeritakse membraanil, on nii kaasasündinud kui ka antikehade poolt vahendatud omandatud immuunvastuste oluline efektorharu. Mõiste "komplement" tulenes asjaolust, et leiti, et see temperatuuritundlik vereseerumi materjal "täiendab" antikehade võimet tappa baktereid. On teada, et komplement mängib olulist rolli kaitsmisel paljude nakkuslike mikroorganismide eest.
Selle kaitsefunktsiooni kõige olulisemad komponendid on: 1) opsoniinide tootmine - molekulid, mis suurendavad makrofaagide ja neutrofiilide fagotsütoosi võimet; 2) anafülatoksiinide tootmine - peptiidid, mis kutsuvad esile lokaalseid ja süsteemseid põletikulisi reaktsioone; 3) mikroorganismide otsene hävitamine.
On teada ka teisi olulisi komplemendi funktsioone, nagu antigeenispetsiifiliste immuunvastuste tugevdamine ja homöostaasi (kehasisese stabiilsuse) säilitamine immuunkomplekside ja surnud või surevate rakkude eemaldamise kaudu. Teame ka, et komplemendi aktiveerimise kontrolli all hoidmine võib kahjustada rakke ja kudesid kehas.
Komplemendi komponendid sünteesitakse maksas, samuti põletikulises reaktsioonis osalevate rakkude poolt. Kõikide komplementvalkude kontsentratsioon ringlevas veres on ligikaudu 3 mg/ml. (Võrdluseks: IgG kontsentratsioon veres on ligikaudu 12 mg/ml) Mõnede komplemendi komponentide kontsentratsioonid on kõrged (näiteks C3 puhul umbes 1 mg/ml), samas kui teiste komponentide (nt faktor D ja C2) kontsentratsioon on kõrge. esineb väikestes kogustes ...
Täiendage aktiveerimisradu
Komplemendi aktiveerimise algstaadiumid on selle komponentide järjestikune kaskaadaktiveerimine. Selles etapis kutsub ühe komponendi aktiveerimine esile ensüümi toime, mis viib omakorda järgmise komponendi aktiveerumiseni. Kuna üks aktiivne ensüümi molekul on võimeline lõikama paljusid substraadimolekule, võimendab see reaktsioonide kaskaad suhteliselt nõrka algsignaali. Need komplemendi süsteemi kaskaadomadused on sarnased teiste seerumi kaskaadidega, mille eesmärk on trombide moodustumine ja kiniinide, vaskulaarsete põletikuliste vahendajate tootmine.Pärast aktiveerimist jagatakse üksikud komponendid fragmentideks, mida tähistatakse väiketähtedega. Lõhustatud fragmentidest väiksemat tähistatakse tavaliselt tähega "a", suuremat tähega "b". Ajalooliselt nimetatakse aga suuremat lõhustatud C2 fragmenti tavaliselt C2a-ks ja väiksemat C2b-ks. (Mõnes tekstis ja artiklis on aga C2 komplemendi komponentide fragmente tähistatud vastupidiselt.) Väikeste tähtedega tähistatakse ka edasisi dekolteefragmente, näiteks C3d.
Komplemendi aktiveerimiseks on kolm teadaolevat viisi: klassikaline, lektiin ja alternatiiv.
Iga aktivatsioonitee algust iseloomustavad oma komponendid ja äratundmisprotsessid, kuid hilisemates etappides kasutatakse kõigil kolmel juhul samu komponente. Allpool käsitletakse iga aktivatsioonitee omadusi ja neid aktiveerivaid aineid.
Klassikaline viis
Klassikaline aktiveerimistee on saanud sellise nime, kuna see tuvastati esimesena. Klassikalise raja valgukomponendid on tähistatud C1, C2, C9. (Numbrid on paigutatud komponentide avamise järjekorras, mitte nende aktiveerimise järjekorras.) Antigeen-antikeha kompleksid on klassikalise raja peamised aktivaatorid. Seega on viimane humoraalse adaptiivse immuunvastuse aktiveerimise peamine efektorrada.Teised aktivaatorid on teatud viirused, surnud rakud ja rakusisesed membraanid (nt mitokondrid), immunoglobuliinide agregaadid ja β-amüloid, mida leidub Alzheimeri tõve naastudes. C-reaktiivne valk on ägeda faasi valk – põletikuvastuse komponent; see kinnitub paljude bakterite (nt Streptococcus pneumoniae) pinnal ekspresseeritavale polüsahhariidile fosforüülkoliinile ja aktiveerib ka klassikalise raja.
Klassikaline rada käivitub, kui C1 kinnitub antigeeni-antikeha kompleksis oleva antikeha külge, näiteks antikehaga, mis on seotud bakteri pinnal ekspresseeritud antigeeniga (joonis 13.1). Komponent C1 on kolme erineva valgu kompleks: Clq (sisaldab kuut identset alamkomponenti), mis on seotud kahe molekuliga (igaüks kaks) – Clr ja Cls. Kui Cl on aktiveeritud, seonduvad selle globulaarsed piirkonnad - Clq alamkomponendid - kas ühe IgM või kahe antigeeniga seotud IgG molekuli Fc fragmentide Clq-spetsiifilise piirkonnaga (IgG seondumine on näidatud joonisel 13.1).
Seega on IgM ja IgG antikehad tõhusad komplemendi aktivaatorid. Inimese immunoglobuliinid, millel on võime Cl-ga seonduda ja seda aktiveerida, selle võime vähendamise järjekorras, paiknevad: IgM>> IgG3> IgG 1 "IgG2. Immunoglobuliinid IgG4, IgD, IgA ja IgE ei interakteeru Clq-ga, ei fikseeri ega aktiveeri seda, st. ärge aktiveerige komplementi klassikalisel viisil.
Pärast C1 seondumist antigeen-antikeha kompleksiga omandab Cls ensümaatilise aktiivsuse. Seda aktiivset vormi tuntakse Cls esteraasina. Ta jagab klassikalise tee järgmise komponendi - C4 - kaheks osaks: C4a ja C4b. Väiksem osa - C4a - jääb lahustunud olekusse, samas kui C4b seostub kovalentselt bakteri või muu aktiveeriva aine pinnaga.
Rakupinnaga seotud C4b osa seondub seejärel C2-ga, mille Cls lõikab. C2 lõhustamine annab C2b fragmendi, mis jääb lahustunud olekusse, ja C2a. C2a omakorda kinnitub rakupinnal C4b-ga, moodustades C4b2a kompleksi. Seda kompleksi nimetatakse klassikalise raja C3-konvertaasiks, sest nagu hiljem näeme, lagundab see ensüüm järgmise komponendi C3.
Lektiini rada
Lektiini rada aktiveerivad valkude terminaalsed mannoosijäägid ja polüsahhariidid bakterite pinnal. Neid jääke imetajarakkude pinnalt ei leidu, seetõttu võib lektiini rada pidada enese ja võõra äratundmise vahendiks. Kuna see aktiveerimisrada ei nõua antikehade olemasolu, on see osa kaasasündinud immuunkaitsesüsteemist.Joonisel fig. 13.1 näitab, kuidas bakterite mannoosijäägid seonduvad ringleva mannoosi siduva lektiinikompleksiga (MSL; struktuurilt sarnane klassikalise raja Clq-ga) ja kahe seotud proteaasiga, nn. mannoosiga seotud seriini proteaasid (MASP-1 ja -2)... See seondumine aktiveerib MASP-1 klassikalise komplemendi raja komponentide - C4 ja C2 - järgnevaks lõhustamiseks koos C4b2a, klassikalise raja C3-konvertaasi moodustumisega bakterite pinnal. Ja MASP-2-l on võimalus C3 otse lõhustada. Seega on lektiini rada pärast C3 aktiveerimisfaasi sarnane klassikalisele.
Alternatiivne viis
Alternatiivset viisi komplemendi aktiveerimiseks käivitab peaaegu iga võõraine. Enim uuritud ainete hulka kuuluvad lipopolüsahhariidid (LPS, tuntud ka kui endotoksiinid gramnegatiivsete bakterite rakuseinas), teatud pärmseente rakuseinad ja kobra mürgis leiduv valk (kobra mürgifaktor). Mõned ained, mis aktiveerivad klassikalist rada – viirused, immunoglobuliinide agregaadid ja surnud rakud – käivitavad ka alternatiivse raja.Aktiveerimine toimub spetsiifiliste antikehade puudumisel. Seega on komplemendi aktiveerimise alternatiivseks rajaks kaasasündinud immuunkaitsesüsteemi efektorharu. Mõned alternatiivse raja komponendid on sellele ainulaadsed (seerumifaktorid B ja D ning prodiin, tuntud ka kui faktor P), samas kui teised (C3, C3b, C5, C6, C7, C8 ja C9) on klassikalise rajaga ühised.
Komponent C3b ilmub veres väikestes kogustes pärast C3 reaktiivse tioolrühma spontaanset lõhustumist. See “olemasolev” C3b on võimeline seonduma rakupindadel ekspresseeritud valkude ja süsivesikute hüdroksüülrühmadega (vt joonis 13.1). C3b akumuleerumine raku pinnal käivitab alternatiivse raja.
See võib esineda nii võõral kui ka keha enda rakul; seega alternatiivse tee seisukohalt on see alati jooksev. Kuid nagu allpool üksikasjalikumalt näidatud, reguleerivad keha enda rakud alternatiivse raja reaktsioonide kulgu, samal ajal kui võõrad rakud ei oma selliseid reguleerimisvõimeid ja ei saa takistada alternatiivse raja järgnevate sündmuste arengut.
Riis. 13.1. Klassika, lektiini ja alternatiivsete radade käivitamine. Iga raja aktiveerimise ja C3-konvertaasi moodustumise demonstreerimine
Alternatiivse raja järgmises etapis ühineb seerumi valk, faktor B, rakupinnal C3b-ga, moodustades C3bB kompleksi. Seejärel lõhustab faktor D faktor B, mis asub C3bB kompleksis raku pinnal, mille tulemusena moodustub Ba fragment, mis vabaneb ümbritsevasse vedelikku, ja Bb, mis jääb seotuks C3b-ga. See C3bBb on C3. - alternatiivse raja konvertaas, mis lõikab C3 C3a ja C3b.
Tavaliselt C3bBb lahustub kiiresti, kuid võib stabiliseerida, kui seda kombineerida propediiniga (vt joonis 13.1). Selle tulemusena on propidiiniga stabiliseeritud C3bBb võimeline siduma ja lagundama suures koguses C3 väga lühikese aja jooksul. Nende kiiresti moodustunud suures koguses C3b kogunemine raku pinnale viib alternatiivse raja peaaegu "plahvatusliku" käivitamiseni. Seega loob propertydiini seondumine C3bBb-ga alternatiivse raja võimendussilmuse. Profaktiini võimet aktiveerida amplifikatsioonisilmust kontrollib regulatoorsete valkude vastupidine toime. Seega ei toimu alternatiivse raja aktiveerimine kogu aeg.
C3 ja C5 aktiveerimine
C3 lõhustamine on kõigi kolme aktivatsioonitee peamine faas. Joonisel fig. 13.2 näitab, et C3-konvertaasid klassikalisel ja alternatiivsel rajal (vastavalt C4b2a ja C3bBb) lõikavad C3 kaheks fragmendiks. Väiksem C3a on lahustuv valk, mida nimetatakse anafülatoksiiniks: see aktiveerib põletikus osalevaid rakke. Suurem fragment, C3b, jätkab komplemendi kaskaadi aktiveerimise protsessi, seondudes aktiveerimiskohta ümbritsevate rakupindadega. Nagu allpool näidatud, osaleb C3b ka keha kaitses, põletikus ja immuunregulatsioonis.
Riis. 13.2. C3-komponendi lõhustamine C3-konvertaasi ja C5-komponendi lõhustamine C5-konvertaasi poolt klassikalisel ja lektiini (ülemisel) ja alternatiivsel (alumisel) rajal. Kõigil juhtudel lõhustatakse C3 C3b-ks, mis ladestub raku pinnale, ja C3a-ks, mis vabaneb vedelasse keskkonda. Samamoodi lõhustatakse C5 C5b-ks, mis ladestub raku pinnale, ja C5a-ks, mis vabaneb vedelasse keskkonda.
C3b seondumine C3 konvertaasiga nii klassikalisel kui ka alternatiivsel rajal käivitab järgmise komponendi C5 seondumise ja lõhustumise (vt joonis 13.2). Sel põhjusel kuuluvad C3b-ga seotud C3-konvertaasid C5-konvertaaside hulka (C4b2a3b klassikalises rajas; C3bBb3b alternatiivina). C5 lõhustamine annab kaks fragmenti. C5a fragment vabaneb lahustuvas vormis ja on aktiivne anafülatoksiin. C5b fragment seondub rakupinnaga ja moodustab tuuma, et seonduda terminaalse komplemendi komponentidega.
Terminali tee
Komplemendi kaskaadi terminaalsed komponendid – C5b, C6, C7, C8 ja C9 – on ühised kõikidele aktivatsiooniteedele. Nad seostuvad üksteisega ja moodustavad membraanirünnaku kompleksi (MAC), mis põhjustab rakkude lüüsi (joonis 13.3).
Riis. 13.3 Membraanirünnaku kompleksi moodustumine. Hilise faasi komplemendi - C5b-C9 - komponendid on järjestikku ühendatud ja moodustavad raku pinnal kompleksi. Arvukad C9 komponendid kinnituvad selle kompleksi külge ja polümeriseeruvad, moodustades polü-C9, luues kanali, mis läbib rakumembraani
MAC moodustumise esimene faas on C6 kinnitumine C5b-le rakupinnal. Seejärel seondub C7 C5b ja C6-ga ning tungib läbi raku välismembraani. Hilisem C8 seondumine C5b67-ga viib kompleksi moodustumiseni, mis tungib sügavamale rakumembraani. Rakumembraanil toimib C5b-C8 retseptorina C9-le, molekulile nagu perforiin, mis seondub C8-ga.
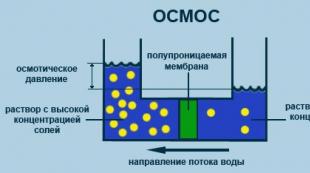
Täiendavad C9 molekulid interakteeruvad kompleksis C9 molekuliga, moodustades polümeriseerunud C9 (polü-C9). Need polü-C9 moodustavad transmembraanse kanali, mis häirib osmootset tasakaalu rakus: ioonid tungivad läbi selle ja vesi siseneb. Rakk paisub, membraan muutub makromolekulidele läbilaskvaks, mis seejärel rakust lahkuvad. Selle tulemusena toimub rakkude lüüs.
R. Koiko, D. Sunshine, E. Benjamini
, Kolloidsüsteemide esteetiline, bioloogiline ja kultuuriline roll, 1. Ohutuse koht ja roll kutsetegevuses ... teha, R&D Raha ja selle roll majanduses.docx, Millist rolli mängib perekond isiksuse kujunemisel .docx, Halperin P.Ya. Vaimse järkjärguline kujunemine action.docx, PR 01 Projektiidee määratlemine. Projekti eesmärkide kujundamine raamistikus, Filosoofia koht ja roll kahekümnenda sajandi kultuuris .. docx.
Komplemendi efektorroll. Membraanirünnaku kompleksi moodustumine ja selle roll raku lüüsis.
a) osaleb mikroobide ja teiste rakkude lüüsis (tsütotoksiline toime);
b) omab kemotaktilist aktiivsust;
c) võtab osa anafülaksiast;
d) osaleb fagotsütoosis.
Komplemendi peamised kasulikud mõjud on järgmised:
abi mikroorganismide hävitamisel;
immuunkomplekside intensiivne eemaldamine;
humoraalse immuunvastuse esilekutsumine ja tugevdamine.
Komplemendi süsteem võib kahjustada oma keha rakke ja kudesid järgmistel juhtudel:
kui selle üldine massiivne aktiveerumine toimub näiteks gramnegatiivsete bakterite põhjustatud septitseemiaga;
kui selle aktiveerumine toimub koenekroosi fookuses, eriti müokardiinfarkti korral;
kui aktiveerumine toimub kudede autoimmuunreaktsiooni käigus.
Esimene faas: C6 kinnitumine C5b-le rakupinnal. Seejärel seondub C7 C5b ja C6-ga ning tungib sisse välimine membraan rakud. Hilisem C8 seondumine C5b67-ga viib kompleksi moodustumiseni, mis tungib sügavamale rakumembraani. Rakumembraanil toimib C5b-C8 retseptorina C9-le, molekulile nagu perforiin, mis seondub C8-ga. Täiendavad C9 molekulid interakteeruvad kompleksis C9 molekuliga, moodustades polümeriseerunud C9 (polü-C9). Need moodustavad transmembraanse kanali, mis rikub rakus osmootset tasakaalu: ioonid tungivad läbi selle ja vesi siseneb. Rakk paisub, membraan muutub makromolekulidele läbilaskvaks, mis seejärel rakust lahkuvad. Selle tulemusena toimub rakkude lüüs.

Komplimentide süsteem - komplekssete valkude kompleks, mis on pidevalt veres olemas. See on kaskaadsüsteem proteolüütilised ensüümid disainitud humoraalne keha kaitsmine võõragentide tegevuse eest, see osaleb selle rakendamises immuunvastus organism. See on nii kaasasündinud kui ka omandatud immuunsuse oluline komponent.

Klassikalisel teel komplemendi aktiveerib antigeen-antikeha kompleks. Selleks piisab, kui antigeeni sidumises osaleb üks IgM molekul või kaks IgG molekuli. Protsess algab C1 komponendi lisamisega AG + AT kompleksilemis laguneb allüksusteksC1q, C1r ja C1s. Lisaks hõlmab reaktsioon järjestikku aktiveeritud "varajase" komplemendi komponente järgmises järjestuses: C4, C2, SZ. C3 komplemendi "varajane" komponent aktiveerib komponendi C5, millel on omadus kinnituda rakumembraanile. "hiliste" komponentide C6, C7, C8, C9 järjestikusel kinnitumisel C5 komponendile tekib lüütiline ehk membraani ründav kompleks, mis rikub membraani terviklikkust (moodustab sellesse augu) ja rakk sureb osmootse lüüsi tulemus.
Alternatiivne viis komplemendi aktiveerimine toimub ilma antikehade osaluseta. See rada on iseloomulik kaitsele gramnegatiivsete mikroobide eest. Alternatiivse raja kaskaadi ahelreaktsioon algab antigeeni interaktsioonist valkudega B, D ja propidiin (P) koos järgneva C3 komponendi aktiveerimisega. Edasi kulgeb reaktsioon samamoodi nagu klassikalisel teel – moodustub membraani ründav kompleks.
Lektiin panna Komplemendi aktiveerimine toimub ka ilma antikehade osaluseta. Selle käivitab spetsiaalne mannoosi siduv valkseerum, mis pärast interaktsiooni mikroobirakkude pinnal olevate mannoosijääkidega katalüüsib C4. Edasine reaktsioonide kaskaad on sarnane klassikalisele teele.
Komplemendi aktiveerimise protsessis moodustuvad selle komponentide proteolüüsi produktid - subühikud C3a ja C3b, C5a ja C5b ning teised, millel on kõrge bioloogiline aktiivsus. Näiteks C3a ja C5a osalevad anafülaktilistes reaktsioonides, on kemoatraktandid, C3b mängib rolli fagotsütoosiobjektide opsoniseerimisel jne. Ca ioonide osalusel toimub komplemendi kompleksne kaskaadreaktsioon. 2+ ja Mg2+.
Komplemendi bioloogilised funktsioonid
Odintsov Yu.N., Perelmuter V.M. Komplemendi bioloogilised funktsioonid
Odintsov Yu.N., Perelmuter V.M.
Siberi Riiklik Meditsiiniülikool, Tomsk
© Odintsov Yu.N., Perelmuter V.M.
Komplement on organismi vastupanuvõime üks olulisemaid tegureid. Komplemendi süsteem võib osaleda erinevates efektormehhanismides, peamiselt mikroorganismide lüüsis (komplementaarne tapmine) ja opsoniseerimises. Makrofaagid võivad olla seotud komplemendi lüütilise funktsiooni muutmisega opsooniliseks. Komplemendi funktsioonid bakterioosi korral sõltuvad nakkushaiguse patogeneesi omadustest.
Võtmesõnad: komplement, bakteriolüüs, opsonisatsioon, nakkusprotsess.
Üks tõelisi põhilisi takistustegureid on komplement. Selle peamised funktsioonid on bakteriaalne lüüs, bakteriaalne opsoneerimine fagotsütoosi jaoks. Opsoonilise funktsiooni lüütilise funktsiooni muutumine sõltub makrofaagidest. Komplemendi funktsioonid bakterioosi korral sõltuvad nakkushaiguse fatogeneesi tunnustest.
Märksõnad: komplement, bakteriolüüs, opsonisatsioon, nakkusprotsess.
UDK 576: 8 097,37
Inimkehal on kaks peamist kaitseliini nakkushaiguste patogeenide vastu: mittespetsiifiline (resistentsus) ja spetsiifiline (immuunsus).
Esimese kaitseliini (resistentsuse) tegureid iseloomustavad mitmed ühised tunnused: 1) need tekivad ammu enne kohtumist patogeeniga (emakasisene periood); 2) mittespetsiifiline; 3) on geneetiliselt määratud; 4) populatsioonis genotüübiliselt ja fenotüübiliselt heterogeenne (heterogeenne); 5) kõrget resistentsust ühele patogeenile saab kombineerida madala resistentsusega teise patogeeni suhtes; 6) resistentsus sõltub eelkõige makrofaagide funktsionaalsest seisundist, mida juhivad HLA-ga mitteseotud geenid, ja komplemendi süsteemi seisundist (kontrollib HLA).
Komplement - mitmekomponentne plasma ensüümsüsteem, mille koostis ja talitlus on üldiselt hästi uuritud, on organismi vastupanuvõime üks olulisemaid tegureid. 1960.-1970. aastatel. eriti populaarne oli komplemendi tiitri määratlemine ühe resistentsuse näitajana. Ja praegu on palju uuringuid, mis on pühendatud komplemendi funktsiooni uurimisele. Siiski on
mitte ainult teatud raskused ja vastuolud komplemendi aktivatsiooni mehhanismi selgitamisel, vaid siiski
mõningaid komplemendi aktiveerimise ja funktsioneerimise mehhanisme ei ole piisavalt uuritud. Need vastuolulised küsimused hõlmavad komplemendi aktiveerimise inhibiitorite toimemehhanismi in vivo, komplemendi aktiveerimise lüütilisest funktsioonist opsooniliseks muutmise mehhanismi ja komplemendi rolli mõistmist sanogeneesis mitmesuguste infektsioonide korral.
Vereplasmas on teada 14 valku (komponenti), mis moodustavad komplemendi süsteemi. Neid sünteesivad hepatotsüüdid, makrofaagid ja neutrofiilid. Enamik neist on p-globuliinid. WHO poolt vastu võetud nomenklatuuri kohaselt tähistatakse komplemendisüsteemi sümboliga C ja selle üksikuid komponente sümbolitega Cl, C2, C3, C4, C5, C6, C7, C8, C9 või suurtähtedega (D, B, P). Mõned komponendid (Cl, C2, C3, C4, C5, B) jagunevad neid moodustavateks alamkomponentideks - raskemateks, millel on ensümaatiline aktiivsus, ja vähem rasketeks, millel puudub ensümaatiline aktiivsus, kuid mis säilitavad oma sõltumatuse. bioloogiline funktsioon... Komplemendisüsteemi valkude aktiveeritud kompleksid on tähistatud kompleksi kohal kriipsuga (näiteks C4b2a3b - C5-konvertaas).
Lisaks komplemendi enda valkudele (C1-C9) on selle bioloogilise aktiivsuse rakendamisel
osalus ja muud regulatiivseid funktsioone täitvad valgud:
a) makroorganismide rakkude membraanide retseptorid komplemendi alamkomponentidele: CR1 (CD35), CR2 (CD21), CR3 (CD11b / CD18), CR4 (CD11c / CD18), C1qR, C3a / C4aR, C5aR;
b) makroorganismi rakkude membraanivalgud: membraani kofaktori valk (MKB ehk MCP – membraaniga assotsieeruv proteolüüsi kofaktor, CD46), dissotsiatsiooni kiirendav tegur (FUD ehk DAF – lagunemist kiirendav tegur, CD55), protektiin (CD59) );
c) positiivset või negatiivset regulatsiooni läbi viivad vereplasma valgud: 1) positiivne regulatsioon - faktor B, faktor D, propediin (P); 2) negatiivne regulatsioon - faktor I, faktor H, valku siduv C4b (C4 siduv proteiin, C4bp), C1-inhibiitor (C1-inh, serpiin), S-valk (vitro-nektiin).
Seega osaleb komplemendisüsteemi funktsioonides üle 30 komponendi. Igal komplemendi valgukomponendil (alakomponendil) on teatud omadused (tabel 1).
Tavaliselt on komplemendi komponendid plasmas passiivsed. Nad muutuvad aktiivseks mitmeastmeliste aktiveerimisreaktsioonide protsessis. Aktiveeritud komplemendi komponendid toimivad teatud järjekorras ensümaatiliste reaktsioonide kaskaadi kujul ja eelmise aktiveerimise saadus toimib katalüsaatorina uue alamkomponendi või komplemendi komponendi kaasamisel järgnevasse reaktsiooni.
Komplemendi süsteem võib osaleda erinevates efektormehhanismides:
1) mikroorganismide lüüs (komplementaarne tapmine);
2) mikroorganismide opsoniseerimine;
3) immuunkomplekside lõhustamine ja nende kliirens;
4) leukotsüütide aktiveerimine ja kemotaktiline külgetõmme põletikukoldesse;
5) spetsiifiliste antikehade indutseerimise tõhustamine: a) antigeeni lokaliseerimise suurendamine B-lümfotsüütide ja antigeeni esitlevate rakkude (APC) pinnal; b) B-lümfotsüütide aktivatsiooni läve alandamine.
Komplemendi funktsioonidest on kõige olulisemad patogeenide membraanilüüsimine ja mikroorganismide opsoniseerimine.
Tabel 1
Komplemendi komponendid ja alamkomponendid, mis on seotud komplemendi aktiveerimise klassikaliste ja alternatiivsete radadega
Komponent (alamkomponent) Molekulmass, kD Alamkomponent Kontsentratsioon vereseerumis, μg / ml Funktsioon
C1 1124 1 C1q 2 C1r 2 C1s – Ensüümikompleks
Clq 460–80 ^ või 1 dM antigeen-antikeha kompleksi pika ahelaga seondumine
Clr 166 - 30-50 Proteaasi aktiveeriv Cb
Cls 166 - 30-50 C4 ja C2 aktiveeriv seriinproteaas
C2 110 2a, 2b 15-25 moodustavad klassikalise tee C3-konvertaasi (C4b2a) ja seejärel C5-konvertaasi (C4b2a3b)
SZ 190 3a, 3b 1200
С4 200 4a, 4b 350-500
С5 191 5a, 5b 75 membraani ründava kompleksi moodustumine, mis moodustab sihtrakumembraanis poori
Tegur B95 Ba, Bb200
Tegur D 25 – 1
Properdiin (P) 220 25 Alternatiivse raja (C3Bb) C3-konvertaasi stabilisaator blokeerib C3Bb dissotsiatsiooni faktori H mõjul
Mikroorganismide komplementaarne lüüs
Mikroorganismide lüüs toimub membraanirünnakukompleksi (MAC) moodustumise tulemusena, mis koosneb
komplemendi komponentidest. Komplemendi aktiveerimiseks on mitu viisi, sõltuvalt sellest, kuidas MAC-i moodustumine toimus.
Klassikaline (immunokompleksne) komplemendi aktiveerimise rada
Seda komplemendi aktiveerimise rada nimetatakse klassikaliseks, kuna seda kirjeldati esmakordselt ja see jäi pikka aega ainsaks tänapäeval teadaolevaks. Klassikalises komplemendi aktiveerimise rajas mängib käivitavat rolli antigeen-antikeha kompleks (immuunkompleks (IC)). Komplemendi aktiveerimise esimene lüli on C1 komponendi C^-alakomponendi seondumine immuunkompleksi immunoglobuliiniga. Täpsemalt, komplemendi aktiveerimise korral klassi G immunoglobuliinide (1d31, IgG3, IgG3, Ig4) poolt teostavad seda DO raske ahela positsioonid 285, 288, 290, 292 aminohappejäägid. Selle saidi aktiveerimine toimub alles pärast antigeen-antikeha kompleksi (AG-AT) moodustumist. Komplemendi aktiveerimise võime klassikalisel rajal väheneb intensiivsusega 1 dM, ^ 3, DO1 ja DO2.
Komplemendi komponent C^ koosneb kolmest alaühikust (joonis 1), millest igaühel on kaks kohta AG-AT kompleksi 1e-ga seondumiseks. Seega on täielikul C^-molekulil kuus sellist tsentrit. AG-1dM kompleksi moodustumisel seondub C2 molekul sama 1dM molekuli vähemalt kahe teise domeeniga (CH2) ja osaledes G-klassi immunoglobuliinide AG-AT kompleksi moodustamisel, seondub see vähemalt kahe erineva molekuli ^ teine domeen (CH2) kompleksides AG-^. AG-AT-ga seotud C2 omandab seriinproteaasi omadused ja käivitab kahe C1r molekuli aktiveerimise ja inkorporeerimise C2-sse. C1r omakorda käivitab kahe teise molekuli C2 aktiveerimise ja liitumise C2-sse. Aktiveeritud C^-l on seriinesteraasi aktiivsus.
Seejärel lõikab C1 kompleksi C2 C4 suuremaks C4b fragmendiks ja väiksemaks C4a fragmendiks. C4b ühendub kovalentsete sidemetega rakumembraani molekulide amino- ja hüdroksüülrühmadega (joonis 2). Membraani pinnale fikseeritud C4b (või AG-AT kompleks) seob C2, mis muutub kättesaadavaks ensümaatiliseks lõhustamiseks sama seriinproteaasi C2 abil. Selle tulemusena moodustub väike fragment 2b ja suurem fragment C2a, mis koos membraani pinnale kinnitatud C4b-ga moodustavad ensümaatilise kompleksi C4b2a,
nimetatakse komplemendi aktiveerimise klassikalise raja SZ-konvertaasiks.
Riis. 1. Ensümaatilise kompleksi C1 (1d2r2e) komponendid ja selle interaktsioon antigeen-antikeha kompleksiga (AG- või AG-1dM): J - ahel, mis ühendab pentameeri monomeere
СЗЬВ - "- СЗЬР
ma -------------------
Võimendussilmus Joon. 2. Komplemendi aktiveerimine klassikalisel viisil
Saadud C3 konvertaas interakteerub C3-ga ja lõikab selle väiksemaks C3a fragmendiks ja suuremaks C3b fragmendiks. C3 kontsentratsioon plasmas on kõigist komplemendi komponentidest kõrgeim ja üks ensüümikompleks C4b2a (C3 konvertaas) on võimeline lõikama kuni 1000 C3 molekuli. See loob membraani pinnale kõrge C3b kontsentratsiooni (C3b moodustumise võimendamine). Seejärel seostub C3b kovalentselt C4b-ga, mis on osa C3 konvertaasist. Moodustunud kolmemolekulaarne kompleks C4b2a3b on C5-konvertaas. C3b C5-konvertaasi osana seondub kovalentselt mikroorganismide pinnaga (joonis 2).
C5-konvertaasi substraadiks on komplemendi C5 komponent, mille lõhustamine lõpeb väiksema C5a ja suurema C5b moodustumisega. Umbes-
C5b moodustumine käivitab membraani ründava kompleksi moodustumise. See toimub ilma ensüümide osaluseta, lisades järjestikku C5b-le komplemendi komponendid C6, C7, C8 ja C9. C5b6 on hüdrofiilne ja C5b67 on hüdrofoobne kompleks, mis on lülitatud membraani lipiidide kaksikkihti. C8 lisamine C5b67-le sukeldab moodustunud C5b678 kompleksi veelgi rohkem membraani. Ja lõpuks, 14 C9 molekuli on fikseeritud C5b678 kompleksiga. Moodustunud C5b6789 on membraani ründav kompleks. C9 molekulide polümerisatsioon C5b6789 kompleksis viib membraanis mittekokkuvarisevate pooride moodustumiseni. Läbi pooride vesi ja N8 + sisenevad rakku, mis viib raku lüüsini (joonis H).
Lahustunud ühendid
MAC moodustumise intensiivsus klassikalises komplemendi aktiveerimise rajas suureneb komplemendi aktiveerimise alternatiivse raja amplifikatsiooniahela tõttu. Amplifikatsioonisilmus algab hetkest, kui C3b kovalentne side moodustub membraani pinnaga. Silmuse moodustamisel osalevad kolm täiendavat plasmavalku: B, D ja P (properdiin). Faktori D (seriini esteraasi) mõjul lõhustatakse C3b-ga seotud proteiin B väiksemaks Ba fragmendiks ja suuremaks Bb-fragmendiks, mis seondub C3b-ga (vt joonis 2). Profaktiini lisamine CZLBL kompleksile, mis toimib C3b Bb kompleksi stabilisaatorina, viib lõpule alternatiivse raja CSTLBP C3-konvertaasi moodustumise. Alternatiivse raja C3-konvertaas lõhustab C3 molekule, moodustades täiendava C3b, mis tagab kõigi molekulide moodustumise. rohkem C5-konvertaas ja lõpuks rohkem MAC-i. MAK tegutseb
see sõltumatult ja võib-olla indutseerib kaspaaside raja kaudu apoptoosi.
Komplemendi aktiveerimise alternatiivne (spontaanne) rada
Komplemendi aktiveerimise mehhanism alternatiivse raja kaudu on tingitud tioeetri sideme spontaansest hüdrolüüsist natiivses C3 molekulis. See protsess toimub plasmas pidevalt ja seda nimetatakse SZ-i tühikäigu aktiveerimiseks. C3 hüdrolüüsi tulemusena moodustub selle aktiveeritud vorm, tähisega C31. Lisaks seob C3i faktorit B. Tegur D lõhustab C3iB kompleksi faktori B väikeseks fragmendiks Ba ja suureks Bb. Saadud C3iBb kompleks on komplemendi aktiveerimise alternatiivse raja vedelfaasi C3-konvertaas. Lisaks lõikab vedelfaasi C3iBb konvertaas C3 C3a ja C3b. Kui C3b jääb vabaks, hävitatakse see veega hüdrolüüsil. Kui C3b seostub kovalentselt bakterimembraani (mis tahes mikroorganismi membraani) pinnaga, siis proteolüüsi ei toimu. Veelgi enam, see algatab alternatiivse raja võimendusahela moodustumise. Fikseeritud C3b-le lisatakse faktor B (C3b on faktori B suhtes suurem afiinsusega kui faktori H suhtes), moodustub kompleks C3bB, millest faktor D
eraldab väikese osa Ba-st. Pärast CSTBb kompleksi stabilisaatoriks oleva properdiini kinnitumist moodustub CSTBBP kompleks, mis on membraanipinnaga seotud alternatiivse raja C3-konvertaas. Seotud C3 konvertaas algatab täiendavate C3b molekulide kinnitumise samas kohas (C3b amplifikatsioon), mis viib C3b kiire lokaalse akumuleerumiseni. Lisaks lõikab seotud C3 konvertaas C3 C3a ja C3b. C3b lisamine C3-konvertaasile moodustab C3b3b (C3b2Bb) kompleksi, mis on alternatiivse raja C5-konvertaas. Seejärel C5 komponent lõhustatakse ja MAA moodustub, nagu klassikalises komplemendi aktiveerimise rajas.
Spontaanne hüdrolüüs
I___________________________I
Võimendussilmus
Riis. 4. Komplemendi aktiveerimise alternatiivne (spontaanne) rada
"Tühikäigu" aktiveerimine
Mikroorganism
Lektiini rada komplemendi aktiveerimiseks
Gramnegatiivsete bakterite lipopolüsahhariidid (LPS), mis võivad sisaldada mannoosi, fukoosi, glükoosamiini jääke, seostuvad lektiinidega (vadakuvalgud, mis seovad tugevalt süsivesikuid) ja kutsuvad esile komplemendi aktivatsiooni lektiini raja. Näiteks võib komplemendi aktiveerimise lektiini raja käivitaja olla mannaani siduv lektiin (MSL), nagu C^, mis kuulub kaltsiumist sõltuvate lektiinide perekonda.
See ühineb mannoosiga, mis on osa bakteriraku seinast, ja omandab võime interakteeruda kahe mannaani siduva lektiiniga seotud seriinproteaasiga - MASP1 ja MASP2, mis on identsed vastavalt C1r ja C1s-ga.
Interaktsioon [MSL-MASP1-MASP2] on sarnane kompleksi [C^-C1r-C^] moodustumisega. Edaspidi toimub komplemendi aktiveerimine samamoodi nagu klassikalisel rajal (joonis 5).
4a 2b C3a C3b C5a
Võimendussilmus
Riis. 5. Komplemendi aktiveerimise lektiini rada (M - mannoos raku pinnastruktuurides, näiteks LPS)
Pentraksiinide perekonna valgud, millel on lektiinide omadused, nagu amüloidvalk, C-reaktiivne valk, on samuti võimelised aktiveerima komplementi lektiini raja kaudu, interakteerudes bakteriraku seinte vastavate substraatidega. Seega aktiveerib C-reaktiivne valk grampositiivsete bakterite rakuseinas forsforüülkoliini. Seejärel käivitab aktiveeritud forsforüülkoliin klassikalise komplemendi kokkupaneku raja.
C3b, mis moodustub C3-st, seondub mis tahes C3-konvertaasi mõjul sihtmembraaniga ja muutub täiendava C3b moodustumise kohaks. Seda etapi etappi nimetatakse "võimendusahelaks". Olenemata komplemendi aktiveerimise teest, kui seda ei blokeeri üks regulatiivsetest teguritest, lõpeb see membraani ründava kompleksi moodustumisega, mis moodustab bakterimembraanis mittekokkuvariseva poori, mis viib selle surmani.
Komplemendi aktiveerimise alternatiivsed ja lektiinirajad nakkushaiguse initsiatsiooni ajastuse järgi on varajased. Need võivad aktiveeruda juba esimestel tundidel pärast patogeeni sisenemist makroorganismi sisekeskkonda. Klassikaline komplemendi aktiveerimise rada on hiline: see hakkab "töötama" alles siis, kui ilmuvad antikehad (1 dM,
Komplemendi aktiveerimise reguleerivad valgud
Komplemendi aktiveerimise protsessi reguleerivad membraani (tabel 2) ja plasma (tabel 3) valgud.
Komplemendi aktiveerimise teid ja MAC moodustumist võivad blokeerida mitmed tegurid:
1) klassikaline, lektiin:
C1-inhibiitori toime, mis seob ja inaktiveerib C1r ja C2;
Klassikalise ja lektiiniradade C3-konvertaasi (C4b2a) moodustumise pärssimine tegurite I, H, C4-bp, FUD, ICD ja C ^ 1 mõjul;
Komplemendi komponentide interaktsiooni pärssimine makroorganismi rakkude pinnaga FUD (55), CR1 (CD35), ICD (46) toimel;
2) alternatiiv:
C3iBb ja C3Bb komplekside dissotsiatsioon faktori H toimel;
C3b lõhustamine faktori I poolt ühe kolmest kofaktorist osalusel: faktor H (plasma), CR1 või ICD (seotud makroorganismi rakkude pinnal);
Alternatiivse raja C3-konvertaasi moodustumise pärssimine makroorganismi rakkude pinnal FUD, CR1 või LAB toimel.
tabel 2
Membraani reguleerivad valgud
Rakuline (asub makroorganismi rakkude membraanidel)
Factor Expression rakkudel Funktsioon Tulemus
CR1^ 35) B-lümfotsüüdid; monotsüüdid (makrofaagid); granulotsüüdid; follikulaarsed dendriitrakud; NK-rakud Inhibeerib C2 seondumist C4b-ga; põhjustab ja kiirendab C4b2a dissotsiatsiooni C4b ja 2a; katabolismi kofaktor C4b faktori I mõjul; katabolismi kofaktor C3b faktori I mõjul; kiirendab C3Bb dissotsiatsiooni C3b vabanemisega Supresseerib komplemendi aktivatsiooni mis tahes rajal oma keha rakumembraanidel
ICD ^ 46) T-lümfotsüüdid; B-lümfotsüüdid; monotsüüdid (makrofaagid); granulotsüüdid; dendriitrakud; NK-rakud Supresseerib konvertaaside moodustumist: C4b2a ja C3bb; katabolismi kofaktor C4b faktori I mõjul; C3b katabolismi kofaktor I faktori mõjul Sama
FUD ^ 55) T-lümfotsüüdid; B-lümfotsüüdid; monotsüüdid (makrofaagid); granulotsüüdid; dendriitrakud; NK-rakud; trombotsüüdid Supresseerib C4b2a konvertaasi moodustumist klassikalisel rajal; pärsib alternatiivse raja CSTBL konvertaasi moodustumist; inhibeerib C2 seondumist C4b-ga; kiirendab C4b2a dissotsiatsiooni C4b ja 2a; kiirendab CZBL dissotsiatsiooni CZB vabanemisega
Protectiin (59) Kõik makrorakud – seondub 5b678-ga ja pärsib selle sukeldumist membraani Hoiab ära lüüsi
organism | ja C9 | oma rakud
Tabel 3
Plasma reguleerivad valgud
Faktor Funktsioon Molekulmass ja kontsentratsioon seerumis Somaatilistele rakkudele ja (või) patogeenidele avalduva toime realiseerimine
Tegur H (seondub kergesti makroorganismi rakkude pinna siaalhapetega) pärsib klassikalise tee C4b2a konvertaasi moodustumist; pärsib alternatiivse raja C3bBb konvertaasi moodustumist; põhjustab vedelfaasi konvertaasi C3iBb dissotsiatsiooni C3i-ks ja Bb-ks; katabolismi kofaktor C3i ja Bb; indutseerib C3bBb konvertaasi dissotsiatsiooni C3b-ks ja Bb-ks 150 Kda, 500 μg/ml Supresseerib komplemendi aktivatsiooni mis tahes raja kaudu oma organismi ja mikroorganismide rakumembraanidel
Faktor I (plasma proteaas) Inhibeerib C4b2a konvertaasi moodustumist klassikalisel rajal 90 KDa, 35 μg/ml Inhibeerib komplemendi aktivatsiooni klassikalisel rajal oma keha ja mikroorganismide rakumembraanidel
Koos ühe kofaktoriga (ICD, CR1, C4bp) lõikab see 4b C4c-ks ja C4d-ks; koos ühe kofaktoriga (ICD, CR1, H) lõikab C3b; katabolismi faktor C3b ja C3i pärsib komplemendi aktivatsiooni mis tahes rajal oma keha rakkude membraanidel
C4bp (C4 siduv valk, valku siduv C4b) Inhibeerib C2 seondumist C4b-ga; pärsib klassikalise raja C4b2a konvertaasi teket; põhjustab C4b2a dissotsiatsiooni C4b ja 2a; katabolismi kofaktor C4b faktori I mõjul 560 Kda, 250 μg / ml Supresseerib komplemendi aktivatsiooni klassikaliste ja lektiiniradade kaudu oma organismi ja mikroorganismide rakkude membraanidel
C1 inhibiitor (C1-inh, serpiin) Seob ja inhibeerib C1r ja C1 s (seriini proteaasi inhibiitor); lõikab C1r ja C1s C1q-st (C1q jääb seotuks Ig Fc fragmendiga); piirab C1 s kokkupuuteaega C4 ja C2-ga; piirab C1 spontaanset aktiveerimist vereplasmas 110 KDa, 180 μg / ml Supresseerib komplemendi aktivatsiooni klassikalise ja lektiini radade kaudu oma keha rakkude ja mikroorganismide membraanidel
S-valk (vitronektiin) Moodustab 5b67-S kompleksi, inaktiveerib selle võimet tungida membraani lipiidikihti 85 Kda, 500 μg / ml Blokeerib MAC moodustumise
MAC moodustumise pärssimine Vastupidi, plasmapäritolu reguleerivad valgud
Ootused pärsivad komplemendi aktivatsiooni mitte ainult somaatiliste rakkude pinnal, vaid ka patogeenide membraanidel.
Mikroorganismide opsoniseerimine komplemendi komponentidega
Mikroorganismide komplementaarne lüüs on mikroorganismi varajane reaktsioon patogeenide sisenemisele tema sisekeskkonda. Komplemendi aktiveerimisel alternatiivse ehk lektiiniraja kaudu moodustunud alamkomponendid C2b, C3a, C4a, C5a, Ba meelitavad rakke põletikukoldesse ja aktiveerivad nende efektorfunktsioonid.
Komplemendi komponentidest omavad opsoneerivad omadused peamiselt 3b ja 4b. Nende moodustamiseks on vaja kahte tingimust: esimene on komplemendi aktiveerimine ühe ülalkirjeldatud radade kaudu, teine on aktiveerimisprotsessi blokeerimine, mille tõttu on MAC-i moodustumine ja patogeeni lüüs võimatu. . See on
patogeenide pinnal.
1. Hüdrofoobset kompleksi C5b67, mis hakkab integreeruma membraani lipiidide kaksikkihti, saab inaktiveerida S-valk (vitronektiin). Moodustunud 5b67S kompleks ei suuda tungida membraani lipiidikihti.
2. Komponendi 8 kinnitamine kompleksi C5b67 külge vedel faas võib blokeerida madala tihedusega lipoproteiinide (LDL) poolt.
3. C5b678 membraani sukeldamine ja C9 kinnitumine hoiab ära CD59 (protektiini), makroorganismi rakkude membraani valgu.
4. Sisseehitatud MAC-ga makroorganismi rakkude membraani fragmentide eemaldamine endotsütoosi või eksotsütoosi teel.
Seega inhibeerivad rakulise päritoluga regulatoorsed valgud iseseisvalt komplemendi aktivatsiooni MAK-i moodustumisega ainult somaatiliste rakkude pinnal ega ole efektiivsed lüütilise inhibeerimisel.
Makroorganismi rakkudel on membraanil C3b ja selle C3b lagunemise membraani alamkomponendil vastavad retseptorid (tabel 4). C3b ja inaktiveeritud C3b (C3N) on ligandid retseptoritele CR1 (C3b, C3N), CR3 (C3N), CR4 (C3N), mis paiknevad neutrofiilidel, monotsüütidel (makrofaagidel), nabanööri endoteelil. СЗБ ja СЗЗ mängivad aktiivsete opsoniinide rolli.
Arvatavasti võib faktorite I ja H kombineeritud toime lülitada lüütilise kompleksi (MAC, komplementaarne tapmine) moodustumise teise patogeeni hävitamise mehhanismi – fagotsütaarse tapmise – vastu (joonis 6). Lahustuvad komplemendi aktivatsiooni inhibiitorid (I ja H), mida toodavad hiljem põletikukoldesse ilmuvad makrofaagid, toimivad fagotsüütide mikrokeskkonnas, takistades C3 konvertaasi moodustumist bakteri pinnal ja tagades seeläbi "vaba" C3b olemasolu. . Makrofaagi retseptor C3b-le, sidudes ligandi (C3b), fikseerib bakteri makrofaagi pinnal. Selle fagotsütoos viiakse läbi kahe ligand-retseptori kompleksi ühisel osalemisel: C3b + C3b ja FcyR + ^ retseptor. Teine paar, C3b + C3N retseptor, käivitab fagotsütoosi isegi ilma antikehadeta.
Komplemendi aktiveerimise lüütiliselt opsooniliseks funktsiooniks muutmise bioloogiline tähendus seisneb tõenäoliselt selles, et kõik bakterid, mida enne fagotsüütidega kohtumist ei lüüsitud, tuleks fagotsüteerida, kasutades C3b-opsoniini. Selline mehhanism komplemendi aktiveerimise ümberlülitamiseks opsooniliseks on vajalik mitte ainult elujõuliste patogeenide fagotsütoosiks infektsiooni varases staadiumis, vaid ka mikroorganismide "fragmentide" kasutamiseks fagotsüütides.
Tabel 4
Komplemendi alamkomponentide retseptorid
Komplemendi retseptori (CR) ligandid Ekspressioon rakkudel Seondumisefekt
CR1 (CD35) C3bi> C3b, C4b Neutrofiilid, monotsüüdid (makrofaagid), B-lümfotsüüdid, folliikulite dendriitrakud, erütrotsüüdid, neeru glomerulaarepiteel Opsoniseeritud fagotsütoos, B-lümfotsüütide aktivatsioon, immuunkomplekside transport erütrotsüütidele
CR3 (CD11b / CD18) C3bi Neutrofiilid, monotsüüdid (makrofaagid), NK-rakud, follikulaarsed dendriitrakud Opsoniseeritud fagotsütoos
CR4 (p 150-95) (CD11c / CD18) C3bi neutrofiilid Opsoniseeritud fagotsütoos
CR2 (CD21), B-lümfotsüütide (BCR + CD19, CR2, CD81) C3bi, C3dg B-rakkude, folliikulite dendriitrakkude korretseptorkompleksi komponent Parandab BCR aktivatsioonireaktsioone, indutseerib AG-AT kompleksi mitte-gootsüüdi sidumist. folliikulite dendriitrakkudel
lüütilise komplemendi aktiveerimisprogrammi lülitamine opsoonilisele.
Nakkusprotsessi tegelikes tingimustes võib regulatiivsete valkude mõju tõttu tekkida üleminek opsoonilise komplemendi aktiveerimisprogrammile, mis tagab patogeeni fagotsütoosi ja immuunkomplekside kliirensi. Komplemendi komponentide kokkupanek membraanil võib lõppeda membraani ründava kompleksi moodustumisega või katkeda 4b moodustumise tasemel ja veelgi aktiivsemalt 3b moodustumise tasemel faktorite I ja H poolt.
Tegur I on peamine ensüüm, mis põhjustab C3b lagunemist. Tegur H mängib selles protsessis kofaktori rolli. Koos toimides on neil võime inaktiveerida nii vedelfaasi kui ka membraani C3b (vaba või osana konvertaasist), eraldades sellest C3f fragmendi (inaktiveeritud C3b tähistatakse kui C3b). Seejärel jätkavad nad SZA lõikamist järgmiselt:
f ^ alamkomponendi alamkomponent
sz gg gg
Komplemendi edasise aktiveerimise blokaad
Bakter
Üleminek fagotsütoosi protsessile
Tegur H (kofaktor)
Makrofaagid
Bakterite omastamine
Y Pc-fragmendi X retseptor, 1 C3b komplemendi komponent
1 | 1 V retseptor СЗБ või СЗЗ komplemendi komponendi jaoks
Riis. 6. Komplemendi aktiveerimise lülitamine fagotsütoosi protsessile
Soovitatav on kaaluda küsimust komplemendi võimalikust rollist erinevate bakterioosirühmade patogeneesis, mis on varem jagatud sanogeneesi mehhanismi järgi.
Toksigeenne bakterioos (difteeria, gaasigangreen, botulism, teetanus jne). Patogeenide tavaline lokaliseerimine on nakkuse sissepääsu värav. Patogeneesi peamine efektor on toksiin (T-sõltuv antigeen, esimest tüüpi antigeen). Nende bakterite T-sõltuvad pinnaantigeenid mängivad immuunvastuse esilekutsumisel ebaolulist rolli. Sanogeneesi peamine efektor on antitoksiin Immuunvastuse tüüp on T1l2. Taastumine toimub tänu immuunkomplekside moodustumisele ja sellele järgnevale eliminatsioonile, samuti bakterite fagotsütaarsele hävitamisele põletikukoldes. Komplemendi roll nendes bakterioosides piirdub tõenäoliselt tema osalemisega immuunkomplekside toksiini - antitoksiini elimineerimisel. Komplement ei mängi olulist rolli toksiini neutraliseerimisel (st toksikogeensete infektsioonide sanogeneesis).
Mittetoksigeensed mittegranulomatoossed bakterioosid
1. Patogeenid sisaldavad pinna T-sõltumatuid antigeene (T "1 antigeenid, teist tüüpi antigeenid):
Bakterid sisaldavad klassikalist LPS-i (enteropatogeensete Escherichia coli, Salmonella, Shigella jt tiantigeenid). Tavaline patogeenide lokaliseerimine on sooletrakti limaskestade sissepääsuväravatest kuni piirkondlike lümfisõlmedeni. Patogeneesi peamine efektor on endotoksiin ja elusbakterid. Immuunvastuse tüüp on T1l2. Immuunsus
vastust LPS-ile iseloomustab 1dM klassi antikehade tootmine. Sanogenees toimub peamiselt bakterite hävitamise tõttu mitte-gotsüütilisel teel nakkusprotsessi immuuneelses faasis lektiini ja komplemendi aktiveerimise alternatiivsete radade tõttu. Nakkusliku protsessi immuunfaasis - immuunlüüsi tõttu 1 dM ja komplemendi osalusel mööda klassikalist aktivatsioonirada. Fagotsütoos ei ole selle rühma bakterioosiga sanogeneesi puhul hädavajalik. Komplemendi süsteemi aktiveerimine nende haiguste korral võib soodustada sanogeneesi;
Bakterid sisaldavad pinna (kapsel) 7!Antigeene (pneumokokid, hemofiilsed bakterid jne). Tavaline patogeenide lokaliseerimine - alates sissepääsu väravatest kuni hingamisteede limaskestadeni kuni piirkondlike lümfisõlmedeni, tungivad sageli vereringesse. Patogeneesi peamine efektor on elusad bakterid. Immuunvastuse tüüp on T1l2. Immuunvastuses pinnaantigeenidele tekivad 1 dM antikehad. Sanogenees toimub peamiselt bakterite hävitamise tõttu mitte-gotsüütilisel teel nakkusprotsessi immuuneelses faasis lektiini ja komplemendi aktiveerimise alternatiivsete radade tõttu. Nakkusliku protsessi immuunfaasis - immuunlüüsi tõttu 1 dM ja komplemendi osalusel mööda klassikalist aktivatsioonirada. Selle rühma bakterite verre tungimise korral mängib põrn peamist rolli makroorganismi puhastamisel patogeenidest - halvasti opsoniseeritud (või opsoneerimata) bakterite fagotsütoosi peamisest kohas - ja võimest.
DM "sihib" selle poolt sensibiliseeritud bakterid Kupfferi rakkude fagotsütoosi suhtes, millele järgneb bakterifragmentide, mis pole veel lõpuni lagunenud, ülekandmine sapikapillaaridesse. Sapisoolad lagundavad bakterite fragmente, mis erituvad soolestikku. Komplemendisüsteemi aktiveerimine selles haiguste rühmas võib samuti soodustada sanogeneesi.
2. Patogeenid sisaldavad pinna T-sõltuvaid antigeene (T-antigeenid, esimest tüüpi antigeenid).
Patogeenide lokaliseerimine (stafülokokid, streptokokid jne) - sissepääsu väravad (nahk, limaskestad), piirkondlikud lümfisõlmed, süsteemsed kahjustused (elundid). Patogeneesi peamisteks mõjutajateks on elusbakterid ja vähemal määral ka nende toksiinid. Immuunvastuses on selgelt jälgitav muutus I dM sünteesis DO-ks. Immuunvastuse tüüp nakkushaiguse piisava käiguga (immuunpuudulikkuse tunnusteta patsientidel) on T1r2. Sanogenees on tingitud immuunfagotsütoosist, immuunlüüsist ja antitoksiinidest. Nende infektsioonide korral toimub immuniteeteelses faasis sanogenees alternatiivse komplemendi aktiveerimise ja bakterite opsoniseerimise teel komplemendi aktivatsiooniproduktidega, millele järgneb nende fagotsütoos. Nakkusliku protsessi immuunfaasis seostatakse sanogeneesi komplementaarse tapmisega klassikalises komplemendi aktiveerimise rajas DM ja DO osalusel, samuti komplemendi ja DO aktivatsiooni produktide poolt opsoniseeritud bakterite fagotsütoosiga.
Granulomatoosne bakterioos
1. Ägeda mitteepiteelirakkude granulomatoosse bakterioosi tekitajad (listeria, tüüfus salmonella, paratüüfus A, B jne).
Patogeenid sisaldavad pinna T-sõltuvaid antigeene. Elusbakterid on patogeneesi mõjutajad. Fagotsütoos on mittetäielik. Immuunvastuse tüüp on T1r2 ja TM. DM-i ilmnemisega kaasneb granuloomide moodustumine. Üleminek DM-lt DO-le viib granuloomide vastupidise arenguni. Sanogenees viiakse läbi alternatiivse komplemendi aktiveerimise ja bakterite opsoniseerimise teel komplemendi aktiveerimisproduktidega, millele järgneb nende fagotsütoos. Nakkusliku protsessi immuunfaasis seostatakse sanogeneesi komplementaarse tapmisega klassikalises komplemendi aktiveerimise rajas DM ja DO osalusel, samuti komplemendi ja DO aktivatsiooni produktide poolt opsoniseeritud bakterite fagotsütoosiga.
2. Kroonilise epiteelirakkude granulomatoosse bakterioosi tekitajad (mycobacterium tuberculosis, pidalitõbi; brutsella jt).
Patogeenid sisaldavad pinna T-sõltuvaid antigeene. Elusbakterid on patogeneesi mõjutajad. Fagotsütoos on mittetäielik. Immuunvastuse tüübid on Th2 ja Th1. Ilmselt võib IgM-i välimus olla ka granuloomide moodustumise juhtiv tegur. Thl tsütokiinide toime ei ole piisav fagotsütoosi lõpuleviimiseks, mis põhjustab epiteelirakkude ilmumist granuloomi. Ükski komplemendi aktiveerimise variantidest sanogeneesis ei mängi olulist rolli.
Järeldus
Komplement (komplementsüsteem) on üks esimesi humoraalseid tegureid, millega patogeen makroorganismi sisekeskkonda sattudes kokku puutub. Komplemendi komponentide aktiveerimise mehhanismid võimaldavad seda kasutada nii patogeenide lüüsimiseks kui ka fagotsütoosi tugevdamiseks. Kõigi bakteriaalsete nakkushaiguste puhul ei saa prognostilise analüüsina kasutada komplemendi sisaldust ja taset veres.
Kirjandus
1. Odintsov Yu.N., Perelmuter V.M., Klimentjeva T.K. Tuftsin: roll mittegranulomatoosse ja granulomatoosse bakterioosi tekkes // Bul. õde ravim. 2002. T. 1. nr 3. S. 98-102.
2. Perelmuter VM, Odintsov Yu.N. Klassi M immunoglobuliinide (IgM) põhiülesanne on reguleerida bakterite ja nende antigeenide hematokudebarjääri läbilaskvust // Bul. õde ravim. 2005. T. 4. nr 3. S. 38-42.
3. Royt A. Immunoloogia alused. Per. inglise keelest Moskva: Mir, 1991.328 lk.
4. Royt A, Brostoff J, Mail D. Immunology. Per. inglise keelest Moskva: Mir, 2000.581 lk.
5. Khaitov R.M., Ignatieva G.A., Sidorovitš I.G. Immunoloogia. M .: Meditsiin, 2000.432 lk.
6. Yarilin AA immunoloogia alused. M .: Meditsiin, 1999. 607 lk.
7. Alban S., Classen B., Brunner G., Blaschek W. Echinacea purpurea arabinogalaktaani valgu ja hepariini komplemendi moduleeriva toime erinevus // Planta Med. 2002. V. 68 (12). Lk 1118-1124.
8. Ambrosio A.R., De Messias-Reason I.J. Leishmania (Viannia) braziliensis: mannoosi siduva lektiini interaktsioon pinnaglüko-konjugaatidega ja komplemendi aktiveerimine. Antikehadest sõltumatu kaitsemehhanism // Parasite Immunol. 2005. V. 27. Lk 333-340.
9. Andersson J., Larsson R, Richter R. et al. Komplemendi aktiveerimise (RCA) mudelregulaatori seondumine biomaterjali pinnaga: pinnaga seotud faktor H pärsib komplemendi aktivatsiooni // Biomaterjalid. 2001. V. 22. Lk 2435-2443.
10. Bohana-Kashtan O., Ziporen L, Donin N. et al. Komplemendi poolt ülekantud rakusignaalid // Mol. Immunol. 2004. V. 41. Lk 583-597.
11. Bohlson S.S., Strasser J.A., Bower J.J., Schorey J.S. Komplemendi roll Mycobacterium avium patogeneesis: peremeesorganismi infektsioonile reageerimise in vivo ja in vitro analüüsid komplemendi komponendi C3 puudumisel // Infect. Immunol. 2001. V. 69. Lk 7729-7735.
12. Brown J.S., Hussell T, Gilliland S.M. et al. Klassikaline rada on domineeriv komplemendi rada, mis on vajalik kaasasündinud immuunsuse loomiseks Streptococcus pneumoniae infektsiooni vastu hiirtel // Proc. Natl. Acad. Sci. USA. 2002. V. 99. Lk 16969-16974.
13. Caragine T.A., Okada N., Frey A.B., Tomlinson S. Varajase, kuid mitte hilise komplemendi lüütilise raja kasvajaga ekspresseeritud inhibiitor suurendab kasvaja kasvu inimese rinnavähi rotimudelis // Cancer Res. 2002. V. 62. Lk 1110-1115.
14. Celik I., Stover C, Botto M. et al. Komplemendi aktiveerimise klassikalise raja roll eksperimentaalselt indutseeritud polümikroobse peritoniidi korral // Infect. Immun. 2001. V. 69. Lk 7304-7309.
15. Donin N, Jurianz K., Ziporen L. et al. Inimese kartsinoomirakkude komplemendiresistentsus sõltub membraani reguleerivatest valkudest, proteiinkinaasidest ja siaalhappest // Clin. Exp. Immunol. 2003. V. 131. Lk 254-263.
16. Fernie-King B. A., Seilly D. J., Willers Ch. et al. Komplemendi streptokoki inhibiitor (SIC) pärsib membraanirünnaku kompleksi, takistades c567 omastamist rakumembraanidele // Immunoloogia. 2001. V. 103. Väljaanne 3. Lk 390-408.
17. Frumeaux-Bacchi V., Dragon-Durey M. A., Blouin J. jt. Komplemendisüsteemi uurimine kliinilises praktikas // Ann. Med. Interne (Pariis). 2003. V. 154. Lk 529-540.
18. Imai M., Ohta R., Okada N, Tomlinson S. Komplemendi regulaatori inhibeerimine in vivo suurendab antikehateraapiat rinnanäärme adenokartsinoomi mudelis // Int. J. Vähk. 2004. V. 110. Lk 875-881.
19. Jiang H, Wagner E, Zhang H, Frank M.M. Komplemendi 1 inhibiitor on alternatiivse komplemendi raja regulaator // J. Exp. Med.
2001. V. 194. nr 11. Lk 1609-1616.
20. Langeggen H, Berge K.E., Johnson E, Hetland G. Inimese nabaveeni endoteelirakud ekspresseerivad komplemendi retseptorit 1 (CD35) ja komplemendi retseptorit 4 (CD11c / CD18) in vitro // Põletik.
2002. V. 26. nr 3. Lk 103 - 110.
21. Laufer J., Katz Y, Passwell J.H. Komplemendi valkude ekstrahepaatiline süntees põletiku korral // Mol. Immunol. 2001. V. 38. Lk 221-229.
22. Leslie R.G.Q., Nielsen C.H. Komplemendi aktiveerimise klassikalised ja alternatiivsed teed mängivad inimese B-lümfotsüütide spontaanses C3-fragmendi ladestumisel ja membraanirünnakukompleksi (MAC) moodustumisel selget rolli // Immunoloogia. 2004. V. 111. Väljaanne 1. Lk 86-98.
23. Lukas T.J., Munoz H., Erickson B.W. C1-vahendatud immuunhemolüüsi inhibeerimine inimese immunoglobuliini G teisest konstantsest domeenist pärinevate monomeersete ja dimeersete peptiididega // J. Immunology. 1981. V. 127. nr 6. Lk 2555-2560.
24. Nauta A. J., Daha M. R., Tijsma O. jt. Komplemendi membraanirünnaku kompleks kutsub esile kaspaaside aktivatsiooni ja apoptoosi // Europ. J. of Immun. 2002. V. 32. Väljaanne 3. Lk 783-792.
25. Nielsen C.H., Marquait H.V., Prodinger W.M., Leslie R.G. Komplemendi alternatiivse raja CR2-vahendatud aktiveerimine põhjustab inimese B-lümfotsüütide membraanirünnaku komplekside moodustumist // Immunol. 2001. V. 104. Lk 418-422.
26. Nielsen C.H., Pedersen M.L., Marquart H.V. et al. 1. tüüpi (CR1, CD35) ja 2. tüüpi (CR2, CD21) komplemendi retseptorite roll C3 fragmentide ladestumise ja membraanirünnaku kompleksi moodustumise soodustamisel inimese normaalsetel perifeersetel B-rakkudel // Eur. J. Immunol. 2002. V. 32. Lk 1359-1367.
27. Ren B., McCrory M. A., Pass C. jt. Streptococcus pneumoniae pinnavalgu A virulentsusfunktsioon hõlmab komplemendi aktivatsiooni pärssimist ja komplemendi retseptori poolt vahendatud kaitse kahjustamist // J. Immunol. 2004. V. 173. Lk 7506-7512.
28. Roos A., Ramwadhdoebe T.H., Nauta A.J. et al. Komplemendi aktiveerimise varase faasi terapeutiline pärssimine // Immunobioloogia. 2002. V. 205. Lk 595-609.
29. Roos A., Bouwman L. H., Munoz J. jt. Komplemendi lektiini raja funktsionaalne iseloomustus inimese seerumis // Mol. Immunol. 2003. V. 39. Lk 655-668.
30. Laul H, He C., Knaak C. jt. Komplemendi retseptori 2-vahendatud komplemendi inhibiitorite sihtimine komplemendi aktiveerimise kohtadesse // J. Clin. Investeeri. 2003. V. 111. Lk 1875-1885.
31. Thiel S, Petersen S. V., Vorup-Jensen T. jt. C1q ja mannaani siduva lektiini (MBL) interaktsioon C1r, C1s, MBL-ga seotud seriinproteaasidega 1 ja 2 ning MBL-ga seotud valguga MAp19 // J. Immunol. 2000. V. 165. Lk 878-887.
32. Windbichler M., Echtenacher B., Hehlgans T. jt. Komplemendi aktiveerimise lektiini raja kaasamine antimikroobsesse immuunkaitsesse eksperimentaalse septilise peritoniidi ajal // Infektsioon ja immuunsus. 2004. V. 72. nr 9. Lk 5247-5252.
Pole regulatiivset mehhanismid toimides mitmel etapil, oleks komplemendisüsteem ebaefektiivne; selle komponentide piiramatu tarbimine võib põhjustada raskeid, potentsiaalselt surmavaid keharakkude ja kudede kahjustusi. Esimeses etapis blokeerib C1 inhibiitor Clr ja Cls ensümaatilise aktiivsuse ning järelikult C4 ja C2 lõhustamise. Aktiveeritud C2 püsib vaid lühikest aega ja selle suhteline ebastabiilsus piirab C42 ja C423 eluiga. Alternatiivse raja C3-aktiveerival ensüümil C3bBb on samuti lühike poolväärtusaeg, kuigi ensüümikompleksi poolt õigediini sidumine pikendab kompleksi eluiga.
V seerum on olemas anafülatoksiini inaktivaator – ensüüm, mis lõikab N-terminaalse arginiini C4a, C3a ja C5a küljest ning vähendab seeläbi järsult nende bioloogilist aktiivsust. Faktor I inaktiveerib C4b ja C3b, faktor H kiirendab C3b inaktiveerimist faktori I poolt ja sarnane tegur, C4-siduv valk (C4-cb), kiirendab C4b lõhustumist faktori I poolt. Rakumembraanide kolm põhivalku – PK1 , membraani kofaktori valk ja lagunemist kiirendav faktor (FUR) – hävitavad nendel membraanidel moodustunud C3- ja C5-konvertaasi kompleksid.
muud rakumembraani komponendid- seotud valgud (mille hulgas on CD59 enim uuritud) - võivad siduda C8 või C8 ja C9, mis takistab membraani ründava kompleksi (C5b6789) integreerumist. Mõned seerumivalgud (mille hulgas on enim uuritud valk S ja klasteriin) blokeerivad C5b67 kompleksi kinnitumist rakumembraanile, C8 või C9 seondumist nende poolt (st täisväärtusliku membraani ründava kompleksi moodustumist) või muul viisil takistada selle kompleksi teket ja liitumist.
Komplemendi kaitsev roll
Neutraliseerimine viirused antikehad võimendavad C1 ja C4 ning suureneb veelgi C3b fikseerimisel, mis moodustub mööda klassikalist või alternatiivset rada. Seega muutub komplement eriti oluliseks viirusnakkuse varases staadiumis, kui antikehade hulk on veel madal. Antikehad ja komplement piiravad vähemalt mõne viiruse nakkavust ja moodustavad tüüpilised komplemendi "augud", mis on nähtavad elektronmikroskoopiaga. Clq interaktsioon selle retseptoriga opsoniseerib sihtmärki, st hõlbustab selle fagotsütoosi.
C4a, C3a ja C5a fikseeritakse nuumrakkude poolt, mis hakkavad eritama histamiini ja teisi vahendajaid, mis põhjustab vasodilatatsiooni ja põletikule iseloomulikku turset ja hüpereemiat. C5a mõjul eritavad monotsüüdid TNF-i ja IL-1, mis suurendavad põletikulist vastust. C5a on neutrofiilide, monotsüütide ja eosinofiilide peamine kemotaktiline tegur, mis on võimeline fagotsüteerima mikroorganisme, mida opsoneerib C3b või selle lõhustumise saadus iC3b. Rakuga seotud C3b edasine inaktiveerimine, mis viib C3d ilmumiseni, jätab selle ilma opsoneerivast aktiivsusest, kuid säilib selle võime seonduda B-lümfotsüütidega. C3b fikseerimine sihtrakule hõlbustab selle lüüsi NK-rakkude või makrofaagide poolt.
Köitmine C3b lahustumatute immuunkompleksidega solubiliseerib need, kuna ilmselt hävitab C3b antigeeni-antikeha kompleksi võrestruktuuri. Samal ajal muutub see kompleks võimalikuks interakteeruda erütrotsüütide C3b retseptoriga (PK1), mis kannab kompleksi maksa või põrna, kus see imendub makrofaagidesse. See nähtus seletab osaliselt seerumihaiguse (immuunkomplekshaiguse) teket C1, C4, C2 või C3 puudulikkusega inimestel.