Энэ нь нэмэлт системийн функц биш юм. Комплемент системийн уураг: шинж чанар, биологийн идэвхжил. Комплемент системийг идэвхжүүлэх сонгодог арга
Комплемент бол биеийн хамгийн чухал олон үйлдэлт системүүдийн нэг юм. Нэг талаас, энэ нь эсрэгбиеээс хамааралтай урвалын гол нөлөөлөл гэж үзэж болно. Энэ нь зөвхөн lytic болон нян устгах урвалд оролцдог төдийгүй бусад эсрэгбиеээс хамааралтай нөлөөнд оролцдог бөгөөд тэдгээрийн дотор фагоцитозын өсөлт нь in vivo дэх хамгийн чухал функцүүдийн нэг юм. Нөгөө талаас, нэмэлт нь үндсэн систем болох үрэвслийн урвалын өсгөгч болдог. Хувьслын хувьд энэ нь түүний үндсэн (анхдагч) функц бөгөөд үүнийг эсрэгбие болон бусад дархлаа судлалын механизмтай холбох шаардлагагүй юм.
Комплементийг идэвхжүүлэх үйл явцын гол үйл явдал бол сонгодог (онц чухал ач холбогдлоос нь биш зөвхөн анх нээсэн учраас нэрлэсэн) болон өөр замуудын дагуу С3 бүрэлдэхүүн хэсгийн хуваагдал юм. Хоёрдахь үндсэн цэг бол процессын боломжит гүн юм: зогсолт
Энэ нь хэд хэдэн биологийн нөлөө үзүүлэхийн зэрэгцээ C3 хуваагдах үе шатанд байгаа эсэх, эсвэл цааш гүнзгийрсэн эсэх (С5-аас С9 хүртэл). Идэвхжүүлэлтийн сүүлчийн үе шатыг ихэвчлэн терминал, эцсийн (мембран довтолгоо) гэж нэрлэдэг бөгөөд энэ нь нийтлэг, сонгодог болон альтернатив замуудын хувьд адилхан бөгөөд нэмэлтүүдийн литик функц нь үүнтэй холбоотой байдаг.
Одоогийн байдлаар плазмын 20-иос доошгүй уураг нь комплементийн системд нэгддэг. Үндсэндээ тэдгээрийг 3 бүлэгт хуваадаг. Сонгодог идэвхжүүлэлтийн зам болон эцсийн (мембран довтолгоо) үе шатанд оролцдог бүрэлдэхүүн хэсгүүдийг Clq, Clr, C1, C4, C2, C3, C5, C6, C7, C8, C9 гэж тодорхойлсон. Альтернатив идэвхжүүлэлтийн замд оролцдог уурагуудыг хүчин зүйл гэж нэрлэдэг бөгөөд C, D, R гэж нэрлэдэг. Эцэст нь, урвалын эрчмийг зохицуулдаг бүлэг уураг буюу хяналтын уургийн бүлэг нь ялгагдана: тэдгээрт C1 дарангуйлагч орно. (C1INH), C3b идэвхгүйжүүлэгч (C3bINa ), pIH хүчин зүйл - C4 - АД, анафилотоксин дарангуйлагч. Үндсэн бүрэлдэхүүн хэсгүүдийн ферментийн задралын үр дүнд үүссэн хэсгүүдийг жижиг үсгээр (жишээлбэл, C3, C3b, C3d, C5a гэх мэт) тэмдэглэнэ. Ферментийн идэвхжилтэй бүрэлдэхүүн хэсэг эсвэл фрагментийг тодорхойлохын тулд тэдгээрийн тэмдэглэгээний дээр баар байрлуулна, жишээлбэл, Cl, C42, C3bBb.
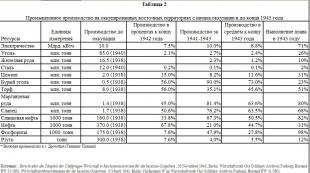
Цусны ийлдэс дэх бие даасан нэмэлт бүрэлдэхүүн хэсгүүдийн агууламж дараах байдалтай байна.
Бүрэлдэхүүн хэсгүүдийн концентраци, мкг/мл
сонгодог арга
C1 70
C1 34
C1 31
С4 600
C2 25
SZ 1200
Альтернатив зам
Үл хөдлөх хөрөнгө 25
B 200 хүчин зүйл
D хүчин зүйл 1
Мембран довтолгооны цогцолбор
C5 85
C6 75
C7 55
C8 55
C9 60
Зохицуулалтын уураг
C1 дарангуйлагч 180
H 500 хүчин зүйл
I хүчин зүйл 34
Комплемент систем нь "гох" ферментүүдийн нэг юм.
калийн систем, түүнчлэн цусны бүлэгнэлтийн систем, фибринолиз, кинин үүсэх. Энэ нь өдөөлтөд хурдан бөгөөд хурдацтай нэмэгдэж буй хариу урвалаар тодорхойлогддог. Энэхүү олшруулалт (олшруулах) нь нэг урвалын бүтээгдэхүүн нь дараагийн урвалын катализатор болж тодорхойлогддог каскадын үзэгдлийн улмаас үүсдэг. Ийм каскад нь шугаман, нэг чиглэлтэй (жишээлбэл, сонгодог нэмэлт зам), эсвэл санал хүсэлтийн гогцоо (өөр зам) байж болно. Ийнхүү хоёр хувилбар хоёулаа комплемент системд явагдана (Схем 1).
Сонгодог зам нь дархлааны цогцолбороор идэвхждэг
эсрэгтөрөгч - эсрэгтөрөгч болох IgM, IgG-ийг агуулсан эсрэгбие (3, 1, 2-р дэд ангиуд; тэдгээр нь үйл ажиллагааны буурах дарааллаар байрладаг). Үүнээс гадна сонгодог замыг IgG агрегатууд, CRP, ДНХ, плазминаар идэвхжүүлж болно. Процесс нь Clq, Clr, Cls гэсэн 3 бүрэлдэхүүн хэсгээс бүрдэх C1-ийг идэвхжүүлснээр эхэлдэг. Clq (харьцангуй молекулын жин 400) нь өвөрмөц бүтэцтэй: коллаген бариултай, коллаген бус толгойтой 6 дэд хэсэг, толгойн эсрэг талын молекулын төгсгөлд 6 саваа нийлдэг. Толгой дээр нь эсрэгбиеийн молекулуудтай хавсаргах цэгүүд байдаг бол C1G ба Cls-ийг холбох газрууд нь коллаген саваа дээр байрладаг. Clq нь AT-д хавсарсны дараа C1r нь конформацийн өөрчлөлтөөр идэвхтэй протеаз болох C1r болдог. Cls-ийг задалж, бүхэл цогцолборыг С1 серинестераза болгон хувиргадаг. Сүүлийнх нь C4-ийг 2 хэсэг болгон хуваадаг - C4a ба C4b, C2 нь C2a ба C2b. Үүссэн C4b2b(a) цогцолбор нь С3 бүрэлдэхүүн хэсгийг (сонгодог замын С3 конвертаза) задалдаг идэвхтэй фермент юм; заримдаа үүнийг C42 гэж тэмдэглэдэг.
Сонгодог замыг C1 дарангуйлагч (C1INH) зохицуулдаг бөгөөд эдгээр ферментүүдтэй эргэлт буцалтгүй холбогдож C1r ба Cls-ийн идэвхийг саатуулдаг. C1INH нь калликреин, плазмин, Хагеман хүчин зүйлийн идэвхийг бууруулдаг болохыг тогтоожээ. Энэ дарангуйлагчийн төрөлхийн дутагдал нь С4 ба С2-ийн хяналтгүй идэвхжилд хүргэдэг бөгөөд энэ нь төрөлхийн хавангийн эсрэг хэлбэрээр илэрдэг.
Альтернатив (зөвшөөрөгдсөн) зам нь Cl, C4, C2 бүрэлдэхүүн хэсгүүдийг оруулаагүй хэд хэдэн дараалсан урвалуудаас бүрдэх ба C3-ийг идэвхжүүлэхэд хүргэдэг. Үүнээс гадна эдгээр урвалууд нь эцсийн мембраны халдлагын механизмыг идэвхжүүлэхэд хүргэдэг. Грам сөрөг бактерийн эндотоксин, инулин, зимозан зэрэг зарим полисахаридууд, IgA эсвэл IgG агуулсан дархлааны цогцолборууд (ICs), зарим бактери, мөөгөнцөр (жишээлбэл, Staf. epidermis, Candida albicans) энэ замыг идэвхжүүлдэг. Урвалд дөрвөн бүрэлдэхүүн хэсэг оролцдог: D ба В хүчин зүйлүүд, С3, пропердин (P). Энэ тохиолдолд D хүчин зүйл (фермент) нь сонгодог замын Cls-тэй төстэй, C3 ба B хүчин зүйл нь C4 ба C2 бүрэлдэхүүн хэсгүүдтэй төстэй байна. Үүний үр дүнд C3bBb хувиргах өөр зам үүсдэг. Үүссэн цогцолбор нь туйлын тогтворгүй бөгөөд үүргээ гүйцэтгэхийн тулд пропердинээр тогтворжиж, илүү төвөгтэй C3bBbF цогцолбор үүсгэдэг. Альтернатив замын зохицуулалтын уураг нь piH ба C3b идэвхгүйжүүлэгч (C3JNA) юм. Эхнийх нь C3b-тэй холбогдож, идэвхгүйжүүлэгчийг (C3bINA) холбох газрыг бүрдүүлдэг. Эдгээр хүчин зүйлсийг зохиомлоор арилгах эсвэл тэдний генетикийн хомсдол нь хүмүүст саяхан тогтоогдсон нь өөр замыг хяналтгүй идэвхжүүлэхэд хүргэдэг бөгөөд энэ нь С3 эсвэл В хүчин зүйлийг бүрэн шавхахад хүргэж болзошгүй юм.
Терминал мембраны халдлагын механизм. Өмнө дурьдсанчлан, хоёр зам нь C3-бүрэлдэхүүн хэсэг дээр нийлдэг бөгөөд энэ нь үүссэн C42 эсвэл C3bBb хувиргагчаар идэвхждэг. Учир нь
С5-конвертаза үүсэхийн тулд нэмэлт C3-ийг задлах шаардлагатай. C3b нь эсийн гадаргуу дээр холбогдож, чөлөөт B, P, эсвэл p1H нь C5-ийг холбох газрыг бүрдүүлдэг бөгөөд сүүлийнх нь C3 хувиргагч бодисын протеолизэд мэдрэмтгий болгодог. Үүний зэрэгцээ жижиг C5a пептид нь C5-аас салж, үлдсэн том C5b нь эсийн мембранд наалдаж, Cb-ийг холбох газартай байдаг. Дараа нь C7, C8, C9 бүрэлдэхүүн хэсгүүдийг дараалан хавсаргана. Үүний үр дүнд эсийн цөсний давхаргаар дамжин ион ба усны хоёр талын хөдөлгөөнийг хангадаг тогтвортой мембраны суваг үүсдэг. Мембран гэмтэж, эс нь үхдэг. Тиймээс, ялангуяа харь гарагийн бичил биетнийг устгах ажиллагаа явагддаг.
Комплементыг идэвхжүүлэх явцад үрэвсэл, фагоцитоз, харшлын урвалын үйл явцад чухал үүрэг гүйцэтгэдэг олон тооны фрагментууд, пептидүүд үүсдэг.
Тиймээс Cls-ийн тусламжтайгаар C4 ба C2-ийн хуваагдал нь судасны нэвчилтийг нэмэгдүүлэхэд хүргэдэг бөгөөд C1 дарангуйлагчийн дутагдалтай холбоотой төрөлхийн хаван үүсэх эмгэгийн үндэс суурь болдог. C3a ба C5a пептидүүд нь анафилотоксины шинж чанартай байдаг. Тэд шигүү мөхлөгт эсүүд болон базофилуудтай нэгдэж гистамин ялгаруулдаг. Ц3 нь ялтастай холбогдон серотонины ялгаралтыг үүсгэдэг. C3a ба C5a-ийн анафилотоксик үйл ажиллагаа нь эдгээр пептидүүдээс аргининыг салгадаг карбоксипептидаз В-ээр амархан устдаг. Үүссэн бүтээгдэхүүнүүд нь полиморфонт эс, эозинофиль, моноцитуудтай холбоотой химиатрактантын шинж чанарыг олж авдаг. Цус задлах шинж чанаргүй C5i67 цогцолбор, В-фрагмент нь зөвхөн полиморф цөмийн лейкоцитуудад химотакси үүсгэдэг. Хүний хэвийн ийлдсэнд CFi хүчин зүйл агуулагддаг бөгөөд энэ нь полиморф цөмийн эсүүдтэй холбоотой C5a-ийн идэвхийг саатуулж, лизосомын ферментийн ялгаралтыг өдөөх чадварыг арилгадаг. Саркоидоз, Ходкины өвчтэй өвчтөнүүдэд CFi илүүдэлтэй байдаг. Энэ нь эдгээр эсийн үйл ажиллагааны доголдлыг тайлбарлаж болно. Өөр нэг C3b пептид нь полиморфонукуляр эс (PMN) болон макрофагуудад зориулсан хүчтэй опсонин юм. Энэ пептидийн рецепторууд нь бусад эсүүд (моноцит ба В-лимфоцитууд) дээр бас олдсон боловч эдгээр эсийн үйл ажиллагаанд тэдний ач холбогдол тодорхойгүй хэвээр байна. Дархлааны цогцолборын нэг хэсэг болох лимфоцитууд нэмэлтийг холбох нь дархлааны анхдагч хариу урвалыг бий болгоход чухал үүрэг гүйцэтгэдэг.
Эмнэлзүйн практикт нэмэлт системийг судлах нь өвчнийг оношлох, үйл явцын үйл ажиллагааг тодорхойлох, эмчилгээний үр нөлөөг үнэлэхэд ашиглагдана. Ямар ч үед ийлдэс дэх нэмэлт бодисын түвшин нь түүний бүрэлдэхүүн хэсгүүдийн синтез, катаболизм, хэрэглээний тэнцвэрт байдлаас хамаарна.
Комплементийн гемолитик үйл ажиллагааны бага утга нь бие даасан бүрэлдэхүүн хэсгүүдийн дутагдал эсвэл цусны эргэлтэнд түүний задралын бүтээгдэхүүн байгаа эсэхийг илэрхийлж болно. Үүнийг бас анхаарч үзэх хэрэгтэй
Гялтан хальс, үе мөчний хөндий зэрэг хэсэгт комплементийг орон нутгийн эрчимтэй хэрэглэх нь комплемент, ийлдэсийн түвшний өөрчлөлттэй хослуулж болохгүй. Жишээлбэл, ревматоид артриттай зарим өвчтөнд ийлдэс дэх нэмэлт бодисын хэмжээ хэвийн байж болох бөгөөд synovial шингэнд идэвхтэй хэрэглэснээс болж огцом буурч болно. Синовиал шингэн дэх нэмэлтийг тодорхойлох нь оношлогоонд маш чухал юм.
Төрөлхийн нэмэлт дутагдал. Комплементийн дутагдлын өв залгамжлал нь аутосомын рецессив буюу кодоминант шинж чанартай байдаг тул гетерозиготуудад комплементийн бүрэлдэхүүн хэсгүүдийн хэвийн түвшний 50 орчим хувийг эзэлдэг. Ихэнх тохиолдолд эхэн үеийн бүрэлдэхүүн хэсгүүдийн төрөлхийн дутагдал (C1, C4, C2) нь системийн чонон хөрвөстэй холбоотой байдаг. С бүрэлдэхүүн хэсгийн дутагдалтай хүмүүс дахин давтагдах пиоген халдварт өртөмтгий байдаг. Терминал бүрэлдэхүүн хэсгийн дутагдал нь гонококк ба менингококкийн халдварт өртөмтгий байдал дагалддаг. Эдгээр нэмэлт дутагдалтай үед системийн чонон хөрвөс бас тохиолддог боловч бага тохиолддог. Хамгийн түгээмэл нь төрөлхийн С2 дутагдал юм. Энэ шинж тэмдгийн гомозигот дутагдал нь чонон хөрвөс, Шонлейн-Хенох өвчин, гломерулонефрит, дерматомиозит зэрэг хэд хэдэн аутоиммун эмгэгүүдэд илэрдэг. Энэ шинж тэмдгээр гомозигот хүмүүс өөр идэвхжүүлэх зам хэвийн ажиллаж байгаа тохиолдолд халдварын мэдрэмтгий байдал нэмэгддэггүй. Бараг эрүүл хүмүүсийн дунд С2-ийн дутагдалтай гомозигот олдсон.
Гетерозигот С2-ийн дутагдал нь насанд хүрээгүй ревматоид артрит, системийн чонон ярын үрэвсэлтэй холбоотой байж болно. Гэр бүлийн судалгаагаар С2 ба С4-ийн дутагдал нь тодорхой HLA гаплотипуудтай холбоотой болохыг тогтоожээ.
Комплемент системийн зохицуулалтын уургийн дутагдал нь эмнэлзүйн шинж тэмдэгтэй байж болно. Тиймээс төрөлхийн C3INA дутагдлын үед C3-ийн дутагдалтай төстэй эмнэлзүйн зураг ажиглагдаж байна, учир нь өөр замаар хэрэглэх нь хяналтгүй болдог.
8381 0
Мембран дээр эргэлдэж, илэрхийлэгддэг 30 орчим уурагаас бүрдэх комплемент систем нь төрөлхийн болон эсрэгбиеийн дасан зохицох дархлааны хариу урвалын чухал нөлөө бүхий салбар юм. "Нэмэлт" гэсэн нэр томъёо нь энэ температурт мэдрэмтгий цусны ийлдэс материал нь эсрэгбиеийн бактерийг устгах чадварыг "нэмэлт" болохыг олж мэдсэнтэй холбоотой юм. Комплемент нь олон халдварт бичил биетнээс хамгаалахад чухал үүрэг гүйцэтгэдэг.
Хамгаалалтын үйл ажиллагааны хамгийн чухал бүрэлдэхүүн хэсгүүд нь: 1) опсониныг үйлдвэрлэх - макрофаг ба нейтрофилуудын фагоцитозын чадварыг нэмэгдүүлдэг молекулууд; 2) анафилатоксин үйлдвэрлэх - орон нутгийн болон системийн үрэвслийн урвалыг өдөөдөг пептидүүд; 3) бичил биетнийг шууд устгах.
Антигенийн өвөрмөц дархлааны хариу урвалыг сайжруулж, дархлааны цогцолбор, үхсэн эсвэл үхэж буй эсийг зайлуулж гомеостазыг (биеийн доторх тогтвортой байдлыг) хадгалах зэрэг бусад чухал нэмэлт функцүүдийг мэддэг. Комплементийн идэвхжилтийг тасалдуулах нь бие махбод дахь эс, эд эсийг гэмтээж болохыг бид мэднэ.
Нэмэлт бүрэлдэхүүн хэсгүүд нь элэг, түүнчлэн үрэвслийн хариу урвалд оролцдог эсүүдээр нийлэгждэг. Цусан дахь бүх нэмэлт уургийн концентраци ойролцоогоор 3 мг / мл байна. (Харьцуулбал: Цусан дахь IgG концентраци ойролцоогоор 12 мг/мл) Зарим нэмэлт бүрэлдэхүүн хэсгүүдийн концентраци өндөр (жишээ нь, C3-ийн хувьд 1 мг/мл орчим), бусад бүрэлдэхүүн хэсгүүд (жишээлбэл D, C2 хүчин зүйл) ул мөр байдаг. хэмжээ..
Идэвхжүүлэх замыг нөхөх
Комплементийг идэвхжүүлэх эхний үе шатууд нь түүний бүрэлдэхүүн хэсгүүдийн дараалсан каскадын идэвхжүүлэлт юм. Энэ үе шатанд нэг бүрэлдэхүүн хэсгийн идэвхжил нь ферментийн үйл ажиллагааг өдөөдөг бөгөөд энэ нь эргээд дараагийн бүрэлдэхүүн хэсгийг идэвхжүүлэхэд хүргэдэг. Нэг идэвхтэй фермент молекул нь олон субстратын молекулыг задлах чадвартай байдаг тул энэхүү урвалын цуваа нь харьцангуй сул анхны дохиог нэмэгдүүлдэг. Комплемент системийн эдгээр каскадын шинж чанарууд нь цусны бүлэгнэл үүсэх, кинин, судасны үрэвслийн зуучлагчийг үйлдвэрлэхэд чиглэсэн бусад ийлдэст ажиглагдсантай төстэй юм.Идэвхжүүлсний дараа бие даасан бүрэлдэхүүн хэсгүүдийг жижиг үсгээр тэмдэглэсэн хэсгүүдэд хуваана. Хуваасан хэсгүүдийн жижиг хэсгийг ихэвчлэн "a" үсгээр, том байх тусам "b" гэж тэмдэглэдэг. Гэсэн хэдий ч түүхэнд C2-ийн хуваагдсан хэсгүүдийн том хэсгийг ихэвчлэн C2a, жижиг хэсгийг C2b гэж нэрлэдэг. (Гэхдээ зарим зохиол, өгүүлэлд С2 нэмэлт бүрэлдэхүүн хэсгүүдийн хэсгүүдийг урвуу байдлаар тэмдэглэсэн байдаг.) Цаашид хуваагдсан хэсгүүдийг мөн жижиг үсгээр тэмдэглэсэн байдаг, жишээлбэл C3d.
Комплементийг идэвхжүүлэх гурван арга зам байдаг:сонгодог, лектин ба альтернатив.
Идэвхжүүлэх зам бүрийн эхлэл нь өөрийн бүрэлдэхүүн хэсэг, таних үйл явцаар тодорхойлогддог боловч дараагийн үе шатанд бүх гурван тохиолдолд ижил бүрэлдэхүүн хэсгүүдийг ашигладаг. Идэвхжүүлэх зам бүрийн шинж чанар, тэдгээрийг идэвхжүүлдэг бодисуудын талаар дараа нь авч үзэх болно.
сонгодог арга
Сонгодог идэвхжүүлэх замыг анх тодорхойлсон учраас ингэж нэрлэдэг. Сонгодог замын уургийн бүрэлдэхүүн хэсгүүдийг C1, C2, C9 гэж нэрлэдэг. (Тоонууд нь бүрэлдэхүүн хэсгүүдийг идэвхжүүлсэн дарааллаар нь биш харин нээсэн дарааллаар байна.) Антиген-эсрэгбиеийн цогцолборууд нь сонгодог замын гол идэвхжүүлэгчид юм. Тиймээс сүүлийнх нь хошин дасан зохицох дархлааны хариу урвалыг идэвхжүүлэх гол нөлөөлөгч зам юм.Бусад идэвхжүүлэгчид нь тодорхой вирусууд, үхсэн эсүүд болон эсийн доторх мембранууд (жишээ нь, митохондри), иммуноглобулины агрегатууд, Альцгеймерийн өвчний товруунд агуулагдах β-амилоид юм. С-реактив уураг нь цочмог фазын уураг - үрэвслийн хариу урвалын бүрэлдэхүүн хэсэг; Энэ нь олон нянгийн (жишээ нь Streptococcus pneumoniae) гадаргуу дээр илэрхийлэгддэг полисахарид фосфорилхолинд наалдаж, мөн сонгодог замыг идэвхжүүлдэг.
С1 нь нянгийн гадаргуу дээр илэрхийлэгдсэн эсрэгтөрөгчтэй холбогдсон эсрэгбие гэх мэт эсрэгтөрөгч-эсрэгбиеийн цогцолбор дахь эсрэгбиемтэй хавсарсан үед сонгодог зам эхэлдэг (Зураг 13.1). Бүрэлдэхүүн хэсэг C1 нь гурван өөр уургийн цогцолбор юм: Clq (тус бүр нь хоёр) - Clr ба Cls гэсэн хоёр молекултай холбоотой Clq (ижил зургаан дэд бүрэлдэхүүн хэсэг). Cl-ийг идэвхжүүлсний дараа түүний бөмбөрцөг мужууд - Clq-ийн дэд бүрэлдэхүүн хэсгүүд нь эсрэгтөрөгчтэй холбоотой нэг IgM эсвэл хоёр хоорондоо ойрхон байрладаг IgG молекулын аль нэгнийх нь Fc фрагментууд дээр Clq-ийн өвөрмөц бүстэй холбогддог (IgG холболтыг Зураг 13.1-д үзүүлэв).
Тиймээс IgM ба IgG эсрэгбие нь нэмэлт идэвхжүүлэгч юм. Cl-тэй холбогдож, түүнийг идэвхжүүлэх чадвартай хүний иммуноглобулинууд нь энэ чадвараа бууруулах дарааллаар: IgM>> IgG3> IgG 1 » IgG2. IgG4, IgD, IgA, IgE иммуноглобулинууд нь Clq-тай харьцдаггүй, үүнийг засахгүй, идэвхжүүлдэггүй, өөрөөр хэлбэл. сонгодог замаар комплементийг идэвхжүүлж болохгүй.
C1 нь Cls антиген-эсрэгбиеийн цогцолбортой холбогдсны дараа ферментийн идэвхийг олж авдаг. Энэ идэвхтэй хэлбэрийг Cls-эстераза гэж нэрлэдэг. Энэ нь сонгодог замын дараагийн бүрэлдэхүүн хэсэг болох C4 -ийг C4a ба C4b гэсэн хоёр хэсэгт хуваадаг. Жижиг хэсэг - C4a нь ууссан төлөвт үлддэг бөгөөд C4b нь нянгийн гадаргуу эсвэл бусад идэвхжүүлэгч бодисуудтай ковалент байдлаар холбогддог.
Дараа нь эсийн гадаргууд наалдсан C4b хэсэг нь Cls-ээр таслагдсан С2-г холбодог. C2-г задлахад ууссан төлөвт үлдэх C2b хэлтэрхий, C2a-г авна. Хариуд нь C2a нь эсийн гадаргуу дээрх C4b-тэй холбогдож C4b2a цогцолбор үүсгэдэг. Энэхүү цогцолборыг С3 конвертазын сонгодог зам гэж нэрлэдэг, учир нь бидний дараа үзэх болно, энэ фермент нь дараагийн бүрэлдэхүүн хэсэг болох С3-ийг задалдаг.
лектиний зам
Лектиний зам нь нянгийн гадаргуу дээр байрлах уураг, полисахаридын төгсгөлийн маннозын үлдэгдэлээр идэвхждэг. Эдгээр үлдэгдэл нь хөхтөн амьтдын эсийн гадаргуу дээр байдаггүй тул лектин замыг өөрийгөө болон өөрийгөө бусдыг таних хэрэгсэл гэж үзэж болно. Энэхүү идэвхжүүлэлтийн зам нь эсрэгбие байхыг шаарддаггүй тул энэ нь төрөлхийн дархлааны хамгаалалтын нэг хэсэг юм.Зураг дээр. Зураг 13.1-д бактерийн маннозын үлдэгдэл нь маннозыг холбогч лектин (MBL) цогцолбортой хэрхэн холбогддогийг харуулж байна; бүтэц нь сонгодог замын Clq-тай төстэй) болон холбогдох хоёр протеазтай холбоотой. маннозтой холбоотой серин протеазууд (MASP-1 ба -2). Энэхүү холболт нь MAP-1-ийг идэвхжүүлж, нянгийн гадаргуу дээрх сонгодог зам С3 конвертаза болох C4b2a-г үүсгэхийн тулд С4 ба С2 сонгодог комплемент замын бүрэлдэхүүн хэсгүүдийг дараа нь задалдаг. Мөн MASP-2 нь C3-ийг шууд задлах чадвартай. Тиймээс C3 идэвхжүүлэлтийн үе шатаас хойшхи лектиний зам нь сонгодог үетэй төстэй юм.
Альтернатив зам
Комплементийг идэвхжүүлэх өөр арга зам нь бараг ямар ч гадны бодисоор өдөөгддөг. Хамгийн их судлагдсан бодисууд нь липополисахаридууд (LPS, эндотоксин гэгддэг) юм. эсийн ханаграм сөрөг бактери), зарим мөөгөнцрийн эсийн хана, кобра хорд агуулагдах уураг (кобра хорны хүчин зүйл). Сонгодог зам, вирус, иммуноглобулины агрегатууд, үхсэн эсийг идэвхжүүлдэг зарим бодисууд нь өөр замыг идэвхжүүлдэг.Идэвхжүүлэлт нь тусгай эсрэгбие байхгүй үед тохиолддог. Иймд комплементийг идэвхжүүлэх өөр зам нь төрөлхийн дархлалын хамгаалалтын системийн эффектор салбар юм. Альтернатив замын зарим бүрэлдэхүүн хэсэг нь түүнд өвөрмөц байдаг (сийвэнгийн B ба D хүчин зүйл ба пропердин, Р хүчин зүйл гэж нэрлэдэг), бусад нь (C3, C3b, C5, C6, C7, C8, C9) нь сонгодог замтай хуваалцдаг.
C3b бүрэлдэхүүн хэсэг нь C3 дахь реактив тиолын бүлгийн аяндаа задралын дараа цусанд бага хэмжээгээр илэрдэг. Энэхүү "урьдчилан байгаа" C3b нь эсийн гадаргуу дээр илэрхийлэгддэг уураг ба нүүрс усны гидроксил бүлгүүдтэй холбогдох чадвартай (Зураг 13.1-ийг үз). Эсийн гадаргуу дээр C3b хуримтлагдах нь өөр замыг эхлүүлдэг.
Энэ нь гадны болон биеийн өөрийн эсэд хоёуланд нь тохиолдож болно; Тиймээс, өөр замын хувьд энэ нь үргэлж ажиллаж байна. Гэсэн хэдий ч доор илүү дэлгэрэнгүй авч үзсэний дагуу биеийн өөрийн эсүүд өөр замын урвалын явцыг зохицуулдаг бол өөрөө бус эсүүд ийм зохицуулах чадваргүй бөгөөд өөр замын дараагийн үйл явдлуудыг хөгжүүлэхээс сэргийлж чадахгүй.
Цагаан будаа. 13.1. Сонгодог, лектин ба өөр аргуудыг эхлүүлэх. Зам бүрийг идэвхжүүлж, С3 конвертаза үүсэхийг харуулах
Альтернатив замын дараагийн шатанд шар сүүний уураг болох В хүчин зүйл нь эсийн гадаргуу дээрх C3b-тэй холбогдож C3bB цогцолбор үүсгэдэг. Дараа нь D хүчин зүйл нь C3bB цогцолбор дахь эсийн гадаргуу дээр байрлах B хүчин зүйлийг задалж, хүрээлэн буй шингэнд ялгардаг Ba-ийн хэлтэрхий, C3b-тай холбоотой хэвээр байгаа Bb-ийн хэсэг үүсдэг.Энэ C3bBb нь өөр зам С3 юм. C3-ийг C3a ба C3b болгон задалдаг конвертаза.
Ихэвчлэн C3bBb хурдан уусдаг боловч пропердинтэй хослуулан тогтворжуулж болно (13.1-р зургийг үз). Үүний үр дүнд, зохих ёсоор тогтворжсон C3bBb нь маш богино хугацаанд их хэмжээний С3-ийг холбож, салгах чадвартай. Эдгээр хурдан үүссэн их хэмжээний C3b эсийн гадаргуу дээр хуримтлагдах нь өөр замыг бараг "тэсрэх" эхлүүлэхэд хүргэдэг. Тиймээс пропердиныг C3bBb-тэй холбох нь өөр зам олшруулах гогцоо үүсгэдэг. Пропердины олшруулалтын гогцоог идэвхжүүлэх чадварыг зохицуулах уургийн эсрэг үйлчлэлээр удирддаг. Тиймээс өөр замыг идэвхжүүлэх нь үргэлж тохиолддоггүй.
C3 ба C5-ийг идэвхжүүлэх
C3 задрал нь бүх гурван идэвхжүүлэлтийн замын гол үе шат юм. Зураг дээр. 13.2-аас харахад сонгодог ба альтернатив зам дахь C3 хувиргагчид (C4b2a ба C3bBb тус тус) С3-ийг хоёр фрагмент болгон хуваадаг. Жижиг C3a нь уусдаг анафилатоксины уураг бөгөөд үрэвслийн хариу урвалд оролцдог эсүүдийг идэвхжүүлдэг. Томоохон фрагмент болох C3b нь идэвхжүүлэлтийн талбайн эргэн тойронд эсийн гадаргуутай холбогдон нэмэлт каскадын идэвхжүүлэх процессыг үргэлжлүүлдэг. Доор үзүүлсэнчлэн C3b нь хостын хамгаалалт, үрэвсэл, дархлааны зохицуулалтад оролцдог.
Цагаан будаа. 13.2. Сонгодог ба лектин (дээд) ба альтернатив (доод) замд С3-конвертаза болон С5-конвертазагаар С3 бүрэлдэхүүн хэсгийг хуваах. Бүх тохиолдолд C3 нь эсийн гадаргуу дээр хуримтлагддаг C3b болон шингэн орчинд ялгардаг C3-д хуваагддаг. Үүнтэй адилаар C5 нь эсийн гадаргуу дээр хуримтлагдсан C5b болон шингэн орчинд ялгардаг C5a-д хуваагдана.
Сонгодог болон альтернатив замуудын аль алинд нь C3b-ийн С3 конвертазыг холбох нь дараагийн бүрэлдэхүүн хэсэг болох C5-ийн холболт болон хуваагдлыг эхлүүлдэг (13.2-р зургийг үз). Энэ шалтгааны улмаас C3b-тэй холбоотой C3 конвертазыг C5 хувиргагч гэж ангилдаг (сонгодог замд C4b2a3b; өөр хувилбарт C3bBb3b). С5-г таслахад хоёр хэлтэрхий үүснэ. C5a фрагмент нь уусдаг хэлбэрээр ялгардаг бөгөөд идэвхтэй анафилатоксин юм. C5b фрагмент нь эсийн гадаргуутай холбогдож, төгсгөлийн нэмэлт бүрэлдэхүүн хэсгүүдтэй холбогдох цөмийг үүсгэдэг.
терминалын зам
Комплементийн каскадын төгсгөлийн бүрэлдэхүүн хэсгүүд - C5b, C6, C7, C8 ба C9 нь идэвхжүүлэх бүх замд нийтлэг байдаг. Тэд бие биетэйгээ холбогдож, мембраны довтолгооны цогцолбор (MAC) үүсгэдэг бөгөөд энэ нь эсийн задралыг үүсгэдэг (Зураг 13.3).
Цагаан будаа. 13.3 Мембран довтолгооны цогцолбор үүсэх. Хожуу үе шатны нэмэлт бүрэлдэхүүн хэсгүүд - C5b-C9 - дараалан холбож, эсийн гадаргуу дээр цогцолбор үүсгэдэг. Олон тооны C9 бүрэлдэхүүн хэсгүүд нь энэ цогцолборт наалдаж, полимержих замаар поли-С9 үүсгэдэг бөгөөд эсийн мембраныг хамарсан суваг үүсгэдэг.
MAC үүсэх эхний үе шат нь эсийн гадаргуу дээр C6-аас C5b-д наалддаг. Дараа нь C7 нь C5b, C6-тай холбогдож, эсийн гаднах мембраныг нэвт шингээдэг. Дараа нь C8-ийг C5b67-тэй холбосноор эсийн мембран руу гүн нэвтэрдэг цогцолбор үүсдэг. Эсийн мембран дээр C5b-C8 нь C8-тэй холбогддог перфорин төрлийн молекул болох C9-ийн рецепторын үүрэг гүйцэтгэдэг.
Нэмэлт С9 молекулууд нь C9 молекултай нийлмэл харилцан үйлчилж, полимержсэн С9 (поли-С9) үүсгэдэг. Эдгээр поли-С9 нь эс дэх осмосын тэнцвэрийг алдагдуулдаг трансмембран суваг үүсгэдэг: ионууд түүгээр нэвтэрч, ус нэвтэрдэг. Эс хавдаж, мембран нь макромолекулуудыг нэвчүүлэх чадвартай болж, дараа нь эсээс гардаг. Үр дүн нь эсийн задрал юм.
Р.Койко, Д.Саншайн, Э.Бенжамини
, Коллоид тогтолцооны гоо зүй, биологи, соёлын үүрэг , 1. Мэргэжлийн үйл ажиллагаанд аюулгүй байдлын байр суурь, үүрэг..do , Мөнгө, эдийн засагт гүйцэтгэх үүрэг.docx , Хувь хүний төлөвшилд гэр бүл ямар үүрэг гүйцэтгэдэг вэ.docx , Галперин П.Я. Сэтгэлгээний аажмаар үүсэх. action.docx , EP 01 Төслийн санааны тодорхойлолт. Төслийн зорилгыг хүрээгээр бүрдүүлэх, 20-р зууны соёлд гүн ухааны байр суурь, үүрэг..docx.
Комплементийн нөлөөллийн үүрэг. Мембраны довтолгооны цогцолбор үүсэх, эсийн задралд гүйцэтгэх үүрэг.
а) бичил биетний болон бусад эсийн задралд оролцдог (цитотоксик нөлөө);
б) химиотактик үйл ажиллагаатай;
в) анафилаксийн үед оролцдог;
г) фагоцитозд оролцдог.
Комплементийн үндсэн ашигтай нөлөө:
бичил биетнийг устгахад туслах;
дархлааны цогцолборыг эрчимтэй арилгах;
хошин дархлааны хариу урвалыг идэвхжүүлэх, сайжруулах.
Комплемент систем нь дараах тохиолдолд таны биеийн эс, эд эсийг гэмтээж болно.
хэрэв түүний ерөнхий идэвхжил, жишээлбэл, грам сөрөг бактерийн улмаас үүссэн септицемитэй бол;
хэрэв түүний идэвхжил нь эд эсийн үхжил, ялангуяа миокардийн шигдээсийн голомтод тохиолддог;
хэрэв эдэд аутоиммун урвалын үед идэвхжил үүсвэл.
Эхний үе шат: эсийн гадаргуу дээр C6-аас C5b-ийн хавсралт. Дараа нь C7 нь C5b, C6-тай холбогдож, ордог гаднах мембранэсүүд. Дараа нь C8-ийг C5b67-тэй холбосноор эсийн мембран руу гүн нэвтэрдэг цогцолбор үүсдэг. Эсийн мембран дээр C5b-C8 нь C8-тэй холбогддог перфорин төрлийн молекул болох C9-ийн рецепторын үүрэг гүйцэтгэдэг. Нэмэлт С9 молекулууд нь C9 молекултай нийлмэл харилцан үйлчилж, полимержсэн С9 (поли-С9) үүсгэдэг. Тэд эс дэх осмосын тэнцвэрийг алдагдуулдаг трансмембран суваг үүсгэдэг: ионууд түүгээр нэвтэрч, ус нэвтэрдэг. Эс хавдаж, мембран нь макромолекулуудыг нэвчүүлэх чадвартай болж, дараа нь эсээс гардаг. Үүний үр дүнд эсийн задрал үүсдэг.

Магтаалын систем - цусанд байнга байдаг нарийн төвөгтэй уургийн цогцолбор. Энэ нь каскадын систем юмпротеолитик ферментүүд зориулавхошин шогийн биеийг гадны төлөөлөгчдийн үйлдлээс хамгаалах, хэрэгжүүлэхэд оролцдогдархлааны хариу урвал организм. Энэ нь төрөлхийн болон олдмол дархлааны чухал бүрэлдэхүүн хэсэг юм.

Сонгодог зам дагуу Комплемент нь эсрэгтөрөгч-эсрэгбиеийн цогцолбороор идэвхждэг. Үүний тулд нэг IgM молекул эсвэл хоёр IgG молекулын эсрэгтөрөгчийг холбоход оролцоход хангалттай. Уг процесс нь AG + AT цогцолборт С1 бүрэлдэхүүн хэсгийг нэмснээр эхэлдэг, энэ нь дэд хэсгүүдэд хуваагддагC1q, C1r болон C1s. Цаашилбал, дараалсан идэвхжүүлсэн "эрт" нэмэлт бүрэлдэхүүн хэсгүүд нь урвалд оролцдог: C4, C2, NW. С3 нэмэлтийн "эрт" бүрэлдэхүүн хэсэг нь эсийн мембранд бэхлэх чадвартай С5 бүрэлдэхүүнийг идэвхжүүлдэг. С5 бүрэлдэхүүн хэсэгт C6, C7, C8, C9 "хожуу" бүрэлдэхүүн хэсгүүдийг дараалан хавсаргаснаар мембраны бүрэн бүтэн байдлыг зөрчсөн (түүн дотор нүх үүсгэдэг) литик буюу мембраны довтолгооны цогцолбор үүсч, эс нь үхдэг. осмосын задралын үр дүн.
Альтернатив зам Комплемент идэвхжүүлэлт нь эсрэгбиеийн оролцоогүйгээр явагддаг. Энэ зам нь грам сөрөг бичил биетнээс хамгаалах онцлог шинж юм. Альтернатив зам дахь каскадын гинжин урвал нь эсрэгтөрөгчийг В уурагтай харьцахаас эхэлдэг., D ба пропердин (P) дараа нь С3 бүрэлдэхүүнийг идэвхжүүлнэ. Цаашилбал, урвал нь сонгодог аргаар явагддаг - мембраны довтолгооны цогцолбор үүсдэг.
Лектин тавьсан Комплемент идэвхжүүлэлт нь эсрэгбиеийн оролцоогүйгээр явагддаг. Энэ нь маннозыг холбогч тусгай уургаар үүсгэгддэгбичил биетний эсийн гадаргуу дээрх маннозын үлдэгдэлтэй харилцан үйлчилсний дараа C4-ийг катализ болгодог цусны ийлдэс. Цаашдын урвалын цуваа нь сонгодог аргатай төстэй.
Комплементийг идэвхжүүлэх явцад түүний бүрэлдэхүүн хэсгүүдийн протеолизийн бүтээгдэхүүнүүд үүсдэг - C3a ба C3b, C5a ба C5b дэд хэсгүүд болон биологийн өндөр идэвхжилтэй бусад. Жишээлбэл, C3a, C5a нь анафилаксийн урвалд оролцдог, химиатрактантууд, C3b нь фагоцитозын объектуудыг опсонжуулах үүрэг гүйцэтгэдэг гэх мэт. Ca ионуудын оролцоотойгоор комплекс комплемент каскадын урвал явагддаг. 2+ ба Mg 2+.
Комплементийн биологийн үүрэг
Одинцов Ю.Н., Перелмутер В.М. Комплементийн биологийн үүрэг
Одинцов Ю.Н., Перелмутер В.М.
Сибирийн Улсын Анагаах Ухааны Их Сургууль, Томск
© Одинцов Ю.Н., Перелмутер В.М.
Комплемент бол биеийн эсэргүүцлийн хамгийн чухал хүчин зүйлүүдийн нэг юм. Комплемент систем нь янз бүрийн эффектийн механизмд, ялангуяа бичил биетний лизис (нэмэлт устгах) ба опсонизацид оролцдог. Макрофагууд нэмэлтийн литик функцийг опсоник руу шилжүүлэхэд оролцож болно. Бактериозын нэмэлт үйл ажиллагаа нь халдварт өвчний эмгэг жамаас хамаардаг.
Түлхүүр үгс: комплемент, бактериолиз, опсонизаци, халдварт үйл явц.
Жинхэнэ эсэргүүцлийн үндсэн хүчин зүйлүүдийн нэг бол комплемент юм. Үүний гол үүрэг нь бактерийн задрал, фагоцитозын бактерийн опсонизаци юм. Опсоны үйл ажиллагааны литик функцийг өөрчлөх нь макрофагуудаас хамаардаг. Бактериозын үед нэмэлт үйл ажиллагаа нь халдварт өвчний патогенезийн онцлогоос хамаардаг.
Түлхүүр үгс: комплемент, бактериолиз, опсонизаци, халдварт үйл явц.
UDC 576:8.097.37
Хүний бие нь өвөрмөц бус (эсэргүүцэл) ба өвөрмөц (дархлаа) халдварт өвчний үүсгэгчээс хамгаалах хоёр үндсэн шугамтай байдаг.
Хамгаалалтын эхний шугамын хүчин зүйлүүд (эсэргүүцэл) нь хэд хэдэн нийтлэг шинж тэмдгээр тодорхойлогддог: 1) эмгэг төрүүлэгчтэй тулгарахаас өмнө үүссэн (төрөхөөс өмнөх үе); 2) өвөрмөц бус; 3) генетикийн хувьд тодорхойлогддог; 4) популяцид генотип болон фенотипийн хувьд нэг төрлийн (гетероген); 5) нэг эмгэг төрүүлэгчийн өндөр эсэргүүцлийг нөгөөд нь бага эсэргүүцэлтэй хослуулж болно; 6) эсэргүүцэл нь үндсэндээ HLA-тай холбоогүй генүүдээр хянагддаг макрофагуудын функциональ байдал, нэмэлт системийн төлөв байдлаас (HLD-ээр хянагддаг) хамаардаг.
Комплемент нь найрлага, үйл ажиллагаа нь ерөнхийдөө сайн судлагдсан олон бүрэлдэхүүн хэсэгтэй плазмын ферментийн систем бөгөөд биеийн эсэргүүцлийн хамгийн чухал хүчин зүйлүүдийн нэг юм. 1960-1970-аад онд. Эсэргүүцлийн үзүүлэлтүүдийн нэг болох комплементийн титрийг тодорхойлох нь ялангуяа түгээмэл байсан. Мөн одоогийн байдлаар олон тооны судалгааг нэмэлт функцийг судлахад зориулж байна. Гэсэн хэдий ч байдаг
нэмэлтийг идэвхжүүлэх механизмыг тайлбарлахад тодорхой бэрхшээл, зөрчилдөөн төдийгүй
нэмэлтийг идэвхжүүлэх, үйл ажиллагааны зарим механизмыг хангалттай судлаагүй байна. Ийм маргаантай асуудлууд нь in vivo-д комплемент идэвхжүүлэх дарангуйлагчдын үйл ажиллагааны механизм, комплемент идэвхжүүлэлтийг литикээс опсоны функц руу шилжүүлэх механизм, янз бүрийн халдварын үед саногенез дэх комплементийн үүргийг ойлгох зэрэг орно.
Цусны сийвэнгийн 14 уураг (бүрэлдэхүүн хэсэг) нь нэмэлт системийг бүрдүүлдэг. Эдгээр нь гепатоцит, макрофаг, нейтрофилуудаар нийлэгждэг. Тэдний ихэнх нь p-глобулинуудад хамаардаг. ДЭМБ-аас баталсан нэршлийн дагуу комплемент системийг С тэмдгээр, түүний бие даасан бүрэлдэхүүн хэсгүүдийг Cl, C2, C3, C4, C5, C6, C7, C8, C9 эсвэл том үсгээр (D, B, P). Бүрэлдэхүүн хэсгүүдийн нэг хэсэг (Cl, C2, C3, C4, C5, B) нь тэдгээрийг бүрдүүлэгч дэд бүрэлдэхүүн хэсгүүдэд хуваагддаг - илүү хүнд, ферментийн идэвхжилтэй, бага жинтэй, ферментийн идэвхгүй боловч бие даасан байдлыг хадгалдаг. биологийн функц. Комплемент системийн уургийн идэвхжүүлсэн цогцолборууд нь цогцолборын дээгүүр баараар тэмдэглэгдсэн байдаг (жишээлбэл, C4b2a3b - C5 конвертаза).
Нэмэлт уураг (C1-C9) -аас гадна биологийн үйл ажиллагааг хэрэгжүүлэхэд тэд авдаг
оролцоо болон зохицуулалтын үүргийг гүйцэтгэдэг бусад уураг:
a) комплементийн дэд бүрэлдэхүүн хэсгүүдийн макроорганизмын эсийн мембраны рецепторууд: CR1(CD35), CR2(CD21), CR3(CD11b/CD18), CR4(CD11c/CD18), C1qR, C3a/C4aR, C5aR;
б) макроорганизмын эсийн мембраны уураг: мембран кофактор уураг (MCP, эсвэл MCP - уураг задлах мембрантай холбоотой кофактор, CD46), диссоциацийг хурдасгагч хүчин зүйл (FAD, эсвэл DAF - ялзралыг хурдасгах хүчин зүйл, CD55), протеин (CD59);
в) эерэг буюу сөрөг зохицуулалтыг гүйцэтгэдэг цусны сийвэнгийн уураг: 1) эерэг зохицуулалт - хүчин зүйл B, хүчин зүйл D, properdin (P); 2) сөрөг зохицуулалт - хүчин зүйл I, хүчин зүйл H, уураг холбох C4b (C4 холбох уураг, C4bp), C1 дарангуйлагч (C1-inh, serpin), S-уураг (vitronectin).
Тиймээс 30 гаруй бүрэлдэхүүн хэсэг нь нэмэлт системийн үйл ажиллагаанд оролцдог. Комплементийн уургийн бүрэлдэхүүн хэсэг (дэд бүрэлдэхүүн хэсэг) бүр тодорхой шинж чанартай байдаг (Хүснэгт 1).
Ихэвчлэн нэмэлт бүрэлдэхүүн хэсгүүд нь сийвэн дэх идэвхгүй байдалд байдаг. Тэд олон үе шаттай идэвхжүүлэх урвалын үйл явцад идэвхтэй болдог. Идэвхжүүлсэн нэмэлт бүрэлдэхүүн хэсгүүд нь ферментийн урвалын каскадын хэлбэрээр тодорхой дарааллаар үйлчилдэг бөгөөд өмнөх идэвхжүүлэлтийн бүтээгдэхүүн нь дараагийн урвалд шинэ дэд бүрэлдэхүүн хэсэг эсвэл нэмэлт бүрэлдэхүүн хэсгийг оруулах катализатор болдог.
Комплемент систем нь янз бүрийн нөлөөллийн механизмд оролцож болно:
1) бичил биетний задрал (нэмэлт устгах);
2) бичил биетний опсонизаци;
3) дархлааны цогцолборыг задлах, тэдгээрийн цэвэрлэгээ;
4) үрэвслийн голомт руу лейкоцитыг идэвхжүүлэх, химотактик татах;
5) өвөрмөц эсрэгбиеийн өдөөлтийг сайжруулах: a) В-лимфоцит ба эсрэгтөрөгчийг илчлэх эсүүд (APCs) гадаргуу дээрх эсрэгтөрөгчийг нутагшуулах чадварыг нэмэгдүүлэх; б) В-лимфоцитуудын идэвхжих босгыг бууруулах.
Комплементийн хамгийн чухал үүрэг бол эмгэг төрүүлэгч мембраны задрал, бичил биетний опсонизаци юм.
Хүснэгт 1
Комплементыг идэвхжүүлэх сонгодог болон альтернатив замд оролцдог нэмэлт бүрэлдэхүүн хэсгүүд болон дэд бүрэлдэхүүн хэсгүүд
Бүрэлдэхүүн хэсэг (дэд бүрэлдэхүүн хэсэг) Молекулын масс, kD Дэд бүрэлдэхүүн хэсгийн цусны ийлдэс дэх концентраци, мкг/мл Үйл ажиллагаа
C1 1124 1 C1q 2 C1r 2 C1s - Ферментийн цогцолбор
Clq 460 - 80 Урт гинжин ^ эсвэл 1dM эсрэгтөрөгч-эсрэгбиеийн цогцолбортой холбогддог.
Clr 166 - 30-50 Протеаз идэвхжүүлэгч Cb
Cls 166 - 30-50 С4 ба С2-г идэвхжүүлдэг серин протеаз
C2 110 2a, 2b 15-25 С3-конвертаза (C4b2a), дараа нь сонгодог замын С5-конвертаза (C4b2a3b) хэлбэр.
SZ 190 3a, 3b 1200
С4 200 4a, 4b 350-500
C5 191 5a, 5b 75 Зорилтот эсийн мембранд нүх үүсгэдэг мембраны довтолгооны цогцолбор үүсэх.
Factor B 95 Ba, Bb 200 хэлбэр C3-конвертаза (C3bbp), дараа нь альтернатив замын C5-конвертаза (Cbbbb)
Фактор D 25 - 1
Properdin(R) 220 25 Альтернатив зам С3-конвертаза тогтворжуулагч (C3bb), H хүчин зүйлийн нөлөөн дор C3bb-ийн диссоциацийг блоклодог.
Бичил биетний нэмэлт задрал
Бичил биетний задрал нь мембраны довтолгооны цогцолбор (MAC) үүссэний үр дүнд үүсдэг.
нэмэлт бүрэлдэхүүн хэсгүүдийн нэг. MAC хэрхэн үүссэнээс хамааран комплементийг идэвхжүүлэх хэд хэдэн арга байдаг.
Комплементийг идэвхжүүлэх сонгодог (дархлааны цогцолбор) зам
Энэхүү нэмэлтийг идэвхжүүлэх замыг сонгодог гэж нэрлэдэг, учир нь энэ нь анх удаа тайлбарлагдаж байсан бөгөөд удаан хугацааны туршид өнөөг хүртэл мэдэгдэж байсан цорын ганц зам хэвээр байна. Комплементыг идэвхжүүлэх сонгодог замд эсрэгтөрөгч-эсрэгбиеийн цогцолбор (дархлааны цогцолбор (IC)) нь эхлэлийн үүрэг гүйцэтгэдэг. Комплементийг идэвхжүүлэх эхний холбоос нь С1 бүрэлдэхүүн хэсгийн С ^ дэд бүрэлдэхүүнийг дархлааны цогцолборын иммуноглобулинтай холбох явдал юм. Ялангуяа G ангиллын иммуноглобулинууд (Ig31, IgG2, IgG3, Ig4) -ээр комплементийг идэвхжүүлсэн тохиолдолд үүнийг DO хүнд гинжин хэлхээний 285, 288, 290, 292-р байрлал дахь амин хүчлийн үлдэгдэл гүйцэтгэдэг. Энэ сайтыг идэвхжүүлэх нь эсрэгтөрөгч-эсрэгбиеийн цогцолбор (AG-AT) үүссэний дараа л тохиолддог. Сонгодог замын дагуу комплементийг идэвхжүүлэх чадвар нь эрчмийг 1dM, Ig3, DO1, DO2-ээр бууруулдаг.
Комплементийн бүрэлдэхүүн хэсэг С^ нь гурван дэд нэгжээс бүрддэг (Зураг 1), тус бүр нь AG-AT цогцолбор дахь 1г-тэй холбогдох хоёр төвтэй. Тиймээс бүрэн C^ молекул нь зургаан ийм төвтэй байдаг. AG-1gM цогцолбор үүсэх үед С^ молекул нь ижил 1gM молекулын дор хаяж хоёр секундын домэйнтэй (CH2) холбогддог ба G ангиллын иммуноглобулинууд AG-AT цогцолбор үүсэхэд оролцох үед AG-^ цогцолбор дахь дор хаяж хоёр өөр молекулын ^ хоёр дахь домэйн (CH2). AG-AT-д хавсарсан C^ нь серин протеазын шинж чанарыг олж авч, C^-д хоёр C1r молекулыг идэвхжүүлж, нэгтгэхийг эхлүүлдэг. C1r нь эргээд C^-д өөр хоёр молекулыг идэвхжүүлж, нэгтгэхийг эхлүүлдэг. Идэвхжүүлсэн C^ нь серин эстеразын идэвхжилтэй.
Дараа нь С1 цогцолборын C^ нь C4-ийг том C4b хэлтэрхий, жижиг C4a хэлтэрхий болгон задалдаг. C4b нь эсийн мембраны молекулуудын амин ба гидроксил бүлгүүдтэй ковалент холбоогоор холбогддог (Зураг 2). Мембран (эсвэл AG-AT цогцолбор) гадаргуу дээр бэхлэгдсэн C4b нь C2-ийг холбодог бөгөөд энэ нь ижил серин протеаз С^-ээр ферментийн задралд орох боломжтой болдог. Үүний үр дүнд жижиг фрагмент 2b ба том фрагмент C2a үүсдэг бөгөөд тэдгээр нь мембраны гадаргуу дээр наалдсан C4b-тэй нийлж, C4b2a ферментийн цогцолборыг үүсгэдэг.
комплемент идэвхжүүлэх сонгодог замын С3-конвертаза гэж нэрлэдэг.
Цагаан будаа. Зураг 1. С1 (1d2r2e) ферментийн цогцолборын бүрэлдэхүүн хэсгүүд ба түүний эсрэгтөрөгч-эсрэгбиеийн цогцолбортой (AG-I эсвэл AG-1gM) харилцан үйлчлэл: J - пентамер мономеруудыг нэгтгэсэн гинж.
SZVV -» -SZVVR
Би-----------------
Арматурын гогцоо Зураг. 2. Сонгодог замаар нэмэлт идэвхжүүлэх
Үүссэн C3 конвертаза нь С3-тэй харилцан үйлчилж, түүнийг жижиг C3 хэсэг ба том C3b фрагмент болгон хуваана. С3-ийн сийвэн дэх концентраци нь бүх нэмэлт бүрэлдэхүүн хэсгүүдийн хамгийн өндөр нь бөгөөд нэг ферментийн цогцолбор C4b2a (С3-конвертаза) нь 1000 хүртэлх С3 молекулыг задлах чадвартай. Энэ нь мембраны гадаргуу дээр C3b-ийн өндөр концентрацийг үүсгэдэг (C3b формацийн олшруулалт). Дараа нь C3b нь C3-конвертазын нэг хэсэг болох C4b-тэй ковалент байдлаар холбогддог. Үүссэн гурван молекулын цогцолбор C4b2a3b нь C5-конвертаза юм. С5-конвертаза дахь C3b нь бичил биетний гадаргуутай ковалент байдлаар холбогддог (Зураг 2).
С5 конвертазын субстрат нь комплементийн С5 бүрэлдэхүүн хэсэг бөгөөд түүний хуваагдал нь жижиг C5a, том C5b үүсэх замаар төгсдөг. тухай-
C5b үүсэх нь мембраны довтолгооны цогцолбор үүсэхийг эхлүүлдэг. Энэ нь C5b-д нэмэлтийн C6, C7, C8, C9 бүрэлдэхүүн хэсгүүдийг дараалан нэмснээр ферментийн оролцоогүйгээр явагддаг. C5b6 нь гидрофиль, C5b67 нь мембраны липидийн давхар давхаргад шингэсэн гидрофобик цогцолбор юм. C5b67 C8-ийн хавсралт нь үүссэн C5b678 цогцолборыг мембран руу шингээнэ. Эцэст нь 14 С9 молекул C5b678 цогцолборт бэхлэгдсэн байна. Үүссэн C5b6789 нь мембраны довтолгооны цогцолбор юм. C5b6789 цогцолбор дахь С9 молекулуудын полимержилт нь мембранд нураагүй нүх үүсэхэд хүргэдэг. Ус ба N8+ нь нүх сүвээр дамжин эсэд ордог бөгөөд энэ нь эсийн задралд хүргэдэг (Зураг 3).
Ууссан нэгдлүүд
Комплементийг идэвхжүүлэх сонгодог зам дахь MAC үүсэх эрч хүч нэмэгддэг. Олшруулах гогцоо нь мембраны гадаргуутай C3b ковалент холбоо үүссэн мөчөөс эхэлдэг. Гурван нэмэлт сийвэнгийн уураг нь гогцоо үүсэхэд оролцдог: B, D, P (proper-din). D хүчин зүйлийн нөлөөн дор (серин эстераз) C3b-тэй холбогдсон уураг В нь жижиг Ba хэсэг болон том Bb фрагмент болж хуваагддаг бөгөөд энэ нь C3b-тэй холбогддог (2-р зургийг үз). C3b Bb цогцолборыг тогтворжуулагчийн үүрэг гүйцэтгэдэг пропердиныг C3bb цогцолборт нэмснээр C3-конвертаза C3bbp-ийн альтернатив зам үүсч дуусна. Альтернатив зам С3 конвертаза нь C3 молекулуудыг задалж, нэмэлт C3b үүсгэдэг бөгөөд энэ нь бүх молекул үүсэхийг баталгаажуулдаг. илүү C5 хувиргах ба эцэст нь илүү MAC. MAC үйлдэл-
гэх мэт бие даасан бөгөөд магадгүй каспасын замаар апоптозыг өдөөдөг.
Альтернатив (аяндаа) нэмэлтийг идэвхжүүлэх зам
Альтернатив замаар комплементийг идэвхжүүлэх механизм нь эх С3 молекул дахь тиоэфирийн бондын аяндаа гидролизтэй холбоотой юм. Энэ процесс нь плазмд байнга явагддаг бөгөөд C3-ийн "сул зогсолт" гэж нэрлэгддэг. C3 гидролизийн үр дүнд түүний идэвхжсэн хэлбэр болох C31 гэж нэрлэгддэг.Цаашилбал, C3i нь B хүчин зүйлийг холбодог. D хүчин зүйл нь C3iB цогцолбор дахь B хүчин зүйлийг жижиг Ба фрагмент болон том Bb фрагмент болгон хуваадаг. Үүссэн C3iBb цогцолбор нь шингэн фазын С3-конвертаза бөгөөд нэмэлтийг идэвхжүүлэх өөр зам юм. Дараа нь шингэн фазын конвертаза C3iBb нь C3-ийг C3a ба C3b болгон задалдаг. Хэрэв C3b чөлөөт хэвээр байвал усаар гидролиз болж устдаг. Хэрэв C3b нь бактерийн мембраны гадаргуутай (ямар нэгэн бичил биетний мембран) ковалент байдлаар холбогддог бол уураг задралд ордоггүй. Түүнчлэн, энэ нь өөр зам олшруулах гогцоо үүсэхийг эхлүүлдэг. Тогтмол C3b-д B хүчин зүйл холбогддог (C3b нь H хүчин зүйлээс илүү В хүчин зүйлтэй илүү хамааралтай), C3bB цогцолбор үүсдэг бөгөөд үүнээс D хүчин зүйл үүсдэг.
Багийн жижиг хэлтэрхийг салгав. C3bb цогцолборын тогтворжуулагч болох пропердиныг нэмсний дараа мембраны гадаргуутай холбогддог C3-конвертазын өөр зам болох C3bbp комплекс үүсдэг. Холбогдсон С3 конвертаза нь нэг сайт дээр нэмэлт C3b молекулуудыг хавсаргаж эхэлдэг (C3b олшруулалт), энэ нь C3b-ийн орон нутгийн хурдан хуримтлалд хүргэдэг. Цаашилбал, холбогдсон C3 конвертаза нь C3-ийг C3a ба C3b болгон задалдаг. C3b-ийн C3 конвертазын хавсралт нь C3bb3 комплекс (C3b2bb) үүсгэдэг бөгөөд энэ нь C5 конвертазын өөр зам юм. Дараа нь комплементийг идэвхжүүлэх сонгодог замтай адил С5 бүрэлдэхүүн хэсэг хуваагдан MAC үүсдэг.
Аяндаа үүсэх гидролиз
Би ___________________________Би
Олз авах гогцоо
Цагаан будаа. 4. Комплементийг идэвхжүүлэх өөр (аяндаа) зам
"сул зогсолт" идэвхжүүлэлт
Бичил биетэн
Лектин нэмэлтийг идэвхжүүлэх зам
Манноз, фукоз, глюкозамины үлдэгдэл агуулсан грам сөрөг бактерийн липополисахаридууд (LPS) нь лектин (нүүрс усыг хүчтэй холбодог шар сүүний уураг) -аар холбогдож, комплементийг идэвхжүүлэх лектиний замыг өдөөдөг. Жишээлбэл, комплементийг идэвхжүүлэх лектины замыг өдөөгч нь кальциас хамааралтай лектины гэр бүлд хамаарах С2 шиг маннан холбогч лектин (MBL) байж болно.
Энэ нь нянгийн эсийн хананы нэг хэсэг болох маннозтой нийлж, C1r ба C13-тай ижилхэн маннан холбогч лектинтэй холбоотой хоёр серин протеиназа болох MASP1 ба MASP2-тай харилцан үйлчлэх чадварыг олж авдаг.
[MSL-MASP1-MASP2] харилцан үйлчлэл нь [C^-C1r-C^] цогцолбор үүсэхтэй адил юм. Дараа нь комплемент идэвхжүүлэлт нь сонгодог замтай ижил аргаар явагддаг (Зураг 5).
4a 2b C3a C3b C5a
Олз авах гогцоо
Цагаан будаа. 5. Комплементийг идэвхжүүлэх лектиний зам (M - эсийн гадаргуугийн бүтцийн нэг хэсэг болох манноз, жишээлбэл, LPS)
Амилоид уураг, С-реактив уураг зэрэг лектины шинж чанартай пентраксины гэр бүлийн уургууд нь бактерийн эсийн хананы харгалзах субстратуудтай харилцан үйлчилж, лектиний замаар дамждаг комплементийг идэвхжүүлдэг. Тиймээс С-реактив уураг нь грам эерэг бактерийн эсийн хананд форилхолиныг идэвхжүүлдэг. Дараа нь идэвхжүүлсэн форсфорилхолин нь нэмэлт бүрэлдэхүүн хэсгүүдийг нэгтгэх сонгодог аргыг эхлүүлдэг.
Аливаа С3-конвертазын нөлөөн дор C3-аас үүссэн C3b нь зорилтот мембрантай холбогдож, C3b нэмэлт үүсэх цэг болдог. Каскадын энэ үе шатыг "олшруулах гогцоо" гэж нэрлэдэг. Комплементийг идэвхжүүлэх ямар зам байсан ч зохицуулалтын хүчин зүйлсийн аль нэгээр нь хаагдахгүй бол энэ нь нянгийн мембранд нурдаггүй нүх үүсгэдэг мембраны довтолгооны цогцолбор үүсч, улмаар үхэлд хүргэдэг.
Халдварт өвчний үед өдөөх хугацаанаас хамааран комплементийг идэвхжүүлэх альтернатив болон лектин замууд эрт байна. Эмгэг төрүүлэгч нь макроорганизмын дотоод орчинд орсны дараа эхний хэдэн цагт тэдгээрийг идэвхжүүлж болно. Комплементийг идэвхжүүлэх сонгодог арга нь хожимдсон: эсрэгбие гарч ирэх үед л "ажиллаж" эхэлдэг (1 дМ,
Идэвхжүүлэх зохицуулалтын уургийн нэмэлт
Комплементийг идэвхжүүлэх үйл явц нь мембран (Хүснэгт 2) ба плазмын (Хүснэгт 3) уургуудаар зохицуулагддаг.
Комплементийн идэвхжүүлэлтийн замууд болон MAC үүсэхийг янз бүрийн хүчин зүйлээр хааж болно.
1) сонгодог, лектин:
C1g болон C^-г холбож идэвхгүй болгодог C1 дарангуйлагчийн үйлдэл;
I, H, C4-Lp, FUD, ICD, C ^ 1 хүчин зүйлсийн нөлөөн дор сонгодог ба лектин замын (C4b2a) C3-конвертаза үүсэхийг дарах;
FUD ^55), CR1 (CD35), ICD ^46-ийн үйлчлэлээр макроорганизмын эсийн гадаргуутай нэмэлт бүрэлдэхүүн хэсгүүдийн харилцан үйлчлэлийг дарах;
2) хувилбар:
H хүчин зүйлийн нөлөөгөөр C3iBb ба C3bb цогцолборуудын диссоциаци;
H хүчин зүйл (плазм), CR1 эсвэл LAB (макроорганизмын эсийн гадаргуу дээр холбогддог) гэсэн гурван кофакторын аль нэгийн оролцоотойгоор I хүчин зүйлээр C3b хуваагдах;
FUD, CR1 эсвэл LAB-ийн нөлөөгөөр макроорганизмын эсийн гадаргуу дээр өөр замын C3-конвертаза үүсэхийг дарах.
хүснэгт 2
Мембран зохицуулалтын уураг
Эсийн (макроорганизмын эсийн мембран дээр байрладаг)
Эсүүд дээрх хүчин зүйлийн илэрхийлэл Функцийн үр дүн
CR1 ^35) В-лимфоцитууд; моноцит (макрофаг); гранулоцитууд; уутанцрын дендрит эсүүд; NK эсүүд C2-ийн C4b-ийн холболтыг дарангуйлдаг; C4b2a-ийн C4b ба 2a-д задралыг үүсгэж, хурдасгах; катаболизмын кофактор C4b хүчин зүйлийн I нөлөөн дор; катаболизм кофактор C3b I хүчин зүйлийн нөлөөн дор; c3b ялгарснаар C3bb-ийн диссоциацийг түргэсгэдэг. Биеийн өөрийн эсийн мембран дээрх аливаа замаар дамждаг комплемент идэвхжлийг дарангуйлдаг.
ICD ^46) Т-лимфоцитууд; В-лимфоцитууд; моноцит (макрофаг); гранулоцитууд; дендрит эсүүд; NK эсүүд Конвертаза үүсэхийг дарангуйлдаг: C4b2a ба C3bb; катаболизмын кофактор C4b хүчин зүйлийн I нөлөөн дор; катаболизм кофактор C3b хүчин зүйлийн нөлөөн дор I ижил
FUD ^55) Т-лимфоцитууд; В-лимфоцитууд; моноцит (макрофаг); гранулоцитууд; дендрит эсүүд; NK эсүүд; тромбоцитууд Сонгодог замын C4b2a конвертаза үүсэхийг саатуулдаг; өөр зам C3bb хувиргах үүсэхийг саатуулдаг; C2-ийн C4b-ийн холболтыг саатуулдаг; C4b2a-ийн задралыг C4b ба 2a болгон хурдасгадаг; c3b ялгарснаар C3bb-ийн диссоциацийг хурдасгадаг
Протектин (L59) Бүх эсүүд макро- 5b678-тэй холбогдож, мембранд нэвчихийг дарангуйлдаг.
организм | болон C9 |-г байршуулах өөрийн эсүүд
Хүснэгт H
Плазмын зохицуулалтын уураг
Хүчин зүйлийн функц Молекулын жин ба ийлдэс дэх концентраци Соматик эс ба (эсвэл) эмгэг төрүүлэгчид үзүүлэх нөлөө
H хүчин зүйл (макроорганизмын эсийн гадаргуу дээрх сиалийн хүчлүүдтэй амархан холбогддог) Сонгодог замын C4b2a конвертаза үүсэхийг дарангуйлдаг; C3bBb конвертазын өөр зам үүсэхийг саатуулдаг; шингэн фазын C3iBb конвертазыг C3i ба Bb болгон задлахад хүргэдэг; катаболизмын кофактор C3i ба Bb; C3bBb конвертазыг C3b болон Bb болгон задлахад хүргэдэг 150 Кда, 500 мкг/мл
Фактор I (плазмын протеаз) Сонгодог зам үүсэхийг дарангуйлдаг C4b2a конвертаза 90 Кда, 35 мкг/мл
Кофакторуудын аль нэгтэй хамт (ICB, CR1, C4bp) 4b-ийг C4c ба C4d болгон хуваана; кофакторуудын аль нэгтэй хамт (MCB, CR1, H) C3b-ийг тасалдаг; катаболизмын хүчин зүйл C3b ба C3i Биеийн өөрийн эсийн мембран дээрх аливаа замаар дамждаг комплементийн идэвхжлийг дарангуйлдаг.
C4bp (C4 холбох уураг, уураг холбох C4b) C2-ийг C4b-тэй холбохыг дарангуйлдаг; сонгодог замын конвертаза C4b2a үүсэхийг саатуулдаг; C4b2a-г C4b ба 2a болгон задлахад хүргэдэг; Катаболизмын кофактор C4b хүчин зүйлийн нөлөөн дор I 560 Кда, 250 мкг/мл
C1 дарангуйлагч (C 1-inh, serpin) C1r ба C1s-ийг холбож, дарангуйлдаг (серин протеазын дарангуйлагч); C1q-аас C1r ба C1s-ийг салгадаг (C1q нь Ig-ийн Fc фрагменттэй холбоотой хэвээр байна); C1 s-ийн C4 ба C2-тэй харилцах хугацааг хязгаарладаг; Цусны сийвэн дэх С1-ийн аяндаа идэвхжихийг хязгаарладаг 110 Кда, 180 мкг/мл
S-уураг (витронектин) 5b67-S цогцолбор үүсгэж, мембраны липидийн давхаргад нэвчих чадварыг идэвхгүйжүүлнэ. 85 Кда, 500 мкг/мл МАК үүсэхийг хориглоно.
MAC үүсэхийг дарах Үүний эсрэгээр плазмын гаралтай зохицуулалтын уураг
ионууд нь зөвхөн соматик эсийн гадаргуу дээр төдийгүй эмгэг төрүүлэгч бичил биетний мембран дээр нэмэлт идэвхжүүлэлтийг саатуулдаг.
Комплемент бүрэлдэхүүн хэсгүүдээр бичил биетнийг опсонизаци хийх
Бичил биетний нэмэлт задрал нь макроорганизмын дотоод орчинд эмгэг төрүүлэгч бичил биетнийг нэвтрүүлэхэд үзүүлэх эрт хариу үйлдэл юм. Альтернатив буюу лектин замаар дамждаг комплементийг идэвхжүүлэх явцад үүссэн C2b, C3a, C4a, C5a, Ba дэд бүрэлдэхүүн хэсгүүд нь үрэвслийн голомт руу эсийг татан авч, тэдгээрийн эффектор үйл ажиллагааг идэвхжүүлдэг.
Нэмэлт бүрэлдэхүүн хэсгүүдээс 3b ба 4b нь голчлон опсонжуулах шинж чанартай байдаг. Тэдний үүсэхэд хоёр нөхцөл шаардлагатай: эхнийх нь дээр дурдсан замуудын аль нэгээр нь комплементийг идэвхжүүлэх, хоёр дахь нь идэвхжүүлэх процессыг хориглох бөгөөд энэ нь MAC үүсэх, эмгэг төрөгч задрах боломжгүй болгодог. Энэ нь үүнээс бүрддэг
эмгэг төрүүлэгчдийн гадаргуу дээр.
1. Мембраны липидийн давхар давхаргад нэгдэж эхэлдэг гидрофобик комплекс C5b67 нь S-уураг (витронектин) -ээр идэвхгүй болно. Үүссэн 5b67S цогцолборыг мембраны липидийн давхаргад нэвтрүүлэх боломжгүй.
2. 8-р бүрэлдэхүүн хэсгийн C5b67 цогцолборын хавсралт шингэн үе шатбага нягтралтай липопротеинууд (LDL) -ээр хааж болно.
3. C5b678-ийн мембранд дүрж, С9-ийг хавсаргаснаар макроорганизмын эсийн мембраны уураг болох CD59 (протектин) үүсэхээс сэргийлдэг.
4. Баригдсан MAC бүхий макроорганизмын эсийн мембраны хэсгүүдийг эндоцитоз эсвэл экзоцитозоор зайлуулах.
Тиймээс эсийн гаралтай зохицуулалтын уураг нь зөвхөн соматик эсийн гадаргуу дээр MAC үүсэх замаар комплементийн идэвхжилтийг бие даан дарангуйлдаг бөгөөд литикийг дарангуйлахад үр дүнтэй байдаггүй.
Макроорганизмын эсүүд дээр C3b мембраны задралын мембраны дэд бүрэлдэхүүн хэсэг болох C3b мембраны харгалзах рецепторууд байдаг (Хүснэгт 4). C3b ба идэвхгүйжүүлсэн C3b (C3b) нь нейтрофил, моноцит (макрофаг), хүйн эндотелид байрлах CR1 (C3b, C3b), CR3 (C3b), CR4 (C3b) рецепторуудын лиганд юм. СЗЬ ба СЗЫ нь идэвхтэй опсонины үүрэг гүйцэтгэдэг.
I болон H хүчин зүйлсийн хосолсон үйлдэл нь литик цогцолбор үүсэхийг (MAC, нэмэлт аллага) эмгэг төрүүлэгчийг устгах өөр механизм болох фагоцит устгах (Зураг 6) руу шилжүүлж болно. Үрэвслийн голомтод хожим гарч ирэх макрофагуудаас үүссэн комплемент идэвхжүүлэлтийн уусдаг дарангуйлагч (I ба H) нь фагоцитийн бичил орчинд үйлчилж, бактерийн гадаргуу дээр С3 конвертаза үүсэхээс сэргийлж, улмаар "чөлөөт" C3b байгаа эсэхийг баталгаажуулдаг. C3b-ийн макрофаг рецептор нь лигандыг (C3b) холбож, макрофагын гадаргуу дээр бактерийг тогтооно. Түүний фагоцитоз нь хоёр лиганд-рецепторын цогцолборын хамтарсан оролцоотой явагддаг: C3b + C3b ба FcyR + ^ рецептор. Нөгөө хос - C3b + C3 рецептор нь эсрэгбиемүүдийн оролцоогүйгээр фагоцитозыг эхлүүлдэг.
Комплемент идэвхжүүлэлтийг литикээс опсоны функц руу шилжүүлэхийн биологийн утга нь фагоциттой тулгарахаасаа өмнө задлаагүй бүх бактери C3b-опсониноор фагоцитозжих ёстой гэсэн үг юм. Комплемент идэвхжүүлэлтийг опсоник руу шилжүүлэх ийм механизм нь халдварын эхний үе шатанд амьдрах чадвартай эмгэг төрүүлэгчдийн фагоцитоз төдийгүй бичил биетний фрагментийг фагоцитоор ашиглахад шаардлагатай байдаг.
Хүснэгт 4
Нэмэлт дэд бүрэлдэхүүн хэсгүүдийн рецепторууд
Рецептор (комплемент рецептор, CR) Лигандууд Эсүүд дээрх илэрхийлэл Холбох нөлөө
CR1 (CD35) C3bi > C3b, C4b Нейтрофил, моноцит (макрофаг), В-лимфоцит, уутанцрын дендрит эс, эритроцит, бөөрний бөөрөнхий хучуур эд.
CR3 (CD11b/CD18) C3bi Нейтрофил, моноцит (макрофаг), NK эс, уутанцрын дендрит эс Опсонжуулсан фагоцитоз
CR4 (х 150-95) (CD11c/CD18) C3bi нейтрофилууд Опсонжуулсан фагоцитоз
CR2 (CD21), В-лимфоцитын гол рецепторын цогцолборын бүрэлдэхүүн хэсэг (BCR + CD19, CR2, CD81) C3bi, C3dg В-эсүүд, уутанцрын дендрит эсүүд BCR идэвхжүүлэх урвалыг сайжруулж, AG-AT цогцолборын фагоцитозгүй холболтыг өдөөдөг. уутанцрын дендрит эсүүд дээр
комплемент идэвхжүүлэх литик програмыг опсоник програм руу шилжүүлэх.
Халдварт үйл явцын бодит нөхцөлд эмгэг төрүүлэгчийн фагоцитоз, дархлааны цогцолборыг цэвэрлэх боломжийг олгодог опсоны нэмэлтийг идэвхжүүлэх хөтөлбөрт шилжих нь зохицуулалтын уургийн нөлөөллөөс болж тохиолдож болно. Мембран дээрх нэмэлт бүрэлдэхүүн хэсгүүдийн угсралт нь мембраны довтолгооны цогцолбор үүсэх замаар дуусч болно, эсвэл I ба H хүчин зүйлийн нөлөөгөөр 4b үүсэх түвшинд, 3b үүсэх түвшинд бүр илүү идэвхтэй тасалддаг.
I хүчин зүйл нь C3b-ийг задалдаг гол фермент юм. Энэ үйл явц дахь H хүчин зүйл нь кофакторын үүрэг гүйцэтгэдэг. Хамтдаа үйлчилснээр тэдгээр нь шингэний фаз ба мембран C3b-ийг (чөлөөт эсвэл аливаа конвертазын нэг хэсэг болгон) идэвхгүй болгож, түүнээс C3f фрагментийг салгах чадвартай (идэвхгүй болгосон C3b нь C3b гэж нэрлэгддэг). Дараа нь тэд C3-ийг дараах байдлаар хуваана.
φ ^ дэд бүрэлдэхүүн хэсэг
sz z z z z
Нэмэлт нэмэлт идэвхжүүлэлтийг блоклох
Бактери
Фагоцитозын процесс руу шилжих
H хүчин зүйл (кофактор)
Макрофаг
Бактерийн шингээлт
Y Pc фрагментийн X,1 C3b нэмэлт бүрэлдэхүүн хэсгийн рецептор
1| |1 В Комплементийн C3b эсвэл C33 бүрэлдэхүүн хэсгийн рецептор
Цагаан будаа. 6. Комплемент идэвхжүүлэлтийг фагоцитоз руу шилжүүлэх
Саногенезийн механизмаас хамааран өмнө нь тусгаарлагдсан бактериозын янз бүрийн бүлгийн эмгэг төрүүлэхэд комплементийн үүрэг гүйцэтгэх боломжтой асуултыг авч үзэх нь зүйтэй юм.
Хортой бактериоз (сахуу, хийн гангрена, ботулизм, татран гэх мэт). Эмгэг төрүүлэгчдийн ердийн нутагшуулалт нь халдварын орох хаалга юм. Эмгэг төрүүлэх гол нөлөөлөгч нь хорт бодис (Т-аас хамааралтай антиген, эхний төрлийн эсрэгтөрөгч) юм. Эдгээр бактерийн Т-аас хамааралтай гадаргуугийн антигенууд нь дархлааны хариу урвалыг өдөөхөд чухал үүрэг гүйцэтгэдэг. Саногенезийн гол нөлөөлөгч нь антитоксин юм.Дархлалын хариу урвалын төрөл нь T1l2. Сэргээх нь дархлааны цогцолбор үүсэх, дараа нь устгах, мөн үрэвслийн голомт дахь нянгийн фагоцитик устгалын улмаас үүсдэг. Эдгээр бактериоз дахь нэмэлтүүдийн үүрэг нь токсин-антитоксины дархлааны цогцолборыг устгахад оролцохоор хязгаарлагддаг. Комплемент нь хорт бодисыг саармагжуулахад чухал үүрэг гүйцэтгэдэггүй (жишээлбэл, токсиген халдварын саногенезид).
Хортой бус грануломатоз бактериозууд
1. Эмгэг төрүүлэгчид нь гадаргуугийн Т-бие даасан эсрэгтөрөгч (T "1 антиген, хоёр дахь төрлийн эсрэгтөрөгч) агуулдаг.
Бактери нь сонгодог LPS (enteropathogenic Escherichia coli, Salmonella, Shigella гэх мэт тантиген) агуулдаг. Эмгэг төрүүлэгчдийн ердийн нутагшуулалт нь гэдэсний салст бүрхэвчийн орох хаалганаас бүс нутгийн тунгалагийн зангилаа хүртэл байдаг. Эмгэг төрүүлэх гол хүчин зүйл нь эндотоксин ба амьд бактери юм. Дархлалын хариу урвалын төрөл нь T1l2 юм. Дархлаатай
LPS-ийн хариу урвал нь IgM ангиллын эсрэгбиемүүдийн үйлдвэрлэлээр тодорхойлогддог. Саногенез нь үндсэндээ лектин ба комплементийг идэвхжүүлэх өөр замуудын улмаас халдварт үйл явцын дархлааны өмнөх үе шатанд бактериудыг фагоцит бус аргаар устгасны үр дүнд үүсдэг. Халдварт үйл явцын дархлааны үе шатанд - идэвхжих сонгодог замын дагуу 1дМ ба комплементийн оролцоотойгоор дархлааны задралын улмаас. Энэ бүлгийн бактериозын саногенезид фагоцитоз чухал биш юм. Эдгээр өвчний үед нэмэлт системийг идэвхжүүлэх нь саногенезид хувь нэмэр оруулах боломжтой;
Бактери нь гадаргуугийн (капсул) 7!-антиген (пневмококк, гемофил бактери гэх мэт) агуулдаг. Эмгэг төрүүлэгчдийн ердийн нутагшуулалт - амьсгалын замын салст бүрхэвчийн орох хаалганаас бүс нутгийн тунгалгийн зангилаа хүртэл ихэвчлэн цусанд нэвчдэг. Эмгэг төрүүлэх гол хүчин зүйл бол амьд бактери юм. Дархлалын хариу урвалын төрөл нь T1l2 юм. Гадаргуугийн эсрэгтөрөгчийн дархлааны хариу урвалд IgM ангиллын эсрэгбие үүсдэг. Саногенез нь үндсэндээ лектин ба комплементийг идэвхжүүлэх өөр замуудын улмаас халдварт үйл явцын дархлааны өмнөх үе шатанд бактериудыг фагоцит бус аргаар устгадагтай холбоотой юм. Халдварт үйл явцын дархлааны үе шатанд - идэвхжих сонгодог замын дагуу 1дМ ба комплементийн оролцоотойгоор дархлааны задралын улмаас. Энэ бүлгийн нянгууд цусанд нэвтэрсэн тохиолдолд сул опсонжуулсан (эсвэл опсонжоогүй) бактерийн фагоцитозын гол цэг болох дэлүү нь макроорганизмыг эмгэг төрүүлэгч бичил биетнээс цэвэрлэх гол үүрэг гүйцэтгэдэг.
ШМ нь Купфер эсүүдийн фагоцитозын мэдрэмтгий нянгуудыг "онилж", улмаар цөсний хялгасан судсанд бүрэн задарч амжаагүй байгаа бактерийн хэсгүүдийг шилжүүлдэг. Цөсний давс нь гэдэс рүү ялгардаг бактерийн хэсгүүдийг задалдаг. Энэ бүлгийн өвчний нэмэлт системийг идэвхжүүлэх нь саногенезид хувь нэмэр оруулдаг.
2. Эмгэг төрүүлэгчид нь гадаргуугийн Т-аас хамааралтай эсрэгтөрөгч (T-антиген, эхний төрлийн эсрэгтөрөгч) агуулдаг.
Эмгэг төрүүлэгчдийг нутагшуулах (стафилококк, стрептококк гэх мэт) - орох хаалга (арьс, салст бүрхэвч), бүс нутгийн тунгалгийн булчирхай, системийн гэмтэл (эрхтэн). Эмгэг төрүүлэх гол хүчин зүйл нь амьд бактери, бага хэмжээгээр тэдгээрийн хорт бодис юм. Дархлааны хариу урвалд!dM-ийн DO-ийн нийлэгжилтийн өөрчлөлт тодорхой харагдаж байна. Халдварт өвчний хангалттай явцтай (дархлал хомсдолын шинж тэмдэггүй өвчтөнүүдэд) дархлааны хариу урвалын төрөл нь T1r2 юм. Саногенез нь дархлааны фагоцитоз, дархлааны задрал, антитоксины нөлөөгөөр явагддаг. Эдгээр халдварын үед дархлааны өмнөх үе шатанд саногенезийг комплемент идэвхжүүлэх бүтээгдэхүүнээр бактерийг опсонжуулах, дараа нь фагоцитоз хийх өөр замаар явуулдаг. Халдварт үйл явцын дархлааны үе шатанд саногенез нь!dM ба DO оролцдог комплемент идэвхжүүлэлтийн сонгодог замд нэмэлт аллага, түүнчлэн комплемент идэвхжүүлэх бүтээгдэхүүн болон DO-ээр опсонжуулсан бактерийн фагоцитозтой холбоотой.
Грануломатоз бактериоз
1. Эпителиоид бус эсийн мөхлөгт хэлбэрийн цочмог бактерийн эмгэг төрүүлэгчид (listeria, salmonella typhoid, paratyphoid A, B гэх мэт).
Эмгэг төрүүлэгчид нь гадаргуугийн Т-аас хамааралтай эсрэгтөрөгчийг агуулдаг. Өвчин үүсгэгч хүчин зүйл бол амьд бактери юм. Фагоцитоз бүрэн бус. Дархлалын хариу урвалын төрөл нь T1r2 ба TM. dM-ийн харагдах байдал нь гранулом үүсэх дагалддаг. dM-ийг DO болгон өөрчлөх нь грануломын урвуу хөгжилд хүргэдэг. Саногенез нь комплементийг идэвхжүүлэх, нянгийн опсонизаци хийх өөр замаар явагддаг бөгөөд тэдгээрийн дараагийн фагоцитозын хамт комплементийг идэвхжүүлдэг бүтээгдэхүүн юм. Халдварт үйл явцын дархлааны үе шатанд саногенез нь!dM ба DO оролцдог комплемент идэвхжүүлэлтийн сонгодог замд нэмэлт аллага, түүнчлэн комплемент идэвхжүүлэх бүтээгдэхүүн болон DO-ээр опсонжуулсан бактерийн фагоцитозтой холбоотой.
2. Архаг эпителоид эсийн грануломатоз бактерийн үүсгэгч бодисууд (микобактерийн сүрьеэ, уяман; бруцелла гэх мэт).
Эмгэг төрүүлэгчид нь гадаргуугийн Т-аас хамааралтай эсрэгтөрөгчийг агуулдаг. Өвчин үүсгэгч хүчин зүйл бол амьд бактери юм. Фагоцитоз бүрэн бус. Дархлалын хариу урвалын төрөл - Th2 ба Th1. IgM-ийн харагдах байдал нь гранулом үүсэх гол хүчин зүйл болж магадгүй юм. Thl-set цитокинуудын үйлдэл нь фагоцитозыг дуусгахад хангалтгүй бөгөөд энэ нь гранулом дахь эпителоид эсүүд үүсэхэд хүргэдэг. Саногенез дэх комплементийг идэвхжүүлэх хувилбаруудын аль нь ч чухал үүрэг гүйцэтгэдэггүй.
Дүгнэлт
Комплемент (комплемент систем) нь эмгэг төрөгч макроорганизмын дотоод орчинд ороход тулгардаг анхны хошин хүчин зүйлүүдийн нэг юм. Комплементийн бүрэлдэхүүн хэсгүүдийг идэвхжүүлэх механизм нь үүнийг эмгэг төрүүлэгчдийг задлах, фагоцитозыг нэмэгдүүлэхэд ашиглах боломжийг олгодог. Бүх бактерийн халдварт өвчнийг цусан дахь нэмэлт бодисын агууламж, түвшинг тодорхойлох урьдчилсан шинжилгээ болгон ашиглаж болохгүй.
Уран зохиол
1. Одинцов Ю.Н., Перелмутер В.М., Климентева Т.К. Туфтсин: мөхлөгт бус ба мөхлөгт бактерийн хөгжилд гүйцэтгэх үүрэг // Бул. ах. эм. 2002. V. 1. No 3. S. 98-102.
2. Перелмутер В.М., Одинцов Ю.Н. М ангиллын иммуноглобулины (IgM) гол үүрэг нь бактери ба тэдгээрийн эсрэгтөрөгчийн гемато-эдийн саадыг нэвтрүүлэх чадварыг зохицуулах явдал юм // Bul. ах. эм. 2005. V. 4. No 3. S. 38-42.
3. Ройт А. Дархлаа судлалын үндэс. Пер. англи хэлнээс. М.: Мир, 1991. 328 х.
4. Roit A, Brostoff J, Mail D. Immunology. Пер. англи хэлнээс. М.: Мир, 2000. 581 х.
5. Хайтов Р.М., Игнатьева Г.А., Сидорович И.Г. Дархлаа судлал. Москва: Анагаах ухаан, 2000. 432 х.
6.Ярилин АА Дархлаа судлалын үндэс. Москва: Анагаах ухаан, 1999. 607 х.
7. Alban S., Classen B., Brunner G., Blaschek W. Echinacea purpurea болон гепаринаас гаралтай арабиногалактан-уургийн нэмэлт модуляцын нөлөөг ялгах // Planta Med. 2002. V. 68(12). P. 1118-1124.
8. Ambrosio A.R., De Messias-Reason I.J. Leishmania (Viannia) braziliensis: маннозыг холбогч лектин нь гадаргуугийн гликоконьюгатуудтай харилцан үйлчлэлцэх ба комплемент идэвхжүүлэлт. Эсрэгбиеээс хамааралгүй хамгаалалтын механизм // Паразит иммунол. 2005. V. 27. P. 333-340.
9. Андерссон Ж., Ларссон Р, Рихтер Р. гэх мэт. Комплемент идэвхжүүлэлтийн загвар зохицуулагчийг (RCA) биоматериалын гадаргуутай холбох: гадаргуутай холбоотой H хүчин зүйл нь комплемент идэвхжүүлэлтийг саатуулдаг // Биоматериал. 2001. V. 22. P. 2435-2443.
10. Bohana-Kashtan O., Ziporen L, Donin N. et al. Комплементаар дамжсан эсийн дохио // Моль. Иммунол. 2004. V. 41. P. 583-597.
11. Bohlson S.S., Strasser J.A., Bower J.J., Schorey J.S. Mycobacterium avium-ийн эмгэг жам дахь комплементийн үүрэг: C3 нэмэлт бүрэлдэхүүн хэсэг байхгүй тохиолдолд халдварын эсрэг хариу урвалын in vivo болон in vitro шинжилгээ // Халдвар. Иммунол. 2001. V. 69. P. 7729-7735.
12. Brown J.S., Hussell T, Gilliland S.M. гэх мэт. Сонгодог зам нь хулганад Streptococcus pneumoniae халдварын төрөлхийн дархлааг бий болгоход шаардлагатай давамгайлах нэмэлт зам юм // Proc. Натл. Акад. шинжлэх ухаан. АНУ. 2002. V. 99. P. 16969-16974.
13. Caragine T.A., Okada N., Frey A.B., Tomlinson S. Эрт боловч хожуу биш нэмэлт литик замын хорт хавдраар илэрхийлэгддэг дарангуйлагч нь хүний хөхний хорт хавдрын харх загварт хавдрын өсөлтийг нэмэгдүүлдэг // Хорт хавдар. 2002. V. 62. P. 1110-1115.
14. Celik I., Stover C, Botto M. et al. Туршилтаар өдөөгдсөн полимикробын перитонит дахь комплементийг идэвхжүүлэх сонгодог замын үүрэг // Халдвар. Дархлаа. 2001. V. 69. P. 7304-7309.
15. Donin N, Jurianz K., Ziporen L. et al. Хүний хорт хавдрын эсийн нэмэлт эсэргүүцэл нь мембран зохицуулалтын уураг, уургийн киназ ба сиалийн хүчилээс хамаардаг // Клин. Exp. Иммунол. 2003. V. 131. P. 254-263.
16. Ферни-Кинг Б.А., Сейли Дж., Виллерс Ч. гэх мэт. Стрептококкийн нэмэлт дарангуйлагч (SIC) нь эсийн мембран дээр c567 шингээхээс сэргийлж мембраны халдлагын цогцолборыг дарангуйлдаг // Дархлаа судлал. 2001. V. 103. Дугаар 3. P. 390-408.
17. Frumeaux-Bacchi V., Dragon-Durey M.A., Blouin J. et al. Эмнэлзүйн практикт комплемент системийг судлах // Анн. Мед. Олон улсын (Парис). 2003. V. 154. P. 529-540.
18. Imai M., Ohta R., Okada N, Tomlinson S. In vivo нэмэлт зохицуулагчийг дарангуйлах нь хөхний булчирхайн аденокарцинома, Int загварт эсрэгбиеийн эмчилгээг сайжруулдаг. J. Хорт хавдар. 2004. V. 110. P. 875-881.
19. Jiang H, WagnerE, Zhang H, Frank M.M. Комплемент 1 дарангуйлагч нь нэмэлт нэмэлт замын зохицуулагч юм // J. Exp. Мед.
2001. V. 194. No 11. P. 1609-1616.
20. Langeggen H, Berge K.E., Johnson E, Hetland G. Хүний хүйн венийн эндотелийн эсүүд нь нэмэлт рецептор 1 (CD35) болон нэмэлт рецептор 4 (CD11c/CD18) in vitro // Үрэвсэл.
2002. V. 26. No 3. P. 103 - 110.
21. Laufer J., Katz Y., Passwell J.H. Үрэвслийн үед нэмэлт уургийн элэгний гаднах синтез // Моль. Иммунол. 2001. V. 38. P. 221-229.
22. Leslie R.G.Q., Nielsen C.H. Комплемент идэвхжүүлэх сонгодог ба өөр замууд нь хүний В лимфоцитууд дээр аяндаа С3 фрагмент хуримтлагдах, мембраны довтолгооны цогцолбор (MAC) үүсэхэд тодорхой үүрэг гүйцэтгэдэг // Дархлаа судлал. 2004. V. 111. Дугаар 1. P. 86-98.
23. Lukas T.J., MunozH., Erickson B.W. Хүний иммуноглобулины G-ийн хоёр дахь тогтмол домэйноос мономер ба димерик пептидүүдээр С1-зуучлагдсан дархлааны гемолизийг дарангуйлах // J. Immunology. 1981. V. 127. No 6. P. 2555-2560.
24. Наута А.Ж., Даха М.Р., Тижсма О. нар. Комплементийн мембраны довтолгооны цогцолбор нь каспазын идэвхжүүлэлт ба апоптозыг өдөөдөг // Europ. Дархлааны Ж. 2002. V. 32. Дугаар 3. P. 783-792.
25. Nielsen C.H., Marquait H.V., Prodinger W.M., Leslie R.G. CR2-ийн зуучлалын замаар нөхөх замыг идэвхжүүлснээр хүний В лимфоцитууд дээр мембраны довтолгооны альтернатив цогцолборууд үүсдэг // Иммунол. 2001. V. 104. P. 418-422.
26. Nielsen C.H., Pedersen M.L., Marquart H.V. гэх мэт. Хүний хэвийн захын В эсүүд дээр C3 фрагментийн хуримтлал, мембраны довтолгооны цогцолбор үүсэхийг дэмжихэд 1 (CR1, CD35) ба 2 (CR2, CD21) төрлийн нэмэлт рецепторуудын үүрэг // Eur. J. Immunol. 2002. V. 32. P. 1359-1367.
27. Ren B., McCrory M.A., Pass C. et al. Streptococcus pneumoniae гадаргуугийн уураг А-ийн хоруу чанарын функц нь комплементийн идэвхжүүлэлтийг дарангуйлах, комплемент рецептороор дамжих хамгаалалтыг сулруулдаг // J. Immunol. 2004. V. 173. P. 7506-7512.
28. Roos A., Ramwadhdoebe T.H., Nauta A.J. гэх мэт. Комплементийг идэвхжүүлэх эхний үе шатыг эмчилгээний дарангуйлах // Immunobiology. 2002. V. 205. P. 595-609.
29. Roos A., Bouwman L.H., Munoz J. et al. Хүний ийлдэс дэх комплементийн лектин замын функциональ шинж чанар // Мол. Иммунол. 2003. V. 39. P. 655-668.
30. Song H, He C., Knaak C. et al. Комплемент рецептор 2-зуучлах замаар комплемент дарангуйлагчдыг комплемент идэвхжүүлэх газруудад чиглүүлэх // J. Clin. Хөрөнгө оруулах. 2003. V. 111. P. 1875-1885.
31. Thiel S, Petersen S.V., Vorup-Jensen T. et al. C1q ба маннан холбогч лектин (MBL) C1r, C1s, MBL-тэй холбоотой серин протеаза 1 ба 2, MBL-тэй холбоотой уураг MAp19 // J. Immunol-ийн харилцан үйлчлэл. 2000. V. 165. P. 878-887.
32. Windbichler M., Echtenacher B., Hehlgans T. et al. Туршилтын септик перитонитийн үед нянгийн эсрэг дархлаа хамгаалахад комплементийг идэвхжүүлэх лектины замын оролцоо // Халдвар ба дархлаа. 2004. V. 72. No 9. P. 5247-5252.
Зохицуулалтгүйгээр механизмуудолон үе шаттайгаар ажилладаг, нэмэлт систем нь үр дүнгүй байх болно; Түүний бүрэлдэхүүн хэсгүүдийн хязгааргүй хэрэглээ нь биеийн эс, эд эсэд ноцтой, үхэлд хүргэж болзошгүй. Эхний шатанд C1 дарангуйлагч нь Clr ба Cls-ийн ферментийн идэвхийг хааж, улмаар C4 ба C2-ийн задралыг саатуулдаг. Идэвхжүүлсэн C2 нь богино хугацаанд үйлчилдэг бөгөөд харьцангуй тогтворгүй байдал нь C42 болон C423-ийн ашиглалтын хугацааг хязгаарладаг. Альтернатив замын C3- идэвхжүүлэгч фермент C3bBb нь мөн богино хагас задралын хугацаатай байдаг ч пропердиныг ферментийн цогцолбортой холбосноор цогцолборын ашиглалтын хугацааг уртасгадаг.
AT ийлдэсанафилатоксины идэвхгүйжүүлэгч байдаг - фермент нь N-терминал аргининыг C4a, C3a, C5a-аас салгаж, улмаар тэдний биологийн идэвхийг эрс бууруулдаг. I хүчин зүйл нь C4b ба C3b-г идэвхгүй болгодог, H хүчин зүйл нь I хүчин зүйлээр C3b-ийн идэвхгүйжилтийг хурдасгадаг ба ижил төстэй хүчин зүйл болох C4-ийг холбогч уураг (C4-bp) нь C4b-ийн задралыг I хүчин зүйлээр хурдасгадаг. задрал (FUR) - C3-ийг устгадаг. ба эдгээр мембран дээр үүсдэг C5-конвертазын цогцолборууд.
Бусад эсийн мембраны бүрэлдэхүүн хэсгүүд- холбогдох уураг (тэдгээрийн дотор CD59 нь хамгийн их судлагдсан байдаг) - C8 эсвэл C8 ба C9-ийг холбож чаддаг бөгөөд энэ нь мембраны довтолгооны цогцолборыг (C5b6789) оруулахаас сэргийлдэг. Цусны сийвэнгийн зарим уураг (үүнээс хамгийн их судлагдсан уураг S ба кластерин) нь C5b67 цогцолборыг эсийн мембрантай холбох, C8 эсвэл C9-тэй холбогдох (өөрөөр хэлбэл, мембраны бүрэн довтолгооны цогцолбор үүсэх) эсвэл бусад аргаар блоклодог. энэ цогцолбор үүсэх, нэгдэхээс урьдчилан сэргийлэх.
Комплементийн хамгаалалтын үүрэг
Саармагжуулах вирусуудС1 ба С4 нь эсрэгбиемүүдээр нэмэгдэж, сонгодог буюу альтернатив замын дагуу үүсдэг C3b тогтсон үед улам бүр нэмэгддэг. Тиймээс эсрэгбиеийн тоо бага хэвээр байгаа вирусын халдварын эхний үе шатанд комплемент онцгой ач холбогдолтой юм. Эсрэг биетүүд болон комплементууд нь электрон микроскопоор харагдахуйц ердийн нэмэлт "нүх" үүсгэснээр наад зах нь зарим вирусын халдварыг хязгаарладаг. Clq-ийн рецептортой харилцан үйлчлэлцэх нь зорилтот хэсгийг опсонжуулж, өөрөөр хэлбэл түүний фагоцитозыг хөнгөвчилдөг.
C4a, C3a болон C5aнь гистамин болон бусад зуучлагчдыг ялгаруулж эхэлдэг шигүү мөхлөгт эсүүдээр бэхлэгддэг бөгөөд энэ нь судас тэлэх, хаван үүсэх, үрэвслийн шинж чанартай гипереми үүсэхэд хүргэдэг. C5a-ийн нөлөөн дор моноцитууд нь TNF болон IL-1-ийг ялгаруулж, үрэвслийн хариу урвалыг сайжруулдаг. C5a нь C3b эсвэл түүний задралын бүтээгдэхүүн iC3b-ээр опсонжуулсан бичил биетнийг фагоцитжуулах чадвартай нейтрофил, моноцит, эозинофилийн гол химотактик хүчин зүйл юм. Цаашид эстэй холбогдсон C3b-ийг идэвхгүйжүүлж, C3d-ийн харагдах байдалд хүргэж, түүнийг опсонжуулах үйл ажиллагаанаас нь салгадаг боловч В-лимфоцитуудтай холбогдох чадвар нь хадгалагдан үлддэг. Зорилтот эсэд C3b бэхлэгдсэнээр NK эсүүд эсвэл макрофагуудаар задрах процессыг хөнгөвчилдөг.
C3b холболтуусдаггүй дархлааны цогцолборууд нь тэдгээрийг уусдаг, учир нь C3b нь эсрэгтөрөгч-эсрэгбиеийн цогцолборын торны бүтцийг устгадаг бололтой. Үүний зэрэгцээ энэ цогцолбор нь эритроцитууд дээрх C3b рецептортой (PK1) харилцан үйлчлэлцэх боломжтой болж, энэ цогцолборыг макрофагуудаар шингээж авдаг элэг эсвэл дэлүү рүү шилжүүлдэг. Энэ үзэгдэл нь С1, С4, С2, С3 дутагдалтай хүмүүст сийвэнгийн өвчин (дархлааны цогцолбор өвчин) үүсэхийг хэсэгчлэн тайлбарладаг.