ការបង្កើតអំប្រ៊ីយ៉ុងនៅដំណាក់កាល gastrula ។ ការបង្កើតសរីរាង្គអ័ក្សនៅក្នុងអំប្រ៊ីយ៉ុង chordate
នៅខាងក្រោយ ដំណើរការបំបែក និងបំផ្ទុះដំណើរការអភិវឌ្ឍន៍មួយចំនួនបន្តបន្ទាប់ទៀត ដែលនាំទៅដល់ការកកើតនៃ primitive, anlages បឋមនៃសរីរាង្គនៃអំប្រ៊ីយ៉ុង ពោលគឺរហូតដល់ការលេចចេញនូវស្រទាប់មេជីវិតពីរដំបូង និងបីស្រទាប់ ឬស្រទាប់ពីដំបូង blastoderm តែមួយនៃ blastula (ស្រទាប់មេរោគខាងក្រៅ ឬ ectoderm ស្លឹកកណ្តាល ឬ mesoderm និងស្លឹកខាងក្នុង ឬ endoderm) ។ ដំណាក់កាលនៃការអភិវឌ្ឍន៍ជាមួយនឹងស្រទាប់ដំណុះពីរត្រូវបានគេហៅថា gastrula ។
បន្ទាប់ពីចុងបញ្ចប់នៃរយៈពេល gastrulationការផ្លាស់ប្តូរស្មុគ្រស្មាញកាន់តែច្រើនកើតឡើងដែលដំបូងបណ្តាលឱ្យមានការបង្កើតផ្នែក dorsal នៃអំប្រ៊ីយ៉ុង (notogenesis) ក្រោយមក - ការបង្កើតរាងកាយរបស់វាការបង្កើតបែហោងធ្មែញរាងកាយដែលហៅថា coelom នៅក្នុង mesoderm (coelomatization) បន្ទាប់មក ការបង្កើតខ្សែ dorsal នោះគឺជាអង្កត់ធ្នូ (chordulation) និងការបង្កើត rudiments នៃប្រព័ន្ធសរសៃប្រសាទក្នុងទម្រង់នៃសរសៃប្រសាទ medullary plate និង medullary tube (neurulation) ។ សត្វកាន់តែចាស់ទៅតាមលក្ខណៈ phylogenetically ពោលគឺ ដំណាក់កាលនៃការអភិវឌ្ឍន៍ phylogenetic របស់វាកាន់តែខ្ពស់ ដំណើរការទាំងនេះកាន់តែស្មុគស្មាញ (ទោះបីជាគ្រោងការណ៍មូលដ្ឋានរបស់វានៅតែមិនផ្លាស់ប្តូរ និងអាចមកពីការអភិវឌ្ឍន៍នៃ lancelet) ហើយដំណើរការទាំងនេះកាន់តែត្រួតគ្នាតាមកាលប្បវត្តិ ( តំណពូជ) ។
នៅក្នុងតំបន់នៃបង្គោលលូតលាស់ blastulaនៅក្នុង blastoderm មានទំហំធំជាង blastomeres នៅបង្គោលសត្វ។ តំបន់នៃ blastomeres ធំជាងនេះនៃបង្គោលលូតលាស់ចាប់ផ្តើមចូលបន្តិចម្តង ៗ ចូលទៅក្នុង blastocoel ឆ្ពោះទៅរកបង្គោលសត្វ។
ដោយសារតែនេះ, បែហោងធ្មែញ blastocoelចាប់ផ្តើមថយចុះ ហើយ blastoderm នៃបង្គោលលូតលាស់ចូលទៅជិតស្រទាប់កោសិកានៃ blastoderm នៃបង្គោលសត្វ។ ទីបំផុត ស្រទាប់ទាំងពីរនេះនៅជាប់គ្នា ដែលក្នុងពេលដំណាលគ្នានាំទៅដល់ការបាត់ខ្លួននៃបែហោងធ្មែញ blastocoel ។ ដំណើរការនេះអាចត្រូវបានប្រៀបធៀបដោយលក្ខណៈរូបវិទ្យាសុទ្ធសាធជាមួយនឹងការចូលបន្ទាត់នៃជញ្ជាំងមួយនៃគ្រាប់បាល់កៅស៊ូ perforated នៅខាងក្នុងឆ្ពោះទៅរកជញ្ជាំងផ្ទុយ។ យើងអាចនិយាយបានថា gastrulation នៅក្នុង lancelet កើតឡើងយោងទៅតាមវិធីសាស្រ្តនៃការ intussusception ( invagination ) ។
មិនមានសកម្មភាពទេ។ ingrownតំបន់នៃបង្គោលលូតលាស់ឆ្ពោះទៅរកបង្គោលសត្វ; ដំណើរការនៃការ invagination គឺពិតជាដោយសារតែការលូតលាស់មិនស្មើគ្នានៃ blastula ។ ដោយសារតែកោសិកានៅក្នុងតំបន់នៃបង្គោលសត្វកើនឡើងលឿនជាងកោសិកានៅក្នុងបង្គោលលូតលាស់ blastoderm នៃបង្គោលសត្វដែលលូតលាស់នៅក្នុងទទឹងចាប់ផ្តើមបិទ រួមទាំងតំបន់លូតលាស់យឺតនៃ blastomeres លូតលាស់។
រួមជាមួយនឹង, គ្មានការសង្ស័យការផ្លាស់ប្តូរនៅក្នុងស្ថានភាព colloidal នៃស្រទាប់ផ្ទៃនៃ cytoplasm នៃកោសិកានៅតាមបណ្តោយគែមនៃ blastopore នេះក៏មានសារៈសំខាន់ផងដែរ។
ដូច្នេះជាលទ្ធផល ការឈ្លានពានការបង្កើតរាងដូចពែងមួយលេចឡើង ជញ្ជាំងដែលមានពីរស្រទាប់រួចហើយ ចាប់តាំងពីតំបន់ផ្ទុយគ្នានៃ blastoderm ជាប់គ្នាទៅវិញទៅមកក្នុងអំឡុងពេល invagination ។ នៅផ្នែកខាងក្រោមនៃបែហោងធ្មែញថ្មីដែលបណ្តាលមកពីការ invagination មានស្រទាប់ខាងក្នុងនៃកោសិកាដែលត្រូវគ្នានឹងអតីតបង្គោលលូតលាស់នៃ blastula ។ ផ្ទុយទៅវិញផ្ទៃខាងក្រៅរបស់វាត្រូវបានគ្របដណ្តប់ដោយស្រទាប់នៃ blastoderm ដែលពីមុនមានទីតាំងនៅលើបង្គោលសត្វ។ ដំណាក់កាលនៃការអភិវឌ្ឍន៍នេះដែលត្រូវបានកំណត់ដោយជញ្ជាំងដែលមានស្រទាប់កោសិកាពីរនៅជាប់គ្នាត្រូវបានគេហៅថា gastrula ។ ស្រទាប់ epithelial ខាងក្រៅនៃ gastrula គឺជាស្រទាប់មេរោគខាងក្រៅ ស្រទាប់គឺ ectoderm ស្រទាប់ខាងក្នុងគឺជាស្រទាប់មេរោគខាងក្នុង - endoderm ។
នៅចុងបញ្ចប់នៃរយៈពេលកំទេចនៅក្នុងសត្វពហុកោសិការយៈពេលនៃការបង្កើតស្រទាប់មេរោគចាប់ផ្តើម - gastrulation. វាត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងចលនានៃសម្ភារៈអំប្រ៊ីយ៉ុង។ ទីមួយ gastrula ដំបូងត្រូវបានបង្កើតឡើងដែលមាន 2 ស្រទាប់មេរោគ (ectoderm និង endoderm) បន្ទាប់មក gastrula យឺតនៅពេលដែលស្រទាប់មេរោគទីបី mesoderm ត្រូវបានបង្កើតឡើង។ អំប្រ៊ីយ៉ុងលទ្ធផលត្រូវបានគេហៅថា gastrula ។
ការបង្កើត gastrula ដំបូងអាចកើតឡើងតាមវិធីជាច្រើន: អន្តោប្រវេសន៍, invagination, epiboly ឬ delamination (រូបភាព 5) ។
នៅ អន្តោប្រវេសន៍(ការបណ្តេញចេញ) ផ្នែកនៃកោសិកា blastoderm ពីផ្ទៃនៃអំប្រ៊ីយ៉ុងចូលទៅក្នុង blastocoel ។ ស្រទាប់ខាងក្រៅ (ectoderm) និងស្រទាប់ខាងក្នុង (endoderm) ត្រូវបានបង្កើតឡើង។ blastocoel ត្រូវបានបំពេញដោយកោសិកា។ វិធីសាស្រ្តនៃការបង្កើត gastrula នេះគឺជារឿងធម្មតាឧទាហរណ៍សម្រាប់បែហោងធ្មែញពោះវៀន។
វិចារណញាណ(invagination) ត្រូវបានគេសង្កេតឃើញនៅក្នុងករណីនៃ coeloblastula ។ ក្នុងអំឡុងពេល invagination ផ្នែកជាក់លាក់នៃ blastoderm (បង្គោលលូតលាស់) បត់ចូល និងទៅដល់បង្គោលសត្វ។ អំប្រ៊ីយ៉ុងពីរស្រទាប់ត្រូវបានបង្កើតឡើង - gastrula ។ ស្រទាប់ខាងក្រៅនៃកោសិកាត្រូវបានគេហៅថា ectoderm ស្រទាប់ខាងក្នុងត្រូវបានគេហៅថា endoderm ។ endoderm តម្រង់បែហោងធ្មែញនៃពោះវៀនបឋម - gastrocoel ។ ការបើកដែលបែហោងធ្មែញទំនាក់ទំនងជាមួយបរិយាកាសខាងក្រៅត្រូវបានគេហៅថាមាត់បឋម - ប្លាស្តូប័រ។ យោងតាមជោគវាសនាជាបន្តបន្ទាប់នៃ blastopore សត្វទាំងអស់ត្រូវបានបែងចែកជាពីរក្រុមធំ ៗ គឺបឋមនិង deuterostomes ។ Protostomes រួមមានសត្វដែល blastopore នៅតែជាមាត់អចិន្ត្រៃយ៍ ឬច្បាស់លាស់នៅក្នុងមនុស្សពេញវ័យ (ពពួក Worm, mollusks, arthropods) ។ នៅក្នុងសត្វដទៃទៀត (echinoderms, chordates) blastopore ប្រែទៅជារន្ធគូថឬលូតលាស់ហើយការបើកមាត់លេចឡើងម្តងទៀតនៅចុងផ្នែកខាងមុខនៃរាងកាយរបស់អំប្រ៊ីយ៉ុង។ សត្វបែបនេះត្រូវបានគេហៅថា deuterostomes (chordates) ។
អេបូលី(fouling) គឺជាលក្ខណៈរបស់សត្វដែលកើតចេញពីពង telolecithal ។ ការបង្កើត gastrula គឺដោយសារតែការបែងចែកយ៉ាងឆាប់រហ័សនៃ micromeres ដែលត្រូវបានហ៊ុំព័ទ្ធដោយបង្គោលលូតលាស់។
អង្ករ។ 5. ប្រភេទនៃ gastrulation (Y.P. Antipchuk, 1983)
ខ្ញុំ - ការឈ្លានពាន; II - epiboly, III - អន្តោប្រវេសន៍, IV - delamination ។
Macromeres ស្ថិតនៅក្នុងអំប្រ៊ីយ៉ុង។ មិនមានការបង្កើត blastopore មិនមាន gastrocoel ទេ។ វិធីសាស្រ្តនៃការ gastrulation នេះត្រូវបានគេសង្កេតឃើញនៅក្នុង cyclostomes និង amphibians ។
ការរំលាយចោល(stratification) កើតឡើងនៅក្នុងសារពាង្គកាយដែល blastula ស្រដៀងទៅនឹង morula ។ កោសិកានៃ blastoderm ត្រូវបានបែងចែកទៅជាស្រទាប់ខាងក្រៅ និងខាងក្នុង។ ស្រទាប់ខាងក្រៅបង្កើតជា ectoderm ស្រទាប់ខាងក្នុងបង្កើតជា endoderm ។ វិធីសាស្រ្តនៃការ gastrulation នេះត្រូវបានគេសង្កេតឃើញនៅក្នុងសត្វឆ្អឹងខ្នងជាច្រើននិងសត្វឆ្អឹងខ្នងខ្ពស់។
នៅក្នុងមនុស្ស, gastrulation កើតឡើងជាពីរដំណាក់កាល។ ដំណាក់កាលទី 1 (ថ្ងៃទី 7) កើតឡើងដោយការបំបែកអំប្រ៊ីយ៉ុង។ សន្លឹកពីរត្រូវបានបង្កើតឡើង: ខាងក្រៅគឺជាអេពីប្លាសនិងផ្នែកខាងក្នុងគឺជាអ៊ីប៉ូផ្លាស។ ដំណាក់កាលទីពីរ (14-15 ថ្ងៃ) កើតឡើងជាមួយនឹងការបង្កើត បន្ទះបឋមនិង nodule បឋមដោយចលនា ការធ្វើចំណាកស្រុកនៃកោសិកា។
នៅក្នុងសារពាង្គកាយពហុកោសិកាទាំងអស់ លើកលែងតែអេប៉ុង និង coelenterates ស្រទាប់មេរោគទីបីត្រូវបានបង្កើតឡើង - mesoderm. វាអាចត្រូវបានបង្កើតឡើងជាបួនវិធី (រូបភាពទី 6) ។
Teloblastic - mesoderm ត្រូវបានបង្កើតឡើងដោយកោសិកាធំ ៗ ជាច្រើននៅចុងក្រោយនៃអំប្រ៊ីយ៉ុង - teloblasts ដែលមានទីតាំងនៅចន្លោះ ectoderm និង endoderm ។ ដោយសារតែការ stratification នៃកោសិកា mesoderm បែហោងធ្មែញរាងកាយបន្ទាប់បន្សំត្រូវបានបង្កើតឡើង - ទាំងមូល។ វិធីសាស្រ្តនៃការបង្កើត mesoderm នេះគឺជាលក្ខណៈនៃ protostomes ។
Enterocoelous - mesoderm ត្រូវបានបង្កើតឡើងពីកោសិកានៃ endoderm ក្នុងពេលដំណាលគ្នាជាមួយនឹងការបង្កើត coelom ។ ធម្មតាសម្រាប់ deuterostomes ។
Ectodermal - mesoderm ត្រូវបានបង្កើតឡើងពីផ្នែកមួយនៃកោសិកានៃ ectoderm ដែលមានទីតាំងនៅចន្លោះវានិង endoderm ។ វិធីសាស្រ្តនៃការបង្កើត mesoderm នេះគឺជាលក្ខណៈនៃសត្វល្មូន បក្សី ថនិកសត្វ និងមនុស្ស។
វាចាប់ផ្តើមជាមួយនឹងការលាយបញ្ចូលគ្នានៃ gametes ពីរ (ការបង្កកំណើត) និងការបង្កើត zygote មួយ។ បន្ទាប់ពីការបង្កកំណើត Zygote ចាប់ផ្តើមបែងចែកយ៉ាងខ្លាំង - រយៈពេលនៃការអភិវឌ្ឍន៍អំប្រ៊ីយ៉ុងត្រូវបានគេហៅថា កំទេច . Cleavage - ការបែងចែក mitotic ជាបន្តបន្ទាប់ជាច្រើននៃ zygote ដែលជាលទ្ធផលដែលបរិមាណដ៏ធំនៃ cytoplasm នៃស៊ុតត្រូវបានបែងចែកទៅជាកោសិកាតូចៗជាច្រើនដែលមានស្នូល។ កោសិកាដែលបង្កើតឡើងជាលទ្ធផលនៃការកំទេចត្រូវបានគេហៅថា blastomeres . លក្ខណៈសំខាន់មួយនៃការកំទេច ដែលបែងចែកវាពីការបែងចែកធម្មតា គឺថា blastomeres ដែលទើបបង្កើតថ្មីមិនបង្កើនទំហំទេ។ នេះគឺដោយសារតែការបាត់បង់រយៈពេលលូតលាស់ interphase រវាងការបែងចែក។ ក្នុងករណីនេះរយៈពេលសំយោគនៃ interphase ចាប់ផ្តើមនៅក្នុង telophase នៃវដ្ត mitotic មុន។ នោះគឺមានការកើនឡើងបន្តិចម្តងៗនៃចំនួន blastomeres ជាមួយនឹងបរិមាណសរុបដែលមិនផ្លាស់ប្តូរជាក់ស្តែង។ ប៉ុន្តែដំណើរការនៃការកម្ទេចមិនអាចបន្តដោយមិនកំណត់បានឡើយ។ ដោយសារតែការពិតដែលថាការបែងចែកនីមួយៗក្នុងអំឡុងពេលការបំបែកត្រូវបានអមដោយការថយចុះនៃទំហំកោសិកាវាមានការកើនឡើងបន្តិចម្តង ៗ នៃតម្លៃនៃសមាមាត្រនុយក្លេអ៊ែរ - ស៊ីតូប្លាសស៊ីមដែលត្រូវបានកាត់បន្ថយក្នុងអំឡុងពេលនៃការលូតលាស់ oocyte ។ នៅចំណុចខ្លះ សមាមាត្រនេះឈានដល់តម្លៃធម្មតាសម្រាប់កោសិកា somatic នៃប្រភេទនេះ។
សារៈសំខាន់ជីវសាស្រ្តនៃដំណើរការកំទេច គឺថាដោយសារតែវដ្តនៃការបន្តពូជម្តងហើយម្តងទៀត genotype នៃ zygote ត្រូវបានគុណ ម៉ាស់កោសិកាត្រូវបានបង្គរសម្រាប់ការបំប្លែងបន្ថែមទៀត ពោលគឺឧ។ អំប្រ៊ីយ៉ុង unicellular ក្លាយជាពហុកោសិកា។
ការបែងចែក Blastomere អាចធ្វើសមកាលកម្ម ឬអសមកាល។ ប្រភេទសត្វភាគច្រើនមានការបែងចែកអសមកាលតាំងពីដើមដំបូងនៃការអភិវឌ្ឍន៍របស់វា ហើយខ្លះទៀតវាក្លាយជាដូច្នេះបន្ទាប់ពីការបែងចែកដំបូង។ ធម្មជាតិនៃការកំទេចត្រូវបានកំណត់ដោយរចនាសម្ព័ន្ធនៃស៊ុត និងជាដំបូងនៃការទាំងអស់ដោយបរិមាណនៃ yolk និងរបៀបដែលវាត្រូវបានចែកចាយនៅក្នុង cytoplasm ។ ក្នុងន័យនេះ ស៊ុតមាន់មានពីរប្រភេទសំខាន់ៗ យោងទៅតាមវិធីសាស្ត្រនៃការបុក - កំទេចទាំងស្រុង និងកំទេចដោយផ្នែក។
កំទេចទាំងស្រុង ហៅថាការបុកបែបនេះ នៅពេលដែល cytoplasm នៃស៊ុតត្រូវបានបែងចែកទៅជា blastomeres ត្រូវបានបែងចែកទាំងស្រុង។ នៅក្នុងវេនវាអាចជា ឯកសណ្ឋាននៅពេលដែល blastomeres បានបង្កើតឡើងមានទំហំ និងរូបរាងដូចគ្នា និង មិនស្មើគ្នាប្រសិនបើ blastomeres លទ្ធផលមានទំហំខុសៗគ្នា។ ការកំទេចឯកសណ្ឋានគឺជាលក្ខណៈនៃស៊ុតជាមួយនឹងការផ្គត់ផ្គង់សារធាតុចិញ្ចឹមតិចតួច ដែលត្រូវបានចែកចាយរាបស្មើនៅទូទាំង cytoplasm នៃកោសិកា។ ការកំទេចមិនស្មើគ្នាគឺជាលក្ខណៈនៃស៊ុតដែលមានមាតិកាមធ្យមនៃ yolk ដែលត្រូវបានចែកចាយមិនស្មើគ្នានៅទូទាំង cytoplasm ។
ប្រសិនបើ cytoplasm នៃស៊ុតមិនត្រូវបានបែងចែកទាំងស្រុងទៅជា blastomeres នេះទេ។ ការបំបែក បានហៅ ផ្នែក . ប្រភេទមួយនៃប្រភេទនៃការកំទេចដោយផ្នែកគឺ កំទេចឌីស្កូឌីស នៅពេលដែលការកំទេចកើតឡើងតែនៅក្នុងតំបន់នៃ cytoplasm ដែលមិនមាន yolk ដែលស្នូលស្ថិតនៅ។ តំបន់នៃ cytoplasm ដែលត្រូវបាន cleaved ត្រូវបានគេហៅថា ឌីសមេរោគ . ប្រភេទនៃការកំទេចនេះគឺជារឿងធម្មតាសម្រាប់ស៊ុតជាមួយ ចំនួនធំ yolk (បក្សី) ។
ទោះបីជាការពិតដែលថាការបែងចែកនៅក្នុងតំណាងនៃក្រុមផ្សេងៗនៃសត្វមានលក្ខណៈផ្ទាល់ខ្លួនរបស់វាក៏ដោយក៏វាបញ្ចប់ដោយការបង្កើតរចនាសម្ព័ន្ធស្រដៀងគ្នានៅក្នុងរចនាសម្ព័ន្ធ - blastula . Blastula គឺជាអំប្រ៊ីយ៉ុងស្រទាប់តែមួយ ដែលមានស្រទាប់កោសិកាដែលកំណត់បែហោងធ្មែញ - blastocoel , ឬ បែហោងធ្មែញរាងកាយបឋម . ការបង្កើត blastula ចាប់ផ្តើមនៅដំណាក់កាលដំបូងនៃការកំទេច ដោយសារតែភាពខុសគ្នានៃ blastomeres ។ នេះបង្កើតបែហោងធ្មែញដែលពោរពេញទៅដោយរាវ។
បន្ទាប់ពីការបង្កើតចុងក្រោយនៃ blastula ។ gastrulation ឬការបង្កើតស្រទាប់មេរោគ - ដំណាក់កាលថ្មីនៃការបង្កើតអំប្រ៊ីយ៉ុង។ ដំណាក់កាលនេះ (gastrulation) ត្រូវបានកំណត់លក្ខណៈដោយចលនាដែលពឹងផ្អែកខ្លាំងទាំងកោសិកាបុគ្គល និងកោសិកា។ ការបែងចែកកោសិកាក្នុងអំឡុងពេល gastrulation គឺអវត្តមានជាក់ស្តែងឬតិចតួចណាស់។ ជាលទ្ធផលនៃការ gastrulation ដំបូងស្រទាប់ពីរហើយបន្ទាប់មកអំប្រ៊ីយ៉ុងបីស្រទាប់ត្រូវបានបង្កើតឡើង (ក្នុងករណីភាគច្រើននៅក្នុងសត្វ) - gastrula . ដំបូងបង្អស់ ខាងក្រៅ ( ectoderm ) និងផ្ទៃក្នុង ( endoderm ) ស្រទាប់មេរោគ។ បន្តិចក្រោយមក រវាង ecto- និង endoderm ស្រទាប់មេរោគទីបីលេចឡើង - mesoderm . ភាគច្រើនជាញឹកញាប់ ស្រទាប់មេរោគនេះត្រូវបាន invaginated ចូលទៅក្នុង blastocoel ក្នុងទម្រង់នៃការរីកដុះដាលដូចហោប៉ៅ ដែលបន្ទាប់មកត្រូវបានបិទ។ ក្នុងអំឡុងពេលនៃការបង្កើត mesoderm បែហោងធ្មែញរាងកាយបន្ទាប់បន្សំត្រូវបានបង្កើតឡើងឬ ខូឡូម .
អាស្រ័យលើប្រភេទនៃ blastula ក៏ដូចជាលក្ខណៈនៃចលនាកោសិកា មានវិធីសាស្រ្តសំខាន់ៗជាច្រើនសម្រាប់ការបង្កើតអំប្រ៊ីយ៉ុងពីរស្រទាប់ ឬវិធីសាស្ត្រនៃការបញ្ចេញទឹកកាម៖
- intussusception។ ជាមួយនឹងវិធីសាស្រ្តនេះ ផ្នែកមួយនៃ blastula ចាប់ផ្តើមប៉ោងចូលទៅក្នុង blastocoel (នៅក្នុង lancelet) ។ ក្នុងករណីនេះ blastocoel ត្រូវបានជំនួសស្ទើរតែទាំងស្រុង។ ថង់ស្រទាប់ពីរត្រូវបានបង្កើតឡើង ដែលជញ្ជាំងខាងក្រៅជា ectoderm បឋម ហើយជញ្ជាំងខាងក្នុងគឺជា endoderm បឋមដែលដាក់នៅក្នុងប្រហោងនៃពោះវៀនបឋម ឬ gastrocoel។ រន្ធដែលបែហោងធ្មែញទាក់ទងជាមួយ បរិស្ថានត្រូវបានគេហៅថា blastopore ឬមាត់បឋម។ នៅក្នុងតំណាងនៃក្រុមផ្សេងគ្នានៃសត្វ, ការអភិវឌ្ឍបន្ថែមទៀតនៃ blastopore គឺខុសគ្នា។ ឧទាហរណ៍នៅក្នុង protostomes (ភាគច្រើន invertebrates) វាប្រែទៅជាការបើកមាត់។ នៅក្នុង deuterostomes (echinoderms, chordates) blastopore រីកធំហើយរន្ធគូថជារឿយៗលេចឡើងនៅកន្លែងរបស់វាខណៈពេលដែលការបើកមាត់ផ្ទុះនៅបង្គោលផ្ទុយ (ផ្នែកខាងមុខនៃរាងកាយ);
- អន្តោប្រវេសន៍ - ការបណ្តេញផ្នែកមួយនៃកោសិកា blastula ចូលទៅក្នុងបែហោងធ្មែញនៃ blastocoel (ធម្មតាសម្រាប់សត្វឆ្អឹងកងខ្ពស់) ។ ពួកវាបង្កើតជា endoderm;
- delamination កើតឡើងនៅក្នុងសត្វដែលមាន blastula ដោយគ្មាន blastocoel (បក្សី) ។ ជាមួយនឹងវិធីសាស្រ្តនៃការ gastrulation នេះ, ចលនាកោសិកាគឺអនុវត្តអវត្តមានឬតិចតួច, ចាប់តាំងពីការ stratification កើតឡើង - កោសិកាខាងក្រៅនៃ blastula បង្កើត ectoderm និងកោសិកាខាងក្នុងបង្កើត endoderm;
- epiboly កើតឡើងនៅពេលដែល blastomeres តូចជាងនៃបង្គោលមួយបំបែកលឿនជាងមុន និងក្លាយជា overgrown ជាមួយ blastomeres ធំជាងនៃបង្គោលផ្សេងទៀត ដូច្នេះបង្កើត ectoderm (នៅក្នុង amphibians) ។ ចំណែកឯកោសិកាធំជាងដែលនៅខាងក្នុងបង្កើតឱ្យមានស្រទាប់មេរោគខាងក្នុងគឺអង់ដូឌែម។
វិធីសាស្រ្តដែលបានពិពណ៌នានៃ gastrulation ជាក្បួនមិនកើតឡើងក្នុងទម្រង់ដ៏បរិសុទ្ធរបស់ពួកគេការរួមផ្សំរបស់ពួកគេជាធម្មតាត្រូវបានគេសង្កេតឃើញ (ឧទាហរណ៍ invagination with epiboly in amphibians or delamination with immigration in echinoderms) ។
ស្រទាប់មេរោគ - ទាំងនេះគឺជាស្រទាប់ដាច់ដោយឡែកនៃកោសិកាដែលកាន់កាប់ទីតាំងជាក់លាក់មួយនៅក្នុងអំប្រ៊ីយ៉ុង ហើយផ្តល់ការកើនឡើងដល់សរីរាង្គ និងប្រព័ន្ធសរីរាង្គដែលត្រូវគ្នា។ ផ្កាដុះពន្លកលេចឡើងទាំងដោយសារចលនានៃកោសិកា និងជាលទ្ធផលនៃភាពខុសគ្នានៃកោសិកា blastula ស្រដៀងគ្នា និងស្រដៀងគ្នា។ នៅក្នុងដំណើរការនៃការ gastrulation ស្រទាប់មេរោគកាន់កាប់ទីតាំងមួយដែលត្រូវគ្នាទៅនឹងផែនការរចនាសម្ព័ន្ធនៃសារពាង្គកាយមនុស្សពេញវ័យ។
ភាពខុសគ្នា (ភាពខុសគ្នា) - នេះគឺជាដំណើរការនៃការកើត និងការលូតលាស់នៃភាពខុសគ្នា morphological មុខងាររវាងកោសិកាបុគ្គល និងផ្នែកនៃអំប្រ៊ីយ៉ុង។
សរីរាង្គ
ហៅថាដំណើរការនៃការបង្កើតសរីរាង្គក្នុងការអភិវឌ្ឍន៍អំប្រ៊ីយ៉ុង។ នៅពេលសាងសង់សរីរាង្គណាមួយ ជាលិកាជាច្រើនត្រូវបានប្រើប្រាស់។ ដូច្នេះ ដំណាក់កាលនៃ organogenesis ក៏ជាដំណាក់កាលមួយ។ histogenesis
.
សរីរាង្គអាចបែងចែកជា ២ ដំណាក់កាល៖
- សរសៃប្រសាទ - ស្មុគស្មាញនៃសរីរាង្គអ័ក្សត្រូវបានបង្កើតឡើង (បំពង់សរសៃប្រសាទអង្កត់ធ្នូបំពង់ពោះវៀន) ស្ទើរតែអំប្រ៊ីយ៉ុងទាំងមូលត្រូវបានចូលរួមនៅក្នុងដំណើរការនេះ;
- ការសាងសង់សរីរាង្គដែលនៅសេសសល់ - ផ្នែកផ្សេងៗនៃរាងកាយទទួលយកទម្រង់ធម្មតានិងលក្ខណៈពិសេសនៃអង្គការផ្ទៃក្នុងសមាមាត្រជាក់លាក់ត្រូវបានបង្កើតឡើង។
ការកើតឡើងនៃសរីរាង្គ ដូចដែលបានពិចារណានៅក្នុងទ្រឹស្ដីនៃស្រទាប់មេរោគដោយ Karl Baer កើតឡើងដោយសារតែការផ្លាស់ប្តូរនៃស្រទាប់មេរោគមួយឬមួយផ្សេងទៀត - ecto-, meso- ឬ endoderm ។ សរីរាង្គមួយចំនួនមានប្រភពដើមចម្រុះ ពោលគឺពួកវាត្រូវបានបង្កើតឡើងដោយមានការចូលរួមពីស្រទាប់មេរោគជាច្រើន។ ឧទាហរណ៍ សាច់ដុំនៃបំពង់រំលាយអាហារ គឺជាដេរីវេនៃ mesoderm ហើយស្រទាប់ខាងក្នុងរបស់វាគឺជាដេរីវេនៃ endoderm ។ ទោះជាយ៉ាងណាក៏ដោយ សម្រាប់ភាពសាមញ្ញមួយចំនួន យើងអាចនិយាយបានថាប្រភពដើមនៃសរីរាង្គសំខាន់ៗ និងប្រព័ន្ធរបស់វាត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងស្រទាប់មេរោគមួយចំនួន។
នៅលើឆាក សរសៃប្រសាទ មេរោគត្រូវបានគេហៅថា ណឺរូឡា . ប្រព័ន្ធប្រសាទឆ្អឹងខ្នងត្រូវបានបង្កើតឡើងពី neuroectoderm - សម្ភារៈដែលជាផ្នែកមួយនៃផ្នែក dorsal (dorsal) នៃ ectoderm ។ វាមានទីតាំងនៅខាងលើគែមនៃអង្កត់ធ្នូ។ អន្តរកម្មនៃ rudiments ទាំងនេះមាន សារៈសំខាន់ដ៏អស្ចារ្យនៅទូទាំងការអភិវឌ្ឍន៍។ ទីមួយ មានការបង្រួមនៃស្រទាប់កោសិកានៅក្នុងតំបន់នៃ neuroectoderm ដែលនាំទៅដល់ការបង្កើត បន្ទះសរសៃប្រសាទ . បន្ទាប់មកមានការឡើងក្រាស់នៃគែមនៃបន្ទះសរសៃប្រសាទ, គែមកើនឡើង, បង្កើត ផ្នត់សរសៃប្រសាទ . ដោយសារចលនានៃកោសិកានៅតាមបណ្តោយបន្ទាត់កណ្តាលនៅកណ្តាលចាន ក ចង្អូរសរសៃប្រសាទ ដែលបែងចែកអំប្រ៊ីយ៉ុងទៅជាផ្នែកខាងឆ្វេង និងខាងស្តាំនាពេលអនាគត។ នៅពាក់កណ្តាលបន្ទាត់ បន្ទះសរសៃប្រសាទចាប់ផ្តើមបង្កើត គែមរបស់វាប៉ះ ហើយបន្ទាប់មកបិទ។ ដំណើរការទាំងអស់នេះជាលទ្ធផល បំពង់សរសៃប្រសាទ . រមៀលបិទដំបូងនៅផ្នែកកណ្តាលនៃចង្អូរសរសៃប្រសាទហើយបន្ទាប់មកនៅខាងក្រោយ។ នៅក្នុងផ្នែកក្បាលដែលធំទូលាយជាងផ្នែកផ្សេងទៀតការបិទនៃ rollers កើតឡើងចុងក្រោយ។ នៅ ការអភិវឌ្ឍន៍បន្ថែមទៀតផ្នែកខាងមុខ ពង្រីកបង្កើតជាខួរក្បាល ហើយខួរឆ្អឹងខ្នងត្រូវបានបង្កើតចេញពីផ្នែកផ្សេងទៀតនៃសរសៃប្រសាទ។ ជាលទ្ធផលចានសរសៃប្រសាទត្រូវបានផ្លាស់ប្តូរទៅជាបំពង់សរសៃប្រសាទដែលស្ថិតនៅក្រោម ectoderm ។
ក្នុងអំឡុងពេលនៃសរសៃប្រសាទ មិនមែនកោសិកាសរសៃប្រសាទទាំងអស់សុទ្ធតែជាផ្នែកនៃបំពង់សរសៃប្រសាទនោះទេ។ ចង្កោមនៃកោសិកាទាំងនេះបង្កើតនៅតាមបណ្តោយបំពង់សរសៃប្រសាទ ឬសរសៃប្រសាទ។ ក្រោយមកពួកវាធ្វើចំណាកស្រុកពាសពេញអំប្រ៊ីយ៉ុង ហើយកោសិកានៃប្រសាទប្រសាទ មេឌុលឡា អាដ្រេណាល កោសិកាសារធាតុពណ៌ ជាដើមត្រូវបានបង្កើតឡើងពីពួកវា។
បន្ថែមពីលើសម្ភារៈបំពង់សរសៃប្រសាទ ectoderm អេពីដេមីស និងនិស្សន្ទវត្ថុរបស់វាមានការរីកចម្រើន (រោម រោម ក្រញ៉ាំ និងក្រញ៉ាំ ក្រពេញស្បែក។
ការបង្កើតសរីរាង្គ endodermal និង mesodermal កើតឡើងក្នុងពេលដំណាលគ្នាជាមួយនឹងការបង្កើតបំពង់សរសៃប្រសាទ។
ការបញ្ឈប់នៃ mesoderm និងអង្កត់ធ្នូកើតឡើងស្ទើរតែក្នុងពេលដំណាលគ្នាជាមួយនឹងដំណើរការនៃសរសៃប្រសាទ។ ដំបូង នៅតាមជញ្ជាំងចំហៀងនៃពោះវៀនបឋម ដុំពក endoderm protrudes និងហោប៉ៅ ឬបត់ជាទម្រង់។ ផ្នែកនៃ endoderm រវាងផ្នត់ទាំងនេះចាប់ផ្តើមក្រាស់, sag, fold, ហើយបន្ទាប់មក lace ចេញពីភាគច្រើននៃ endoderm នេះ។ ដូច្នេះលេចឡើង អង្កត់ធ្នូ . ការលេចចេញរាងដូចហោប៉ៅនៃ endoderm ត្រូវបានខ្ចាត់ខ្ចាយចេញពីពោះវៀនបឋម ប្រែទៅជាថង់បិទជិត ដែលត្រូវបានគេហៅថា ថង់ coelomic . ជញ្ជាំងរបស់ពួកគេត្រូវបានបង្កើតឡើងពី mesoderm ហើយបែហោងធ្មែញខាងក្នុងគឺជាបែហោងធ្មែញរាងកាយបន្ទាប់បន្សំ (ឬ coelom) ។
គ្រប់ប្រភេទនៃជាលិកាភ្ជាប់, គ្រោងឆ្អឹង, ស្បែក, សាច់ដុំរលោងនិង striated, ប្រព័ន្ធ lymphatic និង circulatory, ប្រព័ន្ធបន្តពូជអភិវឌ្ឍពី mesoderm . epithelium នៃក្រពះនិងពោះវៀន, កោសិកាថ្លើម, កោសិកាសម្ងាត់នៃលំពែង, កោសិកានៃក្រពេញក្រពះនិងពោះវៀនអភិវឌ្ឍពីសម្ភារៈ។ endoderm . ផ្នែកខាងមុខនៃពោះវៀនអំប្រ៊ីយ៉ុងបង្កើតជា epithelium នៃសួត និងផ្លូវដង្ហើម ដែលលាក់កំបាំងស្រទាប់ខាងមុខ និងកណ្តាលនៃក្រពេញភីតូរីស ប៉ារ៉ាទីរ៉ូអ៊ីត និងក្រពេញទីរ៉ូអ៊ីត។
gastrulation(ពីឡាតាំង gaster - ក្រពះ) - ដំណើរការស្មុគ្រស្មាញនៃការផ្លាស់ប្តូរគីមីនិង morphological ដែលត្រូវបានអមដោយការបន្តពូជកំណើនចលនាដឹកនាំនិងភាពខុសគ្នានៃកោសិកាដែលជាលទ្ធផលនៅក្នុងការបង្កើតស្រទាប់មេរោគ - ប្រភពនៃ rudiments នៃជាលិកានិងសរីរាង្គនិងស្មុគស្មាញ។ នៃសរីរាង្គអ័ក្ស។
នៅដំណាក់កាលនៃការអភិវឌ្ឍនៃសារពាង្គកាយនេះ អំប្រ៊ីយ៉ុងពីរស្រទាប់ត្រូវបានបង្កើតឡើង - gastrula. ក្នុងករណីនេះស្រទាប់មេរោគពីរត្រូវបានបង្កើតឡើង - ectoderm (ខាងក្រៅ) និង endoderm (ខាងក្នុង) ។ gastrula ត្រូវគ្នានៅក្នុងរចនាសម្ព័ន្ធទៅនឹងសត្វពោះវៀនទំនើប។ នៅដំណាក់កាលចុងក្រោយនៃការរលាកក្រពះ ស្រទាប់មេរោគទីបីត្រូវបានបង្កើតឡើង - mesoderm (កណ្តាល) ។
សន្លឹកទាំងនេះជាបន្តបន្ទាប់ផ្តល់នូវការកើនឡើងដល់ rudiments អំប្រ៊ីយ៉ុង ដែលជាលិកា និងសរីរាង្គត្រូវបានបង្កើតឡើង។
មានបួនប្រភេទនៃ gastrulation (រូបភាព 8) ។
អន្តោប្រវេសន៍(ការលុកលុយ) គឺជាទម្រង់ដំបូងបំផុត នៃការរលាកក្រពះ។ ប្រភេទផ្សេងទៀតទាំងអស់នៃ gastrulation គឺមកពីវា។ ក្នុងករណីនេះកោសិកានៃ blastoderm ផ្លាស់ទីទៅ blastocoel ដែលជាកន្លែងដែលពួកគេតាំងលំនៅនៅលើផ្ទៃខាងក្នុងនិងបង្កើតបានជា endoderm និងកោសិកាខាងក្រៅបង្កើត ectoderm . នេះបង្កើតជាប្រហោងក្រពះ ក្រពះពោះវៀន - បែហោងធ្មែញនៃពោះវៀនបឋម (coelenterates) ។
វិចារណញាណ(invagination) - blastoderm នៅបង្គោលលូតលាស់ បត់ចូលខាងក្នុង blastocoel ហើយទៅដល់កោសិកានៃបង្គោលសត្វ។ ក្នុងករណីនេះ gastrocoel ត្រូវបានបង្កើតឡើងដែលទំនាក់ទំនងជាមួយបរិយាកាសខាងក្រៅតាមរយៈរន្ធ - ប្លាស្តូប័រ - មាត់បឋម។
ជាមួយនឹងការអភិវឌ្ឍនៃ blastopore សត្វត្រូវបានបែងចែកជាពីរក្រុម:
protostomes- ប្លាស្តូប័រប្រែទៅជាមាត់ពិត (ដង្កូវ, មូស, ក្លាស);
deuterostomes- មាត់បឋមប្រែទៅជារន្ធគូថនៅផ្នែកខាងក្រោយនៃរាងកាយ ហើយនៅផ្នែកខាងមុខ ការបើកមាត់លេចឡើងម្តងទៀត (brachiopods, echinoderms, chordates) ។
អេបូលី(fouling) - នៅបង្គោលសត្វនៃ blastula កោសិកាបែងចែកលឿនជាងមុនហើយវារទៅលើកោសិកាធំនៃបង្គោលលូតលាស់។ ពីកោសិកានៃបង្គោលសត្វត្រូវបានបង្កើតឡើង ectoderm និងពីកោសិកានៃបង្គោលលូតលាស់ - endoderm ។ ប្រភេទនៃ gastrulation នេះគឺជារឿងធម្មតាសម្រាប់សត្វដែលស៊ុតមានបរិមាណកើនឡើងនៃ yolk (cyclostomes, amphibians) ។
ការរំលាយចោល(stratification) - កោសិកា blastoderm បែងចែក, កោសិកាកូនស្រីផ្លាស់ទីចូលទៅក្នុង blastocoel, បង្កើត endoderm និងកោសិកាខាងក្រៅបង្កើត ectoderm . ក្នុងករណីនេះ blastopore មិនត្រូវបានបង្កើតឡើងទេ ដូច្នេះ gastrocoel មិនទំនាក់ទំនងជាមួយបរិយាកាសខាងក្រៅទេ។ ប្រភេទនៃ gastrulation នេះគឺជាលក្ខណៈនៃសត្វដែលបានបាត់បង់ទុនបម្រុងដ៏ធំនៃ yolk នៅក្នុងស៊ុត (coelenterates, placental ខ្ពស់ជាង) ។
អង្ករ។ 8. ប្រភេទនៃ gastrulation (Y.P. Antipchuk, 1983):
ខ្ញុំ - ការឈ្លានពាន; II - epiboly, III - អន្តោប្រវេសន៍, IV - delamination
នៅដំណាក់កាលចុងក្រោយនៃការរលាកក្រពះ ស្រទាប់មេរោគទីបីចាប់ផ្តើមបង្កើត - mesoderm . វាអាចត្រូវបានបង្កើតឡើងជាបួនវិធីផ្សេងគ្នា (រូបភាពទី 9) ។

អង្ករ។ 9. ប្រភេទនៃការបង្កើត mesoderm (Yu.P. Antipchuk, 1983):
I - teloblastic, II - enterocoelous, III - អន្តរកាល,
IV - ក្រពេញអេកូ; 1 - ectoderm; 2 - endoderm; 3 - mesoderm;
4 - teloblasts; 5 - blastopore; 6 - សម្ភារៈអង្កត់ធ្នូ
Teloblastic- mesoderm ត្រូវបានបង្កើតឡើងដោយកោសិកាធំ ៗ ជាច្រើននៅចុងបញ្ចប់នៃអំប្រ៊ីយ៉ុង - តេឡូប្លាស ស្ថិតនៅចន្លោះ ectoderm និង endoderm ។ ដោយសារតែការ stratification នៃកោសិកា mesoderm បែហោងធ្មែញរាងកាយបន្ទាប់បន្សំត្រូវបានបង្កើតឡើង - ជាទូទៅ . វិធីសាស្រ្តនៃការបង្កើត mesoderm នេះគឺជាលក្ខណៈនៃ protostomes ។
ពោះវៀនធំ- mesoderm ត្រូវបានបង្កើតឡើងពីកោសិកា endoderm ក្នុងពេលដំណាលគ្នាជាមួយនឹងការបង្កើត coelom ។ ធម្មតាសម្រាប់ deuterostomes ។
ស្បែក ectodermal- mesoderm ត្រូវបានបង្កើតឡើងពីផ្នែកមួយនៃកោសិកានៃ ectoderm ដែលស្ថិតនៅចន្លោះវា និង endoderm ។ វិធីសាស្រ្តនៃការបង្កើត mesoderm នេះគឺជាលក្ខណៈនៃសត្វល្មូន បក្សី ថនិកសត្វ និងមនុស្ស។
លាយ(អន្តរកាល) - mesoderm ត្រូវបានបង្កើតឡើងក្នុងពេលដំណាលគ្នាជាមួយ ectoderm និង endoderm អំឡុងពេល gastrulation ។ ធម្មតាសម្រាប់ត្រី cartilaginous, amphibians ។
blastula ដែលគេហៅម្យ៉ាងទៀតថាថង់ដំណុះគឺជាលទ្ធផលចុងក្រោយនៃការបំបែកស៊ុត។ ដំណាក់កាលបន្ទាប់ដែលកាន់កាប់ទីតាំងកម្រិតមធ្យមរវាងការកំទេច និងសរីរាង្គនៅក្នុង embryogenesis គឺ gastrulation ។ អត្ថន័យសំខាន់របស់វាគឺការបង្កើតស្រទាប់មេរោគចំនួនបី៖ endoderm, ectoderm និង mesoderm ។ នៅក្នុងពាក្យផ្សេងទៀតវាគឺជាមួយនឹង gastrulation ដែលភាពខុសគ្នានៃអំប្រ៊ីយ៉ុងនិង morphogenesis នៃសារពាង្គកាយចាប់ផ្តើម។
និយមន័យនៃពាក្យ "gastrulation"
ត្រលប់ទៅឆ្នាំ 1901 ក្រពះត្រូវបានពិពណ៌នាថាជាផ្លូវដែលកោសិកា mesodermal, endodermal និង ectodermal ចូលទៅក្នុងអំប្រ៊ីយ៉ុង។ និយមន័យនេះបង្កប់ន័យវត្តមាននៃចន្លោះបង្កើតសរីរាង្គពិសេសនៅក្នុង blastula ។ ដោយបានយល់ពីការពិពណ៌នាដ៏សាមញ្ញនេះ វាជាការងាយស្រួលក្នុងការបន្តទៅកាន់អ្វីដែលស្មុគស្មាញជាងមុន។ អត្ថន័យទំនើបរយៈពេល Gastrulation គឺជាលំដាប់នៃចលនា morphogenetic ដែលជាលទ្ធផលនៃចលនានៃ rudiments ជាលិកាទៅកាន់កន្លែងដែលមានបំណងសម្រាប់ពួកគេស្របតាម "ផែនការ" នៃអង្គការរបស់សារពាង្គកាយ។ ដំណើរការគឺស្មុគស្មាញ ការផ្លាស់ប្តូរត្រូវបានអមដោយការលូតលាស់ និងការបន្តពូជ ចលនាដឹកនាំ និងភាពខុសគ្នានៃកោសិកា។
ដោយពិចារណាលើការរំលាយអាហារក្នុងន័យទូទៅ វាអាចត្រូវបានកំណត់ថាជាដំណាក់កាលមធ្យមដែលជាកម្មសិទ្ធិរបស់ដំណើរការថាមវន្តតែមួយ ក្នុងអំឡុងពេលដែលផ្នែក blastula ត្រូវបានរៀបចំឡើងវិញ ដែលជួយសម្រួលយ៉ាងខ្លាំងដល់ការផ្លាស់ប្តូរទៅកាន់ដំណើរការនៃសរីរាង្គ។
ចលនាកោសិកា
ប្រសិនបើផ្តល់ឱ្យ លក្ខណៈទូទៅដំណើរការដែលបានពិចារណា បន្ទាប់មកយើងអាចនិយាយបានថា gastrulation គឺជា embolism និង epiboly មួយ។ ពាក្យទាំងពីរនេះឆ្លុះបញ្ចាំងពីចលនា morphogenetic នៃកោសិកា ដែលកើតឡើងនៅគ្រប់ដំណាក់កាលនៃការអភិវឌ្ឍន៍ ontogenetic នៃសារពាង្គកាយមួយ។ ទោះយ៉ាងណាក៏ដោយ ពួកគេត្រូវបានបញ្ចេញឱ្យឃើញច្រើនបំផុតក្នុងអំឡុងពេល gastrulation ។ Epibolism គឺជាដំណើរការនៃការផ្លាស់ប្តូរកោសិកានៅតាមបណ្តោយផ្ទៃនៃអំប្រ៊ីយ៉ុង ហើយ embolism គឺជាចលនារបស់ពួកគេចូលទៅក្នុងវា។
នៅក្នុងអំប្រ៊ីយ៉ុង ប្រភេទសំខាន់ៗខាងក្រោមនៃការបញ្ចេញទឹកកាម ឬចលនាកោសិកាត្រូវបានសម្គាល់៖ ការបំប្លែង ការធ្វើអន្តោប្រវេសន៍ ការជាប់ពាក់ព័ន្ធ ការរំលាយ និងអេបូលី។ ព័ត៌មានលម្អិតបន្ថែមអំពីពួកគេ - នៅពេលក្រោយនៅក្នុងអត្ថបទ។
ចលនានៃសន្លឹកក្រឡា
មិនត្រឹមតែកោសិកាបុគ្គល (ធ្វើចំណាកស្រុកដោយសេរី) ប៉ុណ្ណោះទេ ប៉ុន្តែស្រទាប់កោសិកាទាំងមូលក៏អាចចូលរួមក្នុងដំណើរការនៃក្រពះផងដែរ។ ទិសដៅត្រូវបានកំណត់ដោយអន្តរកម្មថេរនិងឆ្ងាយ។ កម្លាំងដំបូងត្រូវបានរកឃើញដោយ P. Weiss ក្នុងទសវត្សរ៍ទី 20 នៃសតវត្សចុងក្រោយ ហើយជាក់ស្តែងកើតឡើងនៅក្នុង embryogenesis ទីពីរ - កម្រនិងពិសេសកើតឡើងជាមួយនឹងកម្រិតតូចមួយនៃប្រូបាប៊ីលីតេក្នុងអំឡុងពេល morphogenesis ធម្មតា។
ក្នុងអំឡុងពេល gastrulation ការបំបែកកោសិកាមិនកើតឡើងទេ។ ដូចដែលបានរៀបរាប់ខាងលើ ចលនានៃកោសិការចាប់ផ្តើម ហើយជាលទ្ធផល ការបង្កើតអំប្រ៊ីយ៉ុងពីរស្រទាប់ ហៅថា gastrula ។ Endoderm និង ectoderm អាចមើលឃើញយ៉ាងច្បាស់។ នៅក្នុងសារពាង្គកាយពហុកោសិកាទាំងអស់ (ករណីលើកលែងតែមួយគត់គឺ coelenterates) ស្របគ្នាជាមួយនឹងការរលាកក្រពះ ឬភ្លាមៗបន្ទាប់ពីវា ស្រទាប់មេរោគទីបីត្រូវបានបង្កើតឡើងដែលហៅថា mesoderm ។ វាគឺជាបណ្តុំនៃកោសិកាដែលស្ថិតនៅចន្លោះ ectoderm និង endoderm ។ ជាលទ្ធផលអំប្រ៊ីយ៉ុងក្លាយជាបីស្រទាប់។
វិធីសាស្រ្តនៃការ gastrulation ដោយផ្ទាល់អាស្រ័យលើប្រភេទនៃ blastula ។
រលាកក្រពះពោះវៀន

ឈ្មោះនៃវិធីសាស្រ្តនិយាយដោយខ្លួនឯង។ Invagination គឺជាការឈ្លានពាននៃជញ្ជាំងស្រទាប់តែមួយនៃ blastula (balstoderm) ចូលទៅក្នុង blastocoel ។ ឧទាហរណ៍ដំបូងបំផុត និងជាឧទាហរណ៍បំផុតនឹងនៅជាមួយបាល់កៅស៊ូ។ នៅពេលអ្នកចុចវា ផ្នែកនៃសម្ភារៈត្រូវបានចុចចូល។ ការឈ្លានពានអាចត្រូវបាននាំយកទៅជញ្ជាំងឆ្ងាយបំផុតឬធ្វើឱ្យមិនសំខាន់។ ជាលទ្ធផល blastula ត្រូវបានផ្លាស់ប្តូរហើយ gastrula ត្រូវបានទទួលក្នុងទម្រង់ជាថង់ពីរស្រទាប់ជាមួយ archenteron ។ ជញ្ជាំងខាងក្នុងរបស់វាគឺជា endoderm បឋម ហើយជញ្ជាំងខាងក្រៅរបស់វាគឺ ectoderm បឋម។ លទ្ធផល archenteron (ពោះវៀនបឋម) ទំនាក់ទំនងជាមួយបរិយាកាសខាងក្រៅតាមរយៈរន្ធដែលហៅថា blastopore ។ ឈ្មោះទីពីររបស់វាគឺមាត់ចម្បង។ ការអភិវឌ្ឍន៍បន្ថែមទៀតរបស់វាអាស្រ័យលើប្រភេទនៃសារពាង្គកាយ។ នៅក្នុងសត្វជាច្រើន blastopore នៅទីបំផុតវិវត្តទៅជាមាត់យ៉ាងច្បាស់។ ក្នុងន័យនេះ ពួកវាត្រូវបានគេហៅថា protostomes (mollusks, worms, arthropods)។ នៅក្នុង deuterostomes, blastopore ប្រែទៅជាប្រឡាយសរសៃប្រសាទ - ពោះវៀនដែលមានទីតាំងនៅខាងក្រោយអំប្រ៊ីយ៉ុង (នៅក្នុង chordates) ឬចូលទៅក្នុងរន្ធគូថ។
gastrula អន្តោប្រវេសន៍
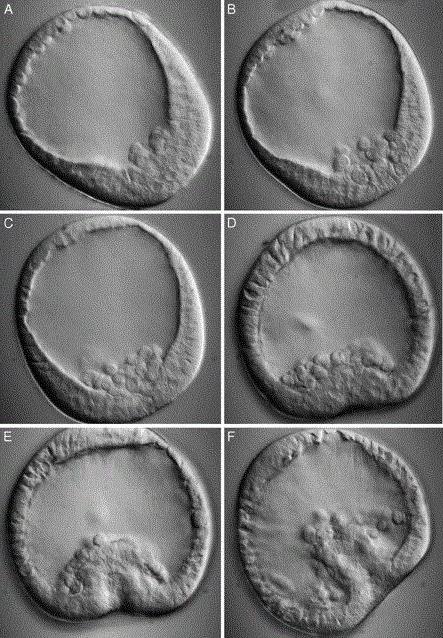
ការធ្វើអន្តោរប្រវេសន៍ gastrulation គឺជាវិធីសាស្រ្តនៃការបង្កើតអំប្រ៊ីយ៉ុងពីរស្រទាប់ដែលជាលក្ខណៈបំផុតនៃ coelenterates ។ gastrula ត្រូវបានបង្កើតឡើងដោយការបណ្តេញចេញយ៉ាងសកម្មនៃផ្នែកមួយនៃកោសិកា blastula ចូលទៅក្នុង blastocoel ។ អន្តោប្រវេសន៍បែបនេះមានលក្ខណៈឯកតោភាគី។ កោសិកាផ្លាស់ទីតែពីបង្គោលលូតលាស់ប៉ុណ្ណោះ។ ក្រោយមកពួកវាបង្កើតជា endoderm ពោលគឺស្រទាប់ខាងក្នុង។ វាគឺនៅក្នុងវិធីនេះដែល gastrulation ត្រូវបានអនុវត្តនៅក្នុង hydroid polyp មួយ jellyfish ។
កោសិកា Blastodermal អាចជ្រាបចូលទៅក្នុង blastocoel មិននៅក្នុងតំបន់ណាមួយឡើយ ប៉ុន្តែនៅលើផ្ទៃទាំងមូលនៃអំប្រ៊ីយ៉ុង។ អន្តោប្រវេសន៍បែបនេះត្រូវបានគេហៅថាពហុប៉ូលប៉ុន្តែវាកម្រណាស់។
នៅក្នុង coelenterates ជាច្រើនដែលត្រូវបានកំណត់ដោយវិធីសាស្រ្តអន្តោរប្រវេសន៍នៃ gastrulation មាន "ការបណ្តេញចេញ" យ៉ាងសកម្មនៃកោសិកា blastula ហើយលទ្ធផល gastrula បាត់បង់ blastocoel ទាំងស្រុង។ ក្នុងករណីនេះលក្ខណៈ blastopore នៃវិធីសាស្ត្រ invagination មុនគឺអវត្តមាន។
ការបំផ្លាញ gastrula

ប្រភេទក្រពះដ៏កម្រនេះត្រូវបានពិពណ៌នាដំបូងដោយ Mechnikov I.I. ហើយវាជារឿងធម្មតាសម្រាប់ពោះវៀន។ ដំណើរការដែលអមដំណើរក្រពះគឺមានលក្ខណៈពិសេសណាស់ ប៉ុន្តែនៅពេលដែលពិចារណាលើករណីធម្មតា គេយល់ឃើញថាសាមញ្ញជាង។ ឧទាហរណ៍ ស៊ុតនៃ scyphomedusa មួយចំនួនមានទីតាំងប្រមូលផ្តុំ និងផ្នែកដែលសម្គាល់យ៉ាងល្អនៃ cytoplasm: ក្រាស់ និង granular (ectoplasm) និង cellular (endoplasm) ។ ពួកវាត្រូវបានកំណត់លក្ខណៈដោយការបែងចែកស៊ីសង្វាក់គ្នា និងឯកសណ្ឋាន៖ 2, 4, 8, 16។ នៅទីបំផុត អំប្រ៊ីយ៉ុងមាន 32 blastomeres ។ ការបែងចែកបន្ថែមទៀតត្រូវបានអនុវត្តស្របទៅនឹងផ្ទៃនៃអំប្រ៊ីយ៉ុង។ ស្រទាប់ខាងក្រៅនៃ blastomeres ត្រូវបានបង្កើតឡើង ដែលរួមមាន ectoplasm និងស្រទាប់ខាងក្នុងមួយផ្នែកនៃ ectoplasm និង endoplasm ។ ម៉្យាងទៀតដំណើរការនៃការបង្កើតអំប្រ៊ីយ៉ុងពហុស្រទាប់ដំណើរការដោយការបំបែកស្រទាប់មួយនៃកោសិកាជាពីរ។ បន្ទាប់មកមានតែ blastomeres ខាងក្នុងប៉ុណ្ណោះដែលត្រូវបានកំទេច ហើយម្តងទៀតស្របទៅនឹងផ្ទៃនៃអំប្រ៊ីយ៉ុង ដែលជាលទ្ធផលនៃការរលាកក្រពះដ៏ចម្លែកបែបនេះ បង្កើតបានជាបាល់មួយ។ វាមានកោសិកាសំប៉ែតចំនួន 64 ដែលបង្កើតជា ectoderm និងកោសិកាប៉ោង 32 ទៀតដែលជាមូលដ្ឋាននៃ endoderm ។
ជំងឺក្រពះពោះវៀន

នៅក្នុងសត្វដែលមានរចនាសម្ព័ន្ធ telolecithal បញ្ចេញសម្លេងនៃស៊ុត (ការផ្លាស់ទីលំនៅនៃ yolk ទៅបង្គោលលូតលាស់) gastrulation កើតឡើងយោងទៅតាមវិធីសាស្រ្តអេពីបូល។ Macromeres គឺជា blastomeres ធំដែលបែងចែកយឺតៗ និងមានបរិមាណច្រើននៃ yolk ។ ពួកវាមិនមានសមត្ថភាពក្នុងការផ្លាស់ទីទេ ដោយភ្ជាប់ជាមួយនេះ មីក្រូម៉េរសកម្មជាងដែលមានទីតាំងនៅលើផ្ទៃក្រឡាតាមព្យញ្ជនៈ "លូន" លើពួកវា។ ជាមួយនឹងការរលាកក្រពះបែបនេះ blastopore គឺអវត្តមាន ហើយ archenteron មិនត្រូវបានបង្កើតឡើងទេ។ មានតែនៅពេលអនាគតនៅពេលដែល macromeres ថយចុះនៅក្នុងទំហំ, តើបែហោងធ្មែញចាប់ផ្តើមបង្កើត, rudiment នៃពោះវៀនបឋម។
ការចូលរួម
Involutionary gastrulation គឺជាដំណើរការមួយដែលមាននៅក្នុង "tucking" ស្រទាប់ខាងក្រៅនៃកោសិកាចូលទៅក្នុងអំប្រ៊ីយ៉ុង។ វារីកធំឡើងនៅតាមផ្ទៃខាងក្នុង។ វិធីសាស្រ្តនៃការ gastrulation នេះគឺជារឿងធម្មតាសម្រាប់សត្វដែលមានស៊ុត mesolecithal - amphibians (amphibians) ។ ចលនានៃកោសិកាជ្រៅឈានមុខគេនៃតំបន់គែមរារាំងការវិវត្តនៃ archenteron ។ វាគឺនៅក្នុងពួកគេនោះ។ កម្លាំងជំរុញការជាប់ពាក់ព័ន្ធ។

វិធីចម្រុះនៃក្រពះ
ដូចដែលអ្នកបានដឹងហើយថា អំប្រ៊ីយ៉ុងហ្សែន គឺជាដំណាក់កាលដំបូងបំផុតក្នុងការអភិវឌ្ឍន៍នៃសារពាង្គកាយនីមួយៗ៖ ពីកំណើតរហូតដល់កំណើត។ Gastrulation គឺជាដំណាក់កាលមួយក្នុងចំណោមដំណាក់កាលរបស់វា ដែលជាដំណាក់កាលទីពីរនៅក្នុងកាលប្បវត្តិបន្ទាប់ពីកំទេច។ វិធីសាស្រ្តរបស់នាងគឺខុសគ្នាខ្លាំងណាស់ដែលពួកគេអាចប្រៀបធៀបជាមួយនឹងកម្រិតខ្ពស់នៃសាមញ្ញ។ ពួកគេម្នាក់ៗទាមទារការសិក្សាលម្អិត និងការវិភាគ។ ទោះយ៉ាងណាក៏ដោយនៅតែមានបន្ទាត់ជាក់លាក់នៃចំនុចប្រសព្វរវាងពួកគេ។ ដូច្នេះ ជាប្រភេទនៃបំរែបំរួលនៃការ invagination ដំណើរការនៃ epiboly អាចត្រូវបានពិចារណា ហើយ delamination មានភាពស្រដៀងគ្នាជាមួយនឹងអន្តោប្រវេសន៍។
ចំណាំថានៅក្នុងសត្វជាច្រើន gastrulation កើតឡើងនៅក្នុងវិធីរួមបញ្ចូលគ្នា។ ក្នុងករណីបែបនេះ អេបូលីស និងការរាតត្បាត ក៏ដូចជាដំណើរការ morphogenetic ផ្សេងទៀតកើតឡើងក្នុងពេលដំណាលគ្នា។ ជាពិសេស នេះជារបៀបដែល gastrulation កើតឡើងនៅក្នុង amphibians ។ ក្នុងន័យនេះ អ្នកនិពន្ធជាច្រើនបានបែងចែកវិធីសាស្រ្តចម្រុះមួយ។
gastrula

តាមព្យញ្ជនៈពីឡាតាំងពាក្យ "gastrula" ត្រូវបានបកប្រែជា "ស្បូនក្រពះ" ។ វាតំណាងឱ្យមេរោគជាក់លាក់នៃសារពាង្គកាយពហុកោសិកា។ លក្ខណៈពិសេសប្លែកនៃ gastrula គឺវត្តមាននៃស្រទាប់មេរោគពីរឬបី។ ដំណើរការនៃការបង្កើតរបស់វាគឺជាដំណាក់កាលនៃ gastrulation ។
ឧបករណ៍សាមញ្ញបំផុតត្រូវបានគេសង្កេតឃើញនៅក្នុងសត្វ។ ពួកវាត្រូវបានកំណត់លក្ខណៈដោយ gastrula រាងពងក្រពើដែលមានស្រទាប់ខាងក្រៅនៃកោសិកា (ectoderm) និងការប្រមូលផ្តុំខាងក្នុងនៃកោសិកា (endoderm) ក៏ដូចជា "ពោះវៀនបឋម" ។ Gastrula ត្រូវបានគេចាត់ទុកថាជាធម្មតា។ urchin សមុទ្រដែលត្រូវបានបង្កើតឡើងដោយ invagination ។ នៅក្នុងមនុស្ស, gastrulation កើតឡើងនៅថ្ងៃទី 8-9 នៃការអភិវឌ្ឍ។ gastrula គឺជាទម្រង់រាងសំប៉ែតដែលបង្កើតចេញពីកោសិកាខាងក្នុង។
តាមក្បួនមួយនៅក្នុងសត្វភាគច្រើននៅដំណាក់កាល gastrula អំប្រ៊ីយ៉ុងមិនអាចរស់នៅដោយសេរីបានទេហើយមានទីតាំងនៅក្នុងស្បូនឬភ្នាសស៊ុត។ ទោះយ៉ាងណាក៏ដោយមានករណីលើកលែង។ ដូច្នេះ larvae នៃបែហោងធ្មែញពោះវៀន planula គឺជា gastrula អណ្តែតដោយសេរី។