Plasmamembraani funktsioonid. Plasmamembraan Plasma struktuur ja funktsioon
Plasmamembraanil on eriline positsioon, kuna see piirab rakku väljastpoolt ja on otseselt seotud rakuvälise keskkonnaga. Selle paksus on umbes 10 nm ja see on kõige paksem rakumembraan. Peamised komponendid on valgud (üle 60%), lipiidid (umbes 40%) ja süsivesikud (umbes 1%). Nagu kõik teised rakumembraanid, sünteesitakse seda EPS-kanalites.
Plasmalemma funktsioonid.
Transport.
Plasmamembraan on poolläbilaskev, st. valikuliselt erinevad molekulid läbivad seda erineva kiirusega. Ainete transportimiseks läbi membraani on kaks võimalust: passiivne ja aktiivne transport.
Passiivne transport. Passiivne transport või difusioon ei vaja energiasisendit. Laenguta molekulid difundeeruvad mööda kontsentratsioonigradienti, laetud molekulide transport sõltub vesiniku prootonite kontsentratsioonigradiendist ja transmembraansest potentsiaalide erinevusest, mis liidetakse elektrokeemiliseks prootonigradiendiks. Reeglina kannab membraani sisemine tsütoplasmapind negatiivset laengut, mis hõlbustab positiivselt laetud ioonide tungimist rakku. Difusiooni on kahte tüüpi: lihtne ja kerge.
Lihtne difusioon on tüüpiline väikestele neutraalsetele molekulidele (H 2 O, CO 2, O 2), samuti hüdrofoobsetele madala molekulmassiga orgaanilistele ainetele. Need molekulid võivad läbida membraani poore või kanaleid ilma igasuguse interaktsioonita membraanivalkudega seni, kuni säilib kontsentratsioonigradient.
Kergendatud difusioon on omane hüdrofiilsetele molekulidele, mis transporditakse läbi membraani ka mööda kontsentratsioonigradienti, kuid põhimõtteliselt spetsiaalsete membraanikandjate valkude abil. vormiriietust.
Hõlbustatud difusioon on väga selektiivne, kuna kandevalgul on transporditava ainega komplementaarne sidumiskeskus ja ülekandega kaasnevad valgu konformatsioonilised muutused. Üks võimalikest hõlbustatud difusiooni mehhanismidest on järgmine: transportvalk (translokaas) seob ainet, seejärel läheneb membraani vastasküljele, vabastab selle aine, omandab esialgse konformatsiooni ja on taas valmis transpordifunktsiooni täitma. Vähe on teada, kuidas valk ise liigub. Teine võimalik ülekandemehhanism hõlmab mitme kandjavalgu osalemist. Sel juhul läheb algselt seotud ühend ise ühelt valgult teisele, seostudes järjestikku ühe või teise valguga, kuni see on membraani vastasküljel.
Aktiivne transport. Selline transport toimub siis, kui ülekanne toimub kontsentratsioonigradienti vastu. See nõuab raku energiakulu. Aktiivne transport on mõeldud ainete kogunemiseks rakus. Energiaallikaks on sageli ATP. Aktiivseks transpordiks on lisaks energiaallikale vajalik ka membraanivalkude osalus. Üks loomarakkude aktiivsetest transpordisüsteemidest vastutab Na ja K + ioonide transpordi eest läbi rakumembraani. Seda süsteemi nimetatakse Na + - K * -pump. See vastutab rakusisese keskkonna koostise säilitamise eest, kus K + ioonide kontsentratsioon on kõrgem kui Na * ioonide oma.
Mõlema iooni kontsentratsioonigradienti hoitakse, kandes K + raku sisemusse ja Na + väljapoole. Mõlemad transpordid toimuvad kontsentratsioonigradienti vastu. Selline ioonide jaotus määrab rakkude veesisalduse, närvirakkude ja lihasrakkude erutatavuse ning muud normaalsete rakkude omadused. Na + -K + -pump on valk - transport ATPaasi. Selle ensüümi molekul on oligomeer ja tungib läbi membraani. Pumba töötamise täistsükli jooksul kantakse rakust rakkudevahelisse ainesse 3 Na + iooni ja vastupidises suunas - 2 K + iooni, kusjuures kasutatakse ATP molekuli energiat. Kaltsiumioonide (Ca 2+ -ATPaas) ülekandmiseks on olemas transpordisüsteemid, prootonpumbad (H + -ATPaas) jne.
Aine aktiivset ülekannet läbi membraani, mis toimub teise aine kontsentratsioonigradiendi energia tõttu, nimetatakse sümptom... Sel juhul on transpordi-ATPaasil mõlema aine sidumissaidid. Antiport on aine liikumine vastu tema kontsentratsioonigradienti. Sel juhul liigub teine aine oma kontsentratsioonigradienti mööda vastupidises suunas. Sümptomid ja antiport (kotransport) võivad ilmneda aminohapete imendumisel soolestikust ja glükoosi reabsorptsioonil primaarsest uriinist, kasutades Na +, K + -ATPaasi poolt tekitatud Na + ioonide kontsentratsioonigradiendi energiat.
Veel kaks transpordiliiki on endotsütoos ja eksotsütoos.
Endotsütoos- suurte osakeste püüdmine raku poolt. Endotsütoosiks on mitu võimalust: pinotsütoos ja fagotsütoos. Tavaliselt all pinotsütoos mõista vedelate kolloidosakeste püüdmist raku poolt, all fagotsütoos- kehakeste püüdmine (tihedamad ja suuremad osakesed kuni teiste rakkudeni). Pino ja fagotsütoosi mehhanism on erinev.
Üldiselt nimetatakse tahkete osakeste või vedelate tilkade sisenemist rakku väljastpoolt heterofagiaks. See protsess on enim levinud algloomadel, kuid inimesel (nagu ka teistel imetajatel) on see väga oluline. Heterofagia mängib olulist rolli keha kaitsmisel (segmenteeritud neutrofiilid - granulotsüüdid; makrofagotsüüdid), luukoe (osteoklastide) remodelleerumisel, türoksiini moodustumisel kilpnäärme folliikulite poolt, valkude ja teiste makromolekulide reabsorptsioonil proksimaalses nefronis. ja muud protsessid.
Pinotsütoos.
Selleks, et välised molekulid saaksid rakku siseneda, peavad need esmalt olema seotud glükokalüksi retseptoritega (membraani pinnavalkudega seotud molekulide kogum) (joonis).
Sellise seondumise kohas leitakse plasmalemma all klatriini valgu molekulid. Plasmalemma koos väljastpoolt kinnitunud molekulide ja tsütoplasma alla jääva klatriiniga hakkab tungima. Eend muutub sügavamaks, selle servad lähenevad ja seejärel sulguvad. Selle tulemusena eraldatakse plasmalemmast mull, mis kannab kinnipüütud molekule. Klatriin selle pinnal näeb elektroonilistel mikrograafidel välja ebaühtlase piirina, seetõttu nimetatakse selliseid mullid ääristatud.
Klatriin takistab vesiikulite kleepumist rakusiseste membraanide külge. Seetõttu saab ääristatud vesiikulid rakus vabalt transportida just nendesse tsütoplasma osadesse, kus nende sisu tuleks kasutada. Nii viiakse tuumasse eelkõige steroidhormoonid. Tavaliselt eemaldavad ääristatud vesiikulid aga varsti pärast plasmalemmast lõhustumist. Klatriin kantakse üle plasmalemmasse ja võib taas osaleda endotsütoosireaktsioonides.
Rakupinnal tsütoplasmas on püsivamad vesiikulid - endosoomid. Piirnevad vesiikulid vabastavad klatriini ja ühinevad endosoomidega, suurendades endosoomide mahtu ja pinda. Seejärel lõhustatakse endosoomide liigne osa uue vesiikulina, milles pole rakku sattunud aineid, need jäävad endosoomi. Uus vesiikul suunatakse raku pinnale ja sulandub membraaniga. Selle tulemusena taastub ääristatud vesiikuli lõhenemisel tekkiv plasmalemma vähenemine, samas kui selle retseptorid pöörduvad samuti tagasi plasmalemma.
Endosoomid sukeldatakse tsütoplasmasse ja liidetakse lüsosoomi membraanidega. Sellise sekundaarse lüsosoomi sissetulevad ained läbivad mitmesuguseid biokeemilisi muundumisi. Protsessi lõppedes võib lüsosoomi membraan laguneda fragmentideks ning lüsosoomi lagunemissaadused ja sisu muutuvad rakusiseste metaboolsete reaktsioonide jaoks kättesaadavaks. Näiteks aminohapped seonduvad tRNA-ga ja viiakse ribosoomidesse, samal ajal kui glükoos võib siseneda Golgi kompleksi või agranulaarse EPS-i tuubulitesse.
Kuigi endosoomidel ei ole klatriini piiri, ei sulandu need kõik lüsosoomidega. Osa neist on suunatud ühelt rakupinnalt teisele (kui rakud moodustavad epiteelikihi). Seal ühineb endosoomi membraan plasmolemmaga ja sisu eritub. Selle tulemusena kanduvad ained raku kaudu ühest keskkonnast teise ilma muutusteta. Seda protsessi nimetatakse transtsütoos... Valgumolekule, eriti immunoglobuliine, saab transportida ka transtsütoosi teel.
Fagotsütoos.
Kui suure osakese pinnal on molekulaarsed rühmitused, mida rakuretseptorid suudavad ära tunda, seostub see. Mitte mingil juhul ei ole võõrosakestel endil selliseid rühmi. Kuid kehasse sattudes ümbritsevad neid immunoglobuliinide (opsoniinide) molekulid, mida leidub alati veres ja rakuvälises keskkonnas. Fagotsüütrakud tunnevad immunoglobuliinid alati ära.
Pärast seda, kui võõrosakest katvad opsoniinid seonduvad fagotsüütide retseptoritega, aktiveerub selle pinnakompleks. Aktiini mikrofilamendid hakkavad müosiiniga suhtlema ja rakupinna konfiguratsioon muutub. Fagotsüütide tsütoplasma väljakasvud on venitatud ümber osakese. Need katavad osakese pinna ja ühinevad selle kohal. Väljakasvude välimised lehed ühinevad, sulgedes rakupinna.
Sügavad väljakasvulehed moodustavad imendunud osakese ümber membraani – see moodustub fagosoom. Fagosoom ühineb lüsosoomidega, mille tulemuseks on nende kompleks - heterolüsosoom (heterosoom, või fagolüsosoom). Selles toimub osakese kinnipüütud komponentide lüüs. Mõned lüüsiproduktid eemaldatakse heterosoomist ja rakud kasutavad neid, samas kui mõned ei pruugi olla lüsosomaalsete ensüümide toimele alluvad. Need jäägid moodustavad jääkkehasid.
Potentsiaalselt kõigil rakkudel on fagotsütoosivõime, kuid kehas on sellele suunale spetsialiseerunud vaid vähesed. Need on neutrofiilsed leukotsüüdid ja makrofaagid.
Eksotsütoos.
See on ainete eemaldamine rakust. Esiteks eralduvad suurmolekulaarsed ühendid Golgi kompleksis transpordimullide kujul. Viimased suunatakse mikrotuubulite osalusel raku pinnale. Vesiikuli membraan on põimitud plasmalemmasse ja vesiikuli sisu on väljaspool rakku (joon.) Vesiikuli sulandumine plasmalemmaga võib toimuda ilma täiendavate signaalideta. Seda eksotsütoosi nimetatakse konstitutiivne. Nii eemaldatakse rakust suurem osa tema enda ainevahetuse saadustest. Mitmed rakud on aga loodud sünteesima spetsiaalseid ühendeid – saladusi, mida kasutatakse teistes kehaosades. Selleks, et saladusega transpordivesiikul ühineks plasmamembraaniga, on vaja väliseid signaale. Alles siis toimub sulandumine ja saladus avalikustatakse. Seda eksotsütoosi nimetatakse reguleeritud... Signaalmolekule, mis soodustavad sekretsiooni vabanemist, nimetatakse liberiinid (vabastavad tegurid), ja kõrvaldamise vältimine - statiinid.
Retseptori funktsioonid.
Neid pakuvad peamiselt glükoproteiinid, mis asuvad plasmalemma pinnal ja on võimelised seonduma oma ligandidega. Ligand vastab oma retseptorile nagu luku võti. Ligandi seondumine retseptoriga põhjustab muutuse polüpeptiidi konformatsioonis. Sellise transmembraanse valgu muutusega luuakse side rakuvälise ja intratsellulaarse keskkonna vahel.
Retseptorite tüübid.
Valkude ioonikanalitega seotud retseptorid. Nad suhtlevad signaalmolekuliga, mis ajutiselt avab või sulgeb ioonide läbimise kanali. (Näiteks neurotransmitteri atsetüülkoliini retseptor on valk, mis koosneb 5 alaühikust, mis moodustavad ioonikanali. Atsetüülkoliini puudumisel kanal suletakse ning peale kinnitumist see avaneb ja laseb läbi naatriumioonid).
Katalüütilised retseptorid. Need koosnevad ekstratsellulaarsest osast (retseptor ise) ja rakusisesest tsütoplasmaatilisest osast, mis toimib ensüümi proliinkinaasina (näiteks kasvuhormooni retseptorid).
G-valkudega seotud retseptorid. Need on transmembraansed valgud, mis koosnevad ligandiga interakteeruvast retseptorist ja G-valgust (guanosiintrifosfaadiga seotud regulaatorvalk), mis edastab signaali membraaniga seotud ensüümile (adenülaattsüklaas) või ioonkanalisse. Selle tulemusena aktiveeritakse tsükliline AMP või kaltsiumiioonid. (Nii toimib adenülaattsüklaasi süsteem. Näiteks maksarakkudes on hormooninsuliini retseptor. Retseptori supertsellulaarne osa seondub insuliiniga. See põhjustab rakusisese osa – ensüümi adenülaattsüklaasi – aktiveerumist. See sünteesib ATP-st tsüklilist AMP-d, mis reguleerib erinevate intratsellulaarsete protsesside kiirust, põhjustades nende või teiste metaboolsete ensüümide aktiveerimist või inhibeerimist.
Retseptorid, mis tajuvad füüsilisi tegureid. Näiteks fotoretseptori valk rodopsiin. Valguse neeldumisel muudab see oma konformatsiooni ja ergastab närviimpulsi.
See koosneb bilipiidkihist, mille lipiidid on rangelt orienteeritud – lipiidide hüdrofoobne osa (saba) on suunatud kihi sissepoole, hüdrofiilne osa (pea) aga väljapoole. Plasmamembraani ehituses osalevad lisaks lipiididele kolme tüüpi membraanivalgud: perifeersed, integraalsed ja poolintegraalsed.
Üks membraanide uurimisvaldkondi on praegu nii erinevate struktuursete ja regulatoorsete lipiidide kui ka membraane moodustavate üksikute integraalsete ja poolintegraalsete valkude omaduste üksikasjalik uurimine.
Integraalsed membraanivalgud
Peamist rolli membraani enda organiseerimisel mängivad integraalsed ja poolintegraalsed valgud, millel on globulaarne struktuur ja mis on lipiidifaasiga seotud hüdrofiilsete - hüdrofoobsete interaktsioonide kaudu. Integraalsete valkude gloobulid tungivad läbi kogu membraani paksuse ning nende hüdrofoobne osa asub kerakese keskel ja on sukeldatud lipiidfaasi hüdrofoobsesse tsooni.
Poolintegraalsed membraanivalgud
Poolintegraalsetes valkudes on hüdrofoobsed aminohapped kontsentreeritud gloobuli ühele poolusele ja vastavalt sellele on gloobulid membraani ainult pooleldi sukeldatud, ulatudes väljapoole ühest (välis- või sise-) membraanipinnast.
Membraanvalkude funktsioonid
Plasmamembraani integraalsetele ja poolintegraalsetele valkudele määrati varem kaks funktsiooni: üldine struktuurne ja spetsiifiline. Vastavalt sellele eristati nende hulgas struktuurseid ja funktsionaalseid valke. Kuid membraanide valgufraktsioonide eraldamise meetodite täiustamine ja üksikute valkude üksikasjalikum analüüs näitavad nüüd, et kõigi membraanide jaoks pole universaalseid struktuurseid valke, millel pole spetsiifilisi funktsioone. Seevastu spetsiifiliste funktsioonidega membraanivalgud on väga mitmekesised. Need on valgud, mis täidavad retseptori funktsioone, valgud, mis on erinevate ühendite aktiivsed ja passiivsed kandjad, ja lõpuks valgud, mis on osa paljudest ensüümsüsteemidest. Materjal saidilt
Membraanvalkude omadused
Kõigi nende integraalsete ja poolintegraalsete membraanivalkude ühine omadus, mis ei erine mitte ainult funktsionaalselt, vaid ka keemiliselt, on nende põhiline võime liikuda, "hõljuda" membraani tasapinnal vedelas lipiidifaasis. Nagu eespool märgitud, on selliste liikumiste olemasolu mõnede rakkude plasmamembraanides katseliselt tõestatud. Kuid see pole kaugeltki ainus membraanivalkudes tuvastatud liikumistüüp. Lisaks külgsuunalisele nihkele võivad üksikud integraalsed ja poolintegraalsed valgud pöörata membraani tasapinnas horisontaalses ja isegi vertikaalses suunas ning muuta ka molekuli lipiidifaasi sukeldumise astet.
Opsin. Kõiki neid valgugloobulite mitmekesiseid ja keerulisi liikumisi illustreerib eriti hästi fotoretseptorrakkude membraanidele spetsiifilise opsiinvalgu näide (joonis 3). Teatavasti on pimedas opsiin seotud karotenoidse võrkkestaga, mis sisaldab topelt-ciss-sidet; võrkkesta ja opsiini kompleks moodustab rodopsiini ehk visuaalselt lilla. Rodopsiini molekul on võimeline membraani horisontaaltasandil külgsuunas liikuma ja pöörlema (joonis 3, A). Valguse käes võrkkest läbib fotoisomerisatsiooni ja muundub transvormiks. Sel juhul muutub võrkkesta konformatsioon ja see eraldub opsiinist, mis omakorda muudab pöörlemistasa horisontaalselt vertikaalseks (joon. 3, B). Selliste transformatsioonide tagajärjeks on membraani ioonide läbilaskvuse muutus, mis põhjustab närviimpulsi ilmumist.
Huvitav on see, et valgusenergiast põhjustatud muutused opsiini gloobulite konformatsioonis ei saa mitte ainult genereerida närviimpulssi, nagu see toimub silma võrkkesta rakkudes, vaid on ka lihtsaim fotosünteesisüsteem, mida leidub spetsiaalsetes lillades bakterites.
Bioloogilised membraanid moodustavad raku struktuurilise korralduse aluse. Plasmamembraan (plasmalemma) on membraan, mis ümbritseb elusraku tsütoplasma. Membraanid koosnevad lipiididest ja valkudest. Lipiidid (peamiselt fosfolipiidid) moodustavad kahekihilise kihi, milles molekulide hüdrofoobsed "sabad" on suunatud membraani sisemuse poole ja hüdrofiilsed - selle pindade poole. Valgu molekulid võivad paikneda membraani välis- ja sisepinnal, nad võivad osaliselt uppuda lipiidikihti või tungida sellesse läbi ja läbi. Enamik sukeldatud membraanivalkudest on ensüümid. See on plasmamembraani struktuuri vedeliku-luu-mosaiikmudel. Valgu- ja lipiidimolekulid on liikuvad, mis tagab membraani dünaamilisuse. Membraanid sisaldavad ka süsivesikuid glükolipiidide ja glükoproteiinide (glükokalüks) kujul, mis paiknevad membraani välispinnal. Valkude ja süsivesikute kogum iga raku membraanipinnal on spetsiifiline ja on omamoodi rakutüübi indikaator.
Membraani funktsioonid:
- Eraldamine. See seisneb barjääri moodustamises raku sisemise sisu ja väliskeskkonna vahel.
- Ainete vahetuse tagamine tsütoplasma ja väliskeskkonna vahel. Vesi, ioonid, anorgaanilised ja orgaanilised molekulid sisenevad rakku (transpordifunktsioon). Rakus moodustunud saadused (sekretoorne funktsioon) erituvad väliskeskkonda.
- Transport. Transport läbi membraani võib toimuda erineval viisil. Passiivne transport toimub ilma energiatarbimiseta, lihtsa difusiooni, osmoosi või hõlbustatud difusiooni teel kandevalkude abil. Aktiivne transport toimub kandevalkude abil ja see nõuab energiat (näiteks naatrium-kaaliumpump). Materjal saidilt
Suured biopolümeeride molekulid sisenevad rakku endotsütoosi tulemusena. See jaguneb fagotsütoosiks ja pinotsütoosiks. Fagotsütoos on suurte osakeste hõivamine ja neeldumine raku poolt. Nähtust kirjeldas esmakordselt I.I. Mechnikov. Esiteks kinnituvad ained plasmamembraanile, spetsiifilistele retseptorvalkudele, seejärel membraan vajub, moodustades depressiooni.
Tekib seedevakuool. See seedib rakku sisenenud aineid. Inimestel ja loomadel on leukotsüüdid võimelised fagotsütoosiks. Valged verelibled neelavad baktereid ja muid tahkeid osakesi.
Pinotsütoos on vedelikupiiskade püüdmine ja neelamine selles lahustunud ainetega. Ained kinnituvad membraanivalkudele (retseptoritele) ja tilk lahust ümbritsetakse membraaniga, moodustades vaakumi. Pinotsütoos ja fagotsütoos tekivad ATP energia kulutamisel.
- Sekretär. Sekretsioon – rakus sünteesitud ainete vabanemine raku poolt väliskeskkonda. Hormoonid, polüsahhariidid, valgud, rasvatilgad suletakse membraaniga piiratud vesiikulitesse ja lähevad plasmalemma. Membraanid ühinevad ja vesiikulite sisu väljutatakse rakku ümbritsevasse keskkonda.
- Rakkude ühendus koes (volditud väljakasvu tõttu).
- Retseptor. Membraanid sisaldavad suurt hulka retseptoreid – spetsiaalseid valke, mille ülesanne on edastada signaale väljastpoolt raku sisemusse.
1. Barjäär- tagab reguleeritud, selektiivse, passiivse ja aktiivse ainevahetuse keskkonnaga.
Rakumembraanidel on selektiivne läbilaskvus: glükoos, aminohapped, rasvhapped, glütserool ja ioonid difundeeruvad aeglaselt läbi nende, membraanid ise reguleerivad seda protsessi aktiivselt – osad ained lastakse läbi, teised aga mitte.
2. Transport- läbi membraani transporditakse aineid rakku ja sealt välja. Transport läbi membraanide tagab: toitainete kohaletoimetamise, ainevahetuse lõppproduktide eemaldamise, erinevate ainete eritumise, ioongradientide loomise, sobiva pH ja ioonide kontsentratsiooni hoidmise rakus, mis on vajalikud rakuensüümide tööks.
Ainete rakku sisenemiseks või nende eemaldamiseks rakust väljapoole on neli peamist mehhanismi:
a) Passiivne (difusioon, osmoos) (ei vaja energiatarbimist)
Difusioon
Ühe aine molekulide või aatomite levimine teise aine molekulide või aatomite vahel, mis viib nende kontsentratsioonide spontaanse ühtlustumiseni kogu hõivatud ruumala ulatuses. Mõnes olukorras on ühel ainel juba võrdsustatud kontsentratsioon ja nad räägivad ühe aine difusioonist teises. Sel juhul toimub aine ülekanne kõrge kontsentratsiooniga alalt madala kontsentratsiooniga alale (mööda kontsentratsiooni gradiendi vektorit (joonis 2.4).
Riis. 2.4. Difusiooniprotsessi diagramm
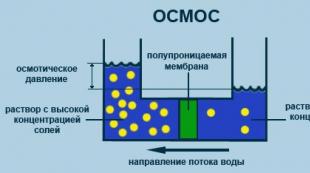
Osmoos
Ühesuunaline difusiooniprotsess läbi lahustimolekulide poolläbilaskva membraani lahustunud aine kõrgema kontsentratsiooni suunas väiksema lahustunud aine kontsentratsiooniga mahust (joonis 2.5).

Riis. 2.5. Osmoosi protsessi diagramm
b) Aktiivne transport (nõuab energiatarbimist)
Naatrium-kaaliumpump- naatriumioonide (rakust) ja kaaliumiioonide (rakku) aktiivse konjugeeritud transmembraanse transpordi mehhanism, mis tagab kontsentratsioonigradiendi ja transmembraanse potentsiaali erinevuse. Viimane on aluseks paljudele rakkude ja elundite funktsioonidele: näärmerakkude sekretsioon, lihaste kontraktsioon, närviimpulsside juhtimine jne. (joon. 2.6).

Riis. 2.6. Kaalium-naatriumpumba skeem
Esimeses etapis seob ensüüm Na + / K + -ATPaas kolm Na + iooni membraani siseküljelt. Need ioonid muudavad ATPaasi aktiivse keskuse konformatsiooni. Pärast seda on ensüüm võimeline hüdrolüüsima üht ATP molekuli. Pärast hüdrolüüsi vabanev energia kulub kandja konformatsiooni muutmisele, mille tõttu tekib membraani välisküljele kolm iooni Na + ja ioon PO 4 3– (fosfaat). Siin eralduvad Na + ioonid ja PO 4 3– asendatakse kahe K + iooniga. Pärast seda naaseb ensüüm oma esialgsesse konformatsiooni ja membraani siseküljele ilmuvad K + ioonid. Siin jagunevad K + ioonid ja kandja on taas tööks valmis.
Selle tulemusena tekib rakuvälises keskkonnas kõrge Na + ioonide kontsentratsioon ja raku sees K + kõrge kontsentratsioon. Seda kontsentratsiooni erinevust kasutatakse rakkudes närviimpulsi juhtimisel.
c) endotsütoos (fagotsütoos, pinotsütoos)
Fagotsütoos(söömine raku poolt) - tahkete esemete, näiteks eukarüootsete rakkude, bakterite, viiruste, surnud rakkude jäänuste jne imendumise protsess raku poolt. Imendunud objekti ümber moodustub suur rakusisene vakuool (fagosoom). Fagosoomide suurus on alates 250 nm ja rohkem. Fagosoomi liitmisel primaarse lüsosoomiga moodustub sekundaarne lüsosoom. Happelises keskkonnas lagundavad hüdrolüütilised ensüümid makromolekule sekundaarses lüsosoomis. Seejärel transporditakse lõhustumisproduktid (aminohapped, monosahhariidid ja muud kasulikud ained) läbi lüsosomaalse membraani raku tsütoplasmasse. Fagotsütoos on väga laialt levinud. Kõrgelt organiseeritud loomadel ja inimestel mängib fagotsütoosi protsess kaitsvat rolli. Leukotsüütide ja makrofaagide fagotsüütiline aktiivsus on väga oluline keha kaitsmisel patogeensete mikroobide ja muude soovimatute osakeste sattumise eest. Fagotsütoosi kirjeldas esmakordselt vene teadlane I.I.Mechnikov (joonis 2.7)
Pinotsütoos(raku joomine) - vedela faasi imendumise protsess lahustuvaid aineid, sealhulgas suuri molekule (valgud, polüsahhariidid jne) sisaldavast keskkonnast. Pinotsütoosiga eralduvad raku sees olevast membraanist väikesed vesiikulid – endosoomid. Need on väiksemad kui fagosoomid (suurus on kuni 150 nm) ega sisalda tavaliselt suuri osakesi. Pärast endosoomi moodustumist läheneb sellele primaarne lüsosoom ja need kaks membraani vesiikulit ühinevad. Saadud organelli nimetatakse sekundaarseks lüsosoomiks. Pinotsütoosi protsessi viivad pidevalt läbi kõik eukarüootsed rakud. (joon. 7)
Retseptor-vahendatud endotsütoos - aktiivne spetsiifiline protsess, mille käigus rakumembraan pundub rakku, moodustades servadega süvendid. Vooderdatud süvendi intratsellulaarne pool sisaldab adaptiivsete valkude komplekti. Makromolekulid, mis seostuvad spetsiifiliste retseptoritega rakupinnal, läbivad seest palju suurema kiirusega kui ained, mis sisenevad rakkudesse pinotsütoosi teel.

Riis. 2.7. Endotsütoos
d) eksotsütoos (negatiivne fagotsütoos ja pinotsütoos)
Rakuline protsess, mille käigus rakusisesed vesiikulid (membraanvesiikulid) sulanduvad raku välismembraaniga. Eksotsütoosi käigus vabaneb sekretoorsete vesiikulite (eksotsütaarsete vesiikulite) sisu väljapoole ja nende membraan ühineb rakumembraaniga. Selle meetodi abil vabanevad rakust peaaegu kõik makromolekulaarsed ühendid (valgud, peptiidhormoonid jne). (joonis 2.8)

Riis. 2.8. Eksotsütoosi skeem
3. Biopotentsiaalide teke ja läbiviimine- membraani abil hoitakse rakus konstantset ioonide kontsentratsiooni: K + iooni kontsentratsioon rakus on palju suurem kui väljaspool ja Na + kontsentratsioon palju madalam, mis on väga oluline, kuna see tagab potentsiaalide erinevuse säilimise membraanil ja närviimpulsi tekke.
4. Mehaaniline- tagab raku autonoomia, selle rakusisesed struktuurid, aga ka ühenduse teiste rakkudega (kudedes).
5. Energia- fotosünteesi käigus kloroplastides ja rakuhingamise käigus mitokondrites toimivad nende membraanides energiaülekandesüsteemid, milles osalevad ka valgud;
6. Retseptor- mõned valgud membraanis on retseptorid (molekulid, mille kaudu rakk tajub teatud signaale).
7. Ensümaatiline- membraanivalgud on sageli ensüümid. Näiteks sooleepiteelirakkude plasmamembraanid sisaldavad seedeensüüme.
8. Maatriks- tagab membraanivalkude teatud vastastikuse paigutuse ja orientatsiooni, nende optimaalse interaktsiooni;
9. Puuri märgistus- membraanil on antigeenid, mis toimivad markeritena - "sildid", mis võimaldavad teil rakku tuvastada. Need on glükoproteiinid (st valgud, mille külge on kinnitatud hargnenud oligosahhariidide kõrvalahelad), mis täidavad "antennide" rolli. Markerite abil suudavad rakud teisi rakke ära tunda ja nendega koos tegutseda, näiteks elundite ja kudede moodustumisel. Samuti võimaldab see immuunsüsteemil võõraid antigeene ära tunda.
Rakulised kandmised
Rakulised kandmised hõlmavad süsivesikuid, rasvu ja valke. Kõik need ained kogunevad raku tsütoplasmas erineva suuruse ja kujuga tilkade ja teradena. Neid sünteesitakse perioodiliselt rakus ja kasutatakse ainevahetusprotsessides.
Tsütoplasma
See on osa elusrakust (protoplastist), millel puudub plasmamembraan ja tuum. Tsütoplasma koostis sisaldab: tsütoplasma maatriksit, tsütoskeletti, organelle ja inklusioone (mõnikord ei klassifitseerita inklusioone ja vakuoolide sisu tsütoplasma elusaineks). Väliskeskkonnast plasmamembraaniga eraldatud tsütoplasma on rakkude sisemine poolvedel sööde. Tuum ja erinevad organellid asuvad eukarüootsete rakkude tsütoplasmas. See sisaldab ka mitmesuguseid lisandeid - rakulise aktiivsuse tooteid, vakuoole, aga ka väikseimaid torukesi ja filamente, mis moodustavad raku skeleti. Tsütoplasma põhiaine koostises on ülekaalus valgud.
Tsütoplasmaatilised funktsioonid
1) selles toimuvad peamised ainevahetusprotsessid.
2) ühendab tuuma ja kõik organellid üheks tervikuks, tagab nende vastasmõju.
3) liikuvus, ärrituvus, ainevahetus ja paljunemine.
Liikuvus avaldub erinevates vormides:
Raku tsütoplasma rakusisene liikumine.
Amööbikujuline liikumine. See liikumisvorm väljendub pseudopoodide moodustumisel tsütoplasma poolt selle või selle stiimuli suunas või sellest eemale. See liikumisvorm on omane amööbile, vere leukotsüütidele, aga ka mõnele koerakkudele.
Virvendav liikumine. See väljendub tillukeste protoplasmaatiliste väljakasvude – ripsmete ja viburite (ripsloomad, hulkraksete loomade epiteelirakud, spermatosoidid jne) peksmisena.
Lepinguline liikumine. See on ette nähtud spetsiaalsete organoidsete müofibrillide olemasolu tõttu tsütoplasmas, mille lühenemine või pikenemine aitab kaasa raku kokkutõmbumisele ja lõõgastumisele. Kokkutõmbumisvõime on kõige enam arenenud lihasrakkudes.
Ärrituvus väljendub rakkude võimes reageerida ärritusele ainevahetuse ja energia muutmise kaudu.
Tsütoskelett
Eukarüootse raku üheks eripäraks on skeleti moodustiste olemasolu selle tsütoplasmas mikrotuubulite ja valgukiudude kimpudena. Tsütoskeleti elemendid, mis on tihedalt seotud välimise tsütoplasmaatilise membraani ja tuumaümbrisega, moodustavad tsütoplasmas keerulisi kudesid.
Tsütoskeleti moodustavad mikrotuubulid, mikrokiud ja mikrotrabekulaarne süsteem. Tsütoskelett määrab raku kuju, osaleb raku liikumises, raku enda jagunemises ja liikumises, organellide rakusiseses transpordis.
Mikrotuubulid sisalduvad kõigis eukarüootsetes rakkudes ja on õõnsad hargnemata silindrid, mille läbimõõt ei ületa 30 nm ja seina paksus on 5 nm. Nende pikkus võib ulatuda mitme mikromeetrini. Kergesti lagunev ja uuesti kokkupandav. Mikrotuubulite sein koosneb peamiselt spiraalselt volditud tubuliini valgu subühikutest (joonis 2.09)
Mikrotuubulite funktsioonid:
1) täidab toetavat funktsiooni;
2) moodustab lõhustumisspindli; tagada kromosoomide lahknemine raku poolustele; vastutavad rakuorganellide liikumise eest;
3) osaleda rakusiseses transpordis, sekretsioonis, rakuseina moodustamises;
4) on ripsmete, lippide, basaalkehade ja tsentrioolide struktuurikomponent.
Mikrokiud on esindatud 6 nm läbimõõduga filamentidega, mis koosnevad aktiinivalgust, mis on lähedane lihase aktiinile. Aktiin moodustab 10-15% raku koguvalgust. Enamikus loomarakkudes moodustub plasmamembraani enda alla tihe aktiinifilamentide ja nendega seotud valkude võrgustik.
Lisaks aktiinile leidub rakus ka müosiini filamente. Nende arv on aga palju väiksem. Aktiini ja müosiini koostoime tõttu toimub lihaste kontraktsioon. Mikrokiud on seotud kogu raku või selle üksikute struktuuride liikumisega selles. Mõnel juhul pakuvad liikumist ainult aktiini filamendid, teistel - aktiin koos müosiiniga.
Mikrofilamentide funktsioonid
1) mehaaniline tugevus
2) võimaldab rakul muuta oma kuju ja liikuda.

Riis. 2.09. Tsütoskelett
Organellid (või organellid)
Jaotatakse mittemembraanne, ühemembraaniline ja kahemembraaniline.
TO mittemembraansed organellid eukarüootsed rakud hõlmavad organelle, millel pole oma suletud membraani, nimelt: ribosoomid ja tubuliini mikrotuubulite baasil ehitatud organellid - rakukeskus (tsentrioolid) ja liikumisorganellid (lipud ja ripsmed). Enamiku ainuraksete organismide ja valdava enamuse kõrgemate (maapealsete) taimede rakkudes tsentrioolid puuduvad.
TO ühemembraanilised organellid seotud: endoplasmaatiline retikulum, Golgi aparaat, lüsosoomid, peroksisoomid, sferosoomid, vakuoolid ja mõned teised. Kõik ühemembraanilised organellid on omavahel ühendatud üheks rakusüsteemiks. Taimerakkudel on spetsiaalsed lüsosoomid, loomarakkudel spetsiaalsed vakuoolid: seede-, eritus-, kontraktiilsed, fagotsüütilised, autofagotsüütilised jne.
TO kahe membraaniga organellid seotud mitokondrid ja plastiidid.
Mittemembraansed organellid
A) Ribosoomid- organellid, mida leidub kõigi organismide rakkudes. Need on väikesed organellid, mida esindavad umbes 20 nm läbimõõduga kerakujulised osakesed. Ribosoomid koosnevad kahest ebavõrdse suurusega subühikust – suurest ja väikesest. Ribosoomide hulka kuuluvad valgud ja ribosomaalne RNA (rRNA). Ribosoome on kahte peamist tüüpi: eukarüootsed (80S) ja prokarüootsed (70S).
Olenevalt lokalisatsioonist rakus paiknevad tsütoplasmas vabad ribosoomid, mis sünteesivad valke ja kinnitunud ribosoomid – ribosoomid, mis on ühendatud suurte alaühikutega EPR membraanide välispinnaga, sünteesivad valke, mis sisenevad Golgi kompleksi ja seejärel sekreteeritakse. rakk. Valkude biosünteesi käigus võivad ribosoomid ühineda kompleksideks – polüribosoomideks (polüsoomideks).
Eukarüootsed ribosoomid moodustuvad tuumas. Esiteks sünteesitakse nukleolaarsel DNA-l rRNA, mis seejärel kaetakse tsütoplasmast tulevate ribosomaalsete valkudega, mis lõhustatakse vajaliku suurusega ja moodustavad ribosoomi subühikuid. Tuumas ei ole täielikult moodustunud ribosoome. Subühikute ühinemine terveks ribosoomiks toimub tsütoplasmas, tavaliselt valkude biosünteesi käigus.
Ribosoome leidub kõigi organismide rakkudes. Igaüks neist koosneb kahest osakesest, väikesest ja suurest. Ribosoomid hõlmavad valke ja RNA-d.
Funktsioonid
valkude süntees.
Sünteesitud valgud kogunevad esmalt endoplasmaatilise retikulumi kanalitesse ja õõnsustesse ning seejärel transporditakse organellidesse ja rakuosadesse. EPS ja selle membraanidel asuvad ribosoomid kujutavad endast ühtset valkude biosünteesi ja transpordi aparaati (joon. 2.10-2.11).

Riis. 2.10. Ribosoomi struktuur

Riis. 2.11. Ribosoomi struktuur
B) Raku keskus (tsentrioolid)
Tsentriool on silinder (0,3 µm pikk ja 0,1 µm läbimõõduga), mille seina moodustavad üheksa kolme ühinenud mikrotuubuli rühma (9 kolmikut), mis on teatud ajavahemike järel ristsidumise teel omavahel ühendatud. Tsentrioolid on sageli paaris, kus nad on üksteise suhtes täisnurga all. Kui tsentriool asub tsiliumi või flagellumi põhjas, nimetatakse seda basaalkehaks.
Peaaegu kõigil loomarakkudel on paar tsentriooli, mis on rakukeskuse keskmine element.
Enne jagunemist lahknevad tsentrioolid vastaspoolustele ja nende lähedale ilmub tütartsentriool. Raku erinevatel poolustel paiknevatest tsentrioolidest moodustuvad üksteise poole kasvavad mikrotuubulid.
Funktsioonid
1) moodustavad mitootilise spindli, mis aitab kaasa geneetilise materjali ühtlasele jaotumisele tütarrakkude vahel,
2) on tsütoskeleti organisatsiooni keskus. Osa spindli filamente on kromosoomide külge kinnitatud.
Tsentrioolid on tsütoplasma isepaljunevad organellid. Need tekivad olemasolevate dubleerimise tulemusena. See juhtub siis, kui tsentrioolid lahknevad. Ebaküps tsentriool sisaldab 9 üksikut mikrotuubulit; ilmselt on iga mikrotuubul maatriks küpsele tsentrioolile iseloomulike kolmikute kokkupanekuks (joon. 2.12).
Tsetrioole leidub madalamate taimede (vetikate) rakkudes.

Riis. 2.12. Rakukeskuse tsentrioolid
Ühe membraaniga organellid
D) Endoplasmaatiline retikulum (EPS)
Kogu tsütoplasma sisemine tsoon on täidetud arvukate väikeste kanalite ja õõnsustega, mille seinad on plasmamembraaniga sarnased membraanid. Need kanalid hargnevad, ühenduvad üksteisega ja moodustavad võrgu, mida nimetatakse endoplasmaatiliseks retikulumiks. Endoplasmaatiline retikulum on struktuurilt heterogeenne. Tuntakse kahte tüüpi seda - granuleeritud ja sile.
Granuleeritud võrgu kanalite ja õõnsuste membraanidel on palju väikeseid ümaraid kehasid - ribosoom mis annavad membraanidele kareda välimuse. Sileda endoplasmaatilise retikulumi membraanid ei kanna oma pinnal ribosoome. EPS täidab palju erinevaid funktsioone.
Funktsioonid
Granuleeritud endoplasmaatilise retikulumi põhiülesanne on osalemine ribosoomides toimuvas valgusünteesis. Sileda endoplasmaatilise retikulumi membraanidel sünteesitakse lipiide ja süsivesikuid. Kõik need sünteesiproduktid kogunevad kanalitesse ja õõnsustesse ning transporditakse seejärel raku erinevatesse organellidesse, kus need tarbitakse või kogunevad tsütoplasmasse rakuliste inklusioonidena. EPS ühendab raku peamised organellid (joon. 2.13).

Riis. 2.13. Endoplasmaatilise retikulumi (EPS) ehk retikulumi struktuur
E) Golgi aparaat
Selle organoidi struktuur on taime- ja loomaorganismide rakkudes sarnane, hoolimata selle kuju mitmekesisusest. Sellel on palju olulisi funktsioone.
Ühe membraaniga organoid. See on lamestatud pikendatud servadega "tsisternide" virn, millega on seotud väikeste ühemembraaniliste mullide süsteem (Golgi mullid). Golgi mullid on koondunud peamiselt EPS-i külgnevale küljele ja virnade perifeeriasse. Arvatakse, et nad viivad valgud ja lipiidid üle Golgi aparaati, mille molekulid, liikudes tsisternist tsisternisse, läbivad keemilise modifitseerimise.
Kõik need ained esmalt akumuleeruvad, keemiliselt komplitseerivad ning seejärel sisenevad suurte ja väikeste mullidena tsütoplasmasse ning kas neid kasutatakse rakus endas selle eluea jooksul või eemaldatakse sealt ja kasutatakse organismis. (joon. 2.14-2.15).

Riis. 2.14. Golgi aparaadi struktuur
Funktsioonid:
Valkude, lipiidide, süsivesikute muutmine ja kogunemine;
Sissetulevate orgaaniliste ainete pakkimine membraani vesiikulitesse (vesiikulitesse);
Lüsosoomide moodustumise koht;
Sekretoorne funktsioon on seetõttu Golgi aparaat sekretoorsetes rakkudes hästi arenenud.

Riis. 2.15. Golgi kompleks
E) Lüsosoomid
Need on väikesed ümarad kehad. Lüsosoomi sees on ensüümid, mis lagundavad valke, rasvu, süsivesikuid, nukleiinhappeid. Lüsosoomid lähenevad tsütoplasmasse sattunud toiduosakesele, ühinevad sellega ning moodustub üks seedevakuool, mille sees on lüsosomaalsete ensüümidega ümbritsetud toiduosake.
Lüsosomaalsed ensüümid sünteesitakse töötlemata EPS-il, kantakse üle Golgi aparaati, kus neid modifitseeritakse ja pakitakse lüsosoomide membraani vesiikulitesse. Lüsosoom võib sisaldada 20 kuni 60 erinevat tüüpi hüdrolüütilist ensüümi. Ainete lagunemist ensüümide abil nimetatakse lüüsimine.
Eristage primaarseid ja sekundaarseid lüsosoome. Primaarseteks lüsosoomideks nimetatakse neid, mis on tekkinud Golgi aparaadist.
Sekundaarseid lüsosoome nimetatakse, mis moodustuvad primaarsete lüsosoomide ühinemise tulemusena endotsüütiliste vakuoolidega. Sel juhul seedivad nad fagotsütoosi või pinotsütoosi teel rakku sattunud aineid, mistõttu võib neid nimetada seedevakuoolideks.
Lüsosoomide funktsioonid:
1) endotsütoosi käigus rakku püütud ainete või osakeste (bakterid, muud rakud) seedimine;
2) autofaagia - raku jaoks mittevajalike struktuuride hävitamine, näiteks vanade organellide asendamisel uutega, või raku enda sees toodetud valkude ja muude ainete seedimine;
3) autolüüs - raku ise seedimine, mis viib selle surmani (mõnikord pole see protsess patoloogiline, vaid kaasneb keha arenguga või mõne spetsialiseeritud rakkude diferentseerumisega) (joon. 2.16-2.17).
Näide: Kui kulles muutub konnaks, seedivad sabarakkudes olevad lüsosoomid seda: saba kaob ning selle protsessi käigus tekkinud ained imenduvad ja kasutavad ära teised keharakud.

Riis. 2.16. Lüsosoomide moodustumine

Riis. 2.17. Lüsosoomide funktsioneerimine
G) Peroksisoomid
Organoidid, mis on struktuurilt sarnased lüsosoomidele, vesiikulid läbimõõduga kuni 1,5 mikronit homogeense maatriksiga, mis sisaldab umbes 50 ensüümi.
Katalaas põhjustab vesinikperoksiidi 2H 2 O 2 → 2H 2 O + O 2 lagunemist ja takistab lipiidide peroksüdatsiooni
Peroksisoomid tekivad pungudes varem olemasolevatest, s.o. kuuluvad isepaljunevate organellide hulka, hoolimata asjaolust, et nad ei sisalda DNA-d. Nad kasvavad ensüümide sisenemise tõttu neisse, peroksisoomi ensüümid moodustuvad karedal EPS-il ja hüaloplasmas (joonis 2.18).

Riis. 2.18. Peroksisoom (kristallilise nukleoidi keskel)
H) Vakuoolid
Ühe membraaniga organellid. Vakuoolid on "mahutid", mis on täidetud orgaaniliste ja anorgaaniliste ainete vesilahustega. EPS ja Golgi aparaat osalevad vakuoolide moodustumisel.
Noored taimerakud sisaldavad palju väikeseid vakuoole, mis seejärel rakkude kasvades ja diferentseerumisel üksteisega ühinevad ja moodustavad ühe suure tsentraalse vakuooli.
Tsentraalne vakuool võib hõivata kuni 95% küpse raku mahust, samal ajal kui tuum ja organellid surutakse tagasi rakumembraani. Taime vakuooli piiravat membraani nimetatakse tonoplast.
Vedelikku, mis täidab taime vakuooli, nimetatakse rakumahlaks. Rakumahla koostis sisaldab vees lahustuvaid orgaanilisi ja anorgaanilisi sooli, monosahhariide, disahhariide, aminohappeid, lõpp- või mürgiseid ainevahetusprodukte (glükosiidid, alkaloidid), mõningaid pigmente (antotsüaniine).
Orgaanilistest ainetest säilitatakse sagedamini suhkruid ja valke. Suhkrud - sagedamini lahuste kujul, valgud tulevad EPR-mullide ja Golgi aparaadi kujul, mille järel vakuoolid dehüdreeritakse, muutudes aleurooni teradeks.
Loomarakkudes on väikesed seede- ja autofaagilised vakuoolid, mis kuuluvad sekundaarsete lüsosoomide rühma ja sisaldavad hüdrolüütilisi ensüüme. Üherakulistel loomadel on ka kontraktiilsed vakuoolid, mis täidavad osmoregulatsiooni ja eritumise funktsiooni.
Funktsioonid
Taimedes
1) vedeliku kogunemine ja turgori säilitamine,
2) varutoitainete ja mineraalsoolade kogunemine,
3) lillede ja viljade värvimine ning seeläbi tolmeldajate ning viljade ja seemnete levitajate meelitamine.
Loomadel:
4) seedevakuoolid – hävitavad orgaanilisi makromolekule;
5) kontraktiilsed vakuoolid reguleerivad raku osmootset rõhku ja viivad rakust välja mittevajalikud ained
6) antigeenide immuunrakkude poolt fagotsütoosi käigus moodustuvad fagotsütaarsed vakuoolid
7) autofagotsütaarsed vakuoolid moodustuvad fagotsütoosi käigus nende enda kudede immuunrakkude poolt
Kahe membraaniga organellid (mitokondrid ja plastiidid)
Need organellid on poolautonoomsed, kuna neil on oma DNA ja oma valke sünteesiv aparaat. Mitokondreid leidub peaaegu kõigis eukarüootsetes rakkudes. Plastiide leidub ainult taimerakkudes.
I) Mitokondrid
Need on raku ainevahetusprotsesside energiavarustuse organellid. Hüaloplasmas on mitokondrid tavaliselt hajusalt jaotunud, kuid spetsialiseeritud rakkudes on nad koondunud nendesse piirkondadesse, kus on kõige suurem energiavajadus. Näiteks lihasrakkudes on suur hulk mitokondreid koondunud piki kontraktiilseid fibrillid, piki sperma flagellumit, neerutuubulite epiteeli, sünapside piirkonda jne. Selline mitokondrite paigutus tagab vähem ATP kadu. selle difusiooni ajal.
Välismembraan eraldab mitokondrid tsütoplasmast, on endaga suletud ega moodusta invaginatsioone. Sisemembraan piirab mitokondrite sisemist sisu – maatriksit. Iseloomulikuks tunnuseks on arvukate invaginatsioonide - cristae moodustumine, mille tõttu sisemembraanide pindala suureneb. Cristae arv ja arenguaste sõltub koe funktsionaalsest aktiivsusest. Mitokondritel on oma geneetiline materjal (joon. 2.19).
Mitokondriaalne DNA on suletud ringikujuline kaheahelaline molekul, inimese rakkudes on selle suurus 16569 nukleotiidipaari, mis on ligikaudu 105 korda väiksem kui tuumas paiknev DNA. Mitokondritel on oma valke sünteesiv süsteem, samas kui mitokondriaalsest mRNA-st transleeritud valkude arv on piiratud. Mitokondriaalne DNA ei saa kodeerida kõiki mitokondriaalseid valke. Enamik mitokondrites leiduvaid valke on tuuma geneetilise kontrolli all.

Riis. 2.19. Mitokondriaalne struktuur
Mitokondrite funktsioonid
1) ATP moodustumine
2) valgusüntees
3) osalemine spetsiifilistes sünteesides, näiteks steroidhormoonide (neerupealiste) sünteesis
4) kulunud mitokondrid võivad koguneda ka eritusprodukte, kahjulikke aineid, s.o. suudab täita teiste rakuorganellide funktsioone
K) Plastiidid
Plastiidid–Organellid, iseloomulikud ainult taimedele.
Plastiide on kolme tüüpi:
1) kloroplastid(rohelised plastiidid);
2) kromoplastid(kollased, oranžid või punased plastiidid)
3) leukoplastid(värvitud plastiidid).
Tavaliselt leidub rakus ainult ühte tüüpi plastiide.
Kloroplastid
Neid organelle leidub lehtede ja teiste roheliste taimeorganite rakkudes, aga ka mitmesugustes vetikates. Kõrgemad taimed sisaldavad tavaliselt ühes rakus mitukümmend kloroplasti. Kloroplastide roheline värvus sõltub klorofüllipigmendi sisaldusest neis.
Kloroplast on taimerakkude peamine organoid, milles toimub fotosüntees ehk orgaaniliste ainete (süsivesikute) moodustumine anorgaanilistest (CO 2 ja H 2 O) päikesevalguse energia abil. Kloroplastid on struktuurilt sarnased mitokondritega.
Kloroplastidel on keeruline struktuur. Need on hüaloplasmast piiritletud kahe membraaniga – välise ja sisemise membraaniga. Sisemist sisu nimetatakse strooma... Sisemembraan moodustab keeruka, rangelt järjestatud membraanide süsteemi lamedate mullide kujul, nn tülakoidid.
Tülakoidid kogutakse hunnikutesse - terad meenutavad mündisambaid . Granad on omavahel ühendatud stromaalsete tülakoididega, läbides neid mööda plastiidi (joon. 2.20-2.22). Klorofüll ja kloroplastid tekivad ainult kokkupuutel valgusega.

Riis. 2.20. Kloroplastid valgusmikroskoobi all

Riis. 2.21. Kloroplasti struktuur elektronmikroskoobi all

Riis. 2.22. Kloroplastide skemaatiline struktuur
Funktsioonid
1) fotosüntees(orgaaniliste ainete teke anorgaanilistest ainetest valguse energia toimel). Klorofüll mängib selles protsessis keskset rolli. See neelab valgusenergiat ja suunab selle läbi viima fotosünteesireaktsioone. Kloroplastides, nagu ka mitokondrites, sünteesitakse ATP.
2) osaleda aminohapete ja rasvhapete sünteesis,
3) toimida ajutiste tärklisevarude hoidlana.
Leukoplastid- väikesed värvitud plastiidid, mida leidub päikesevalguse eest varjatud elundite rakkudes (juured, risoomid, mugulad, seemned). Nende struktuur on sarnane kloroplastide struktuuriga. (joon. 2.23).
Erinevalt kloroplastidest on leukoplastidel aga halvasti arenenud sisemembraanisüsteem, sest nad osalevad varutoitainete – tärklise, valkude ja lipiidide – sünteesis ja kogunemises. Valguses võivad leukoplastid muutuda kloroplastideks.

Riis. 2.23. Leukoplasti struktuur
Kromoplastid- oranži, punase ja kollase värvusega plastiidid, mida põhjustavad karotenoidide rühma kuuluvad pigmendid. Kromoplaste leidub paljude taimede kroonlehtede rakkudes, küpsetes viljades, harva ka juurviljades, aga ka sügisestes lehtedes. Sisemembraani süsteem kromoplastides reeglina puudub (joonis 24).

Riis. 2.24. Kromoplasti struktuur
Kromoplastide tähtsust pole veel täielikult välja selgitatud. Enamik neist on vananevad plastiidid. Need arenevad reeglina kloroplastidest, samal ajal kui plastiidides hävivad klorofüll ja sisemembraani struktuur ning karotenoidid kogunevad. See juhtub siis, kui viljad valmivad ja lehed muutuvad sügisel kollaseks. Kromoplastide bioloogiline tähtsus seisneb selles, et need määravad lillede ja viljade erksa värvuse, mis meelitab putukaid risttolmlema ja teisi loomi vilju levitama. Leukoplastid võivad muutuda ka kromoplastideks.
Plastiidide funktsioonid
Orgaaniliste ainete süntees klorofüllis lihtsatest anorgaanilistest ühenditest: süsinikdioksiid ja vesi päikesevalguse kvantide juuresolekul - fotosüntees, ATP süntees fotosünteesi valgusfaasis
Valkude süntees ribosoomidel (kloroplasti sisemembraanide vahel on DNA, RNA ja ribosoomid, seetõttu sünteesitakse nii kloroplastides kui ka mitokondrites valku, mis on vajalik nende organellide tegevuseks).
Kromoplastide olemasolu seletatakse lillede, puuviljade ja sügislehtede võra kollase, oranži ja punase värviga.
Leukoplastid sisaldavad säilitusaineid (vartes, juurtes, mugulates).
Kloroplastid, kromoplastid ja leukoplastid on võimelised rakkudevaheliseks üleminekuks. Niisiis, kui viljad valmivad või lehtede värvus sügisel muutub, muutuvad kloroplastid kromoplastideks ja leukoplastid kloroplastideks, näiteks kui kartulimugulad muutuvad roheliseks.
Evolutsioonilises mõttes on plastiidide esmane, esialgne tüüp kloroplastid, millest pärinevad ülejäänud kahe tüübi plastiidid. Plastiididel on palju sarnasusi mitokondritega, eristades neid teistest tsütoplasma komponentidest. See on esiteks kahe membraani kest ja suhteline geneetiline autonoomia, mis on tingitud oma ribosoomide ja DNA olemasolust. See organellide iseärasus pani aluse ideele, et plastiidide ja mitokondrite eelkäijateks on bakterid, mis evolutsiooni käigus osutusid eukarüootseks rakuks ehitatud ning järk-järgult kloroplastideks ja mitokondriteks muutunud. (joon. 2.25).

Riis. 2.25. Mitokondrite ja kloroplastide teke vastavalt sümbiogeneesi teooriale
Kamber- elava süsteemi elementaarüksus. Elusraku erinevaid struktuure, mis vastutavad teatud funktsiooni täitmise eest, nimetatakse organellideks, nagu kogu organismi elundeid. Spetsiifilised funktsioonid rakus jagunevad organellide, teatud kujuga rakusiseste struktuuride vahel, nagu raku tuum, mitokondrid jne.
Rakulised struktuurid:
Tsütoplasma... Raku kohustuslik osa, mis on suletud plasmamembraani ja tuuma vahele. Tsütosool See on mitmesuguste soolade ja orgaaniliste ainete viskoosne vesilahus, mida läbib valgufilamentide süsteem - tsütoskelett. Enamik raku keemilistest ja füsioloogilistest protsessidest toimub tsütoplasmas. Struktuur: tsütosool, tsütoskelett. Funktsioonid: hõlmab erinevaid organelle, raku sisekeskkonda
Plasma membraan... Iga loomade, taimede rakk on plasmamembraaniga piiratud keskkonnast või teistest rakkudest. Selle membraani paksus on nii väike (umbes 10 nm), et seda saab näha ainult elektronmikroskoobiga.

Lipiidid need moodustavad membraanis topeltkihi ning valgud imbuvad läbi kogu selle paksuse, on sukeldatud erinevatel sügavustel lipiidikihti või paiknevad membraani välis- ja sisepinnal. Kõigi teiste organellide membraanide struktuur on sarnane plasmamembraaniga. Struktuur: kahekordne lipiidide, proteiinide, süsivesikute kiht. Funktsioonid: piiravad, säilitavad raku kuju, kaitsevad kahjustuste eest, reguleerivad ainete sissevõtmist ja eemaldamist.
Lüsosoomid... Lüsosoomid on membraani organellid. Need on ovaalse kujuga ja läbimõõduga 0,5 mikronit. Need sisaldavad ensüümide komplekti, mis hävitavad orgaanilist ainet. Lüsosoomi membraan on väga tugev ja takistab oma ensüümide tungimist raku tsütoplasmasse, kuid kui lüsosoom on mistahes välismõjude poolt kahjustatud, siis hävib kogu rakk või osa sellest.
Lüsosoome leidub kõigis taimede, loomade ja seente rakkudes.
Seedides erinevaid orgaanilisi osakesi, annavad lüsosoomid täiendavat "toorainet" rakus toimuvateks keemilisteks ja energiaprotsessideks. Nälgimise ajal seedivad lüsosoomirakud mõningaid organelle ilma rakku tapmata. See osaline seedimine varustab rakku mõneks ajaks vajaliku minimaalse toitainetega. Mõnikord seedivad lüsosoomid terveid rakke ja rakurühmi, mis mängib loomade arengus olulist rolli. Näiteks võib tuua saba kaotuse, kui kulles muutub konnaks. Struktuur: ovaalsed vesiikulid, membraan väljas, ensüümid sees. Funktsioonid: orgaanilise aine lõhestamine, surnud organellide hävitamine, kulunud rakkude hävitamine.
Golgi kompleks... Endoplasmaatilise retikulumi õõnsuste ja tuubulite luumenisse sisenevad biosünteesi saadused kontsentreeritakse ja transporditakse Golgi aparaadis. Selle organoidi suurus on 5–10 µm.

Struktuur: membraaniga ümbritsetud õõnsused (vesiikulid). Funktsioonid: akumuleerumine, pakendamine, orgaanilise aine väljutamine, lüsosoomide moodustamine
Endoplasmaatiline retikulum... Endoplasmaatiline retikulum on süsteem orgaaniliste ainete sünteesiks ja transportimiseks raku tsütoplasmas, mis on ühendatud õõnsuste ažuurne struktuur.
Endoplasmaatilise retikulumi membraanidele on kinnitatud suur hulk ribosoome – raku väikseimad organellid, mis näevad välja nagu 20 nm läbimõõduga kera. ja koosneb RNA-st ja valgust. Valkude süntees toimub ribosoomidel. Seejärel sisenevad äsja sünteesitud valgud õõnsuste ja tuubulite süsteemi, mida mööda nad raku sees liiguvad. Õõnsused, torukesed, tuubulid membraanidest, ribosoomimembraanide pinnal. Funktsioonid: orgaaniliste ainete süntees ribosoomide abil, ainete transport.
Ribosoomid... Ribosoomid kinnituvad endoplasmaatilise retikulumi membraanidele või paiknevad vabalt tsütoplasmas, paiknevad rühmadena, nendel sünteesitakse valgud. Valgu koostis, ribosomaalne RNA Funktsioonid: tagab valkude biosünteesi (valgumolekuli kokkupanek alates).
Mitokondrid... Mitokondrid on energia organellid. Mitokondrite kuju on erinev, need võivad olla ülejäänud, vardakujulised, niitjad keskmise läbimõõduga 1 mikron. ja pikkus 7 mikronit. Mitokondrite arv sõltub raku funktsionaalsest aktiivsusest ja võib putukate lendavates lihastes ulatuda kümnete tuhandeteni. Mitokondrid on väljast piiratud välismembraaniga, all on sisemine membraan, mis moodustab arvukalt väljakasvu - cristae.

Mitokondrite sees on RNA, DNA ja ribosoomid. Selle membraanidesse on ehitatud spetsiifilised ensüümid, mille abil muudetakse toiduainete energia mitokondrites raku ja kogu organismi eluks vajalikuks ATP energiaks.
Membraan, maatriks, väljakasvud - cristae. Funktsioonid: ATP molekuli süntees, oma valkude, nukleiinhapete, süsivesikute, lipiidide süntees, oma ribosoomide moodustamine.
Plastiidid... Ainult taimerakus: lekoplastid, kloroplastid, kromoplastid. Funktsioonid: orgaaniliste varuainete kogumine, tolmeldavate putukate ligimeelitamine, ATP ja süsivesikute süntees. Kloroplastid on 4–6 µm läbimõõduga ketta või kera kujulised. Topeltmembraaniga - välimine ja sisemine. Kloroplasti sees on ribosoomi DNA ja spetsiaalsed membraanistruktuurid – terad, mis on omavahel ja kloroplasti sisemembraaniga seotud. Igas kloroplastis on umbes 50 tera, mis on parema valguse püüdmise tagamiseks jaotatud. Klorofüll asub granmembraanides, tänu sellele muundub päikesevalguse energia ATP keemiliseks energiaks. ATP energiat kasutatakse kloroplastides orgaaniliste ühendite, eelkõige süsivesikute sünteesiks.
Kromoplastid... Kromoplastides leiduvad punased ja kollased pigmendid annavad erinevatele taimeosadele punase ja kollase värvuse. porgand, tomati puuviljad.
Leukoplastid on varutoitaine - tärklise - kogunemise koht. Eriti palju on leukoplaste kartulimugulate rakkudes. Valguses võivad leukoplastid muutuda kloroplastideks (selle tulemusena muutuvad kartulirakud roheliseks). Sügisel muutuvad kloroplastid kromoplastideks ning rohelised lehed ja viljad muutuvad kollaseks ja punaseks.
Raku keskus... Koosneb kahest silindrist, tsentrioolist, mis asuvad üksteisega risti. Funktsioonid: keermetugi spindlite jagamiseks

Tsütoplasmas tekivad rakulised kandmised, mis seejärel raku elutegevuse käigus kaovad.

Tihedad graanulite kujul olevad inklusioonid sisaldavad varutoitaineid (tärklis, valgud, suhkrud, rasvad) või raku jääkaineid, mida pole veel võimalik eemaldada. Kõikidel taimerakkude plastiididel on võime sünteesida ja koguda varutoitaineid. Taimerakkudes toimub varutoitainete kogunemine vakuoolidesse.
Terad, graanulid, tilgad Funktsioonid: mittepüsivad moodustised, mis talletavad orgaanilist ainet ja energiat
Tuum... Kahe membraani tuumamembraan, tuumamahl, tuum. Funktsioonid: päriliku teabe säilitamine rakus ja selle taastootmine, RNA süntees - informatiivne, transport, ribosomaalne. Tuumamembraanis on eosed, mille kaudu toimub aktiivne ainevahetus tuuma ja tsütoplasma vahel. Tuum ei salvesta pärilikku teavet mitte ainult antud raku kõigi tunnuste ja omaduste kohta, protsesside kohta, mis temaga peavad toimuma (näiteks valgusüntees), vaid ka organismi kui terviku omaduste kohta. Teave salvestatakse DNA molekulidesse, mis on kromosoomide põhiosa. Tuumas on tuum. Tuum täidab pärilikku teavet sisaldavate kromosoomide olemasolu tõttu keskuse ülesandeid, mis kontrollib kogu raku elutähtsat tegevust ja arengut.