Плазмалық мембрананың қызметтері. Плазмалық мембрана Плазманың құрылысы мен қызметі
Плазмалық мембрана ерекше орын алады, өйткені ол жасушаны сыртында шектейді және жасушадан тыс ортамен тікелей байланысты. Оның қалыңдығы шамамен 10 нм және жасушаның ең қалың қабығы. Негізгі компоненттері белоктар (60%-дан астам), липидтер (шамамен 40%) және көмірсулар (шамамен 1%). Барлық басқа жасуша мембраналары сияқты ол EPS арналарында синтезделеді.
Плазмалемма функциялары.
Көлік.
Плазмалық мембрана жартылай өткізгіш, яғни. ол арқылы әртүрлі жылдамдықпен таңдамалы түрде әртүрлі молекулалар өтеді. Заттарды мембрана арқылы тасымалдаудың екі жолы бар: пассивті және белсенді тасымалдау.
Пассивті тасымалдау.Пассивті тасымалдау немесе диффузия энергия енгізуді қажет етпейді. Зарядталмаған молекулалар концентрация градиенті бойымен диффузияланады, зарядталған молекулалардың тасымалдануы сутегі протондарының концентрация градиентіне және электрохимиялық протон градиентіне біріктірілген трансмембраналық потенциалдар айырмасына байланысты. Әдетте, мембрананың ішкі цитоплазмалық беті теріс зарядты алып жүреді, бұл оң зарядталған иондардың жасушаға енуін жеңілдетеді. Диффузияның екі түрі бар: қарапайым және жеңіл.
Қарапайым диффузия кіші бейтарап молекулаларға (H 2 O, CO 2, O 2), сондай-ақ гидрофобты төмен молекулалы органикалық заттарға тән. Бұл молекулалар концентрация градиенті сақталған кезде мембрана белоктарымен өзара әрекеттесусіз мембрананың тесіктері немесе арналары арқылы өте алады.
Жеңілдетілген диффузия гидрофильді молекулаларға тән, олар мембрана арқылы концентрация градиенті бойынша да тасымалданады, бірақ принцип бойынша арнайы мембраналық тасымалдаушы ақуыздардың көмегімен. формалар.
Жеңілдетілген диффузия жоғары селективті, өйткені тасымалдаушы ақуыздың тасымалданатын затқа комплементарлы байланыс орталығы бар, ал тасымалдау белоктың конформациялық өзгерістерімен бірге жүреді. Жеңілдетілген диффузияның мүмкін механизмдерінің бірі келесідей: тасымалдаушы ақуыз (транслоказа) затты байланыстырады, содан кейін мембрананың қарама-қарсы жағына жақындайды, бұл затты босатады, бастапқы конформацияны қабылдайды және қайтадан тасымалдау функциясын орындауға дайын болады. Ақуыздың өзі қалай қозғалатыны туралы аз біледі. Басқа ықтимал тасымалдау механизмі бірнеше тасымалдаушы ақуыздардың қатысуын қамтиды. Бұл жағдайда бастапқы байланысқан қосылыстың өзі бір белоктан екіншісіне өтіп, мембрананың қарама-қарсы жағында болғанша бір немесе басқа белокпен дәйекті түрде байланысады.
Белсенді тасымалдау.Мұндай тасымалдау тасымалдау концентрация градиентіне қарсы жүргізілгенде орын алады. Ол жасушаның энергияны жұмсауын талап етеді. Белсенді тасымалдау жасуша ішінде заттардың жинақталуына қызмет етеді. Энергия көзі көбінесе АТФ болып табылады. Белсенді тасымалдау үшін энергия көзінен басқа мембраналық ақуыздардың қатысуы қажет. Жануарлар жасушаларындағы белсенді тасымалдау жүйелерінің бірі жасуша мембранасы арқылы Na және K + иондарын тасымалдауға жауап береді. Бұл жүйе деп аталады На + - K * -сорғы.Ол K+ иондарының концентрациясы Na * иондарынан жоғары болатын жасушаішілік ортаның құрамын сақтауға жауапты.
Екі ионның концентрация градиенті K+ жасушаның ішкі жағына, ал Na+ сыртына ауысу арқылы сақталады. Екі тасымалдау да концентрация градиентіне қарсы жүреді. Иондардың осылайша таралуы жасушалардағы судың мөлшерін, жүйке жасушалары мен бұлшықет жасушаларының қозғыштығын және қалыпты жасушалардың басқа да қасиеттерін анықтайды. Na + -K + -сорғы - ақуыз - тасымалдау ATPase.Бұл ферменттің молекуласы олигомер болып табылады және мембранадан өтеді. Сорғы жұмысының толық циклі кезінде жасушадан жасушааралық затқа 3 Na+ иондары, ал қарама-қарсы бағытта – 2 К+ иондары ауысады, бұл ретте АТФ молекуласының энергиясы жұмсалады. Кальций иондарын тасымалдауға арналған транспорттық жүйелер (Ca 2+ -ATPase), протондық сорғылар (H + -ATPase) және т.б.
Басқа заттың концентрация градиентінің энергиясы есебінен жүзеге асырылатын заттың мембрана арқылы белсенді өтуі деп аталады. симптом... Бұл жағдайда тасымалдау ATPase екі зат үшін де байланыстыру орындары бар. Антипортзаттың концентрация градиентіне қарсы қозғалысы. Бұл жағдайда басқа зат өзінің концентрация градиенті бойынша қарама-қарсы бағытта қозғалады. Симптом және антипорт (котранспорт) ішектен амин қышқылдарын сіңіру және глюкозаны бірінші реттік несептен қайта сіңіру кезінде, Na +, K + -ATPase арқылы құрылған Na + иондарының концентрация градиентінің энергиясын пайдалана отырып пайда болуы мүмкін.
Тасымалдаудың тағы 2 түрі - эндоцитоз және экзоцитоз.
Эндоцитоз- жасушаның үлкен бөлшектерді ұстауы. Эндоцитоздың бірнеше жолы бар: пиноцитоз және фагоцитоз. Әдетте астында пиноцитозСұйық коллоидты бөлшектерді жасушаның, астында ұстауын түсіну фагоцитоз- денешіктерді ұстау (басқа жасушаларға дейін тығызырақ және үлкенірек бөлшектер). Пино- және фагоцитоздың механизмі әртүрлі.
Жалпы, қатты бөлшектердің немесе сұйық тамшылардың жасушаға сырттан енуін гетерофагия деп атайды. Бұл процесс қарапайымдыларда кеңінен таралған, бірақ ол адамдарда (басқа сүтқоректілерде де) өте маңызды. Гетерофагия организмді қорғауда (сегменттелген нейтрофилдер – гранулоциттер; макрофагоциттер), сүйек тінінің қайта құрылуына (остеокласттар), қалқанша без фолликулалары арқылы тироксиннің түзілуіне, протеиннің және басқа да проксимальды нефрондағы макромолекулалардың реабсорбциясына маңызды рөл атқарады. және басқа процестер.
Пиноцитоз.
Сыртқы молекулалар жасушаға ену үшін алдымен гликокаликс рецепторларымен (мембрананың беткі белоктарымен байланысқан молекулалар жиынтығы) байланысуы керек (сурет).
Мұндай байланысу орнында плазмалемманың астында клатрин ақуызы молекулалары кездеседі. Плазмалемма сырттан бекітілген молекулалармен және цитоплазма астындағы клатринмен бірге басып кіре бастайды. Шығыңқы жер тереңдей түседі, оның шеттері жақындайды, содан кейін жабылады. Нәтижесінде ұсталған молекулаларды тасымалдайтын плазмалеммадан көпіршік бөлінеді. Оның бетіндегі клатрин электронды микросуреттерде біркелкі емес шекараға ұқсайды, сондықтан мұндай көпіршіктер жиектелген деп аталады.
Клатрин везикулалардың жасушаішілік мембраналарға жабысуын болдырмайды. Сондықтан шекаралас көпіршіктер жасушада олардың мазмұнын пайдалану қажет цитоплазманың дәл сол бөліктеріне еркін тасымалдана алады. Осылайша, атап айтқанда, стероидты гормондар ядроға жеткізіледі. Дегенмен, әдетте шекаралас көпіршіктер плазмалеммадан бөлінгеннен кейін көп ұзамай шекарасын төгеді. Клатрин плазмалеммаға ауысады және қайтадан эндоцитоз реакцияларына қатыса алады.
Цитоплазмада жасуша бетінде тұрақты көпіршіктер – эндосомалар көбірек болады. Шектелген көпіршіктер клатринді бөліп, эндосомалармен біріктіріліп, эндосомалардың көлемі мен бетін арттырады. Содан кейін эндосомалардың артық бөлігі жаңа везикула түрінде бөлінеді, онда жасушаға енген заттар болмайды, олар эндосомада қалады. Жаңа везикула жасуша бетіне бағытталып, мембранамен біріктіріледі. Нәтижесінде шекаралас көпіршіктің ыдырауы кезінде пайда болатын плазмалемманың төмендеуі қалпына келеді, сонымен бірге оның рецепторлары да плазмалеммаға қайтады.
Эндосомалар цитоплазмаға батырылады және лизосома мембраналарымен біріктіріледі. Мұндай екінші реттік лизосоманың ішіне кіретін заттар әртүрлі биохимиялық өзгерістерге ұшырайды. Процесс аяқталғаннан кейін лизосома мембранасы фрагменттерге ыдырауы мүмкін, ал лизосоманың ыдырау өнімдері мен мазмұны жасушаішілік метаболикалық реакциялар үшін қолжетімді болады. Мысалы, амин қышқылдары тРНҚ-мен байланысып, рибосомаларға жеткізіледі, ал глюкоза Гольджи кешеніне немесе агранулярлық EPS түтікшелеріне түсе алады.
Эндосомалардың клатриндік шекарасы болмаса да, олардың барлығы лизосомалармен біріктірілмейді. Олардың кейбіреулері бір жасуша бетінен екіншісіне бағытталған (егер жасушалар эпителий қабатын құраса). Онда эндосоманың мембранасы плазмолеммамен қосылып, ішіндегісі сыртқа шығарылады. Осының нәтижесінде заттар жасуша арқылы бір ортадан екінші ортаға өзгеріссіз тасымалданады. Бұл процесс деп аталады трансцитоз... Белок молекулалары, атап айтқанда иммуноглобулиндер де трансцитоз арқылы тасымалдана алады.
Фагоцитоз.
Егер үлкен бөлшектің бетінде жасуша рецепторлары тани алатын молекулалық топтар болса, ол байланысады. Ешбір жағдайда бөгде бөлшектердің өздері мұндай топтарға ие бола бермейді. Дегенмен, денеге түсіп, олар әрқашан қанда және жасушадан тыс ортада болатын иммуноглобулиндер (опсониндер) молекулаларымен қоршалған. Иммуноглобулиндер әрқашан фагоцитарлық жасушалармен танылады.
Бөгде бөлшекті жабатын опсониндер фагоцит рецепторларымен байланысқаннан кейін оның беткі кешені белсендіріледі. Актин микрофиламенттері миозинмен әрекеттесе бастайды, жасуша бетінің конфигурациясы өзгереді. Фагоцит цитоплазмасының өсінділері бөлшектің айналасында созылған. Олар бөлшектің бетін жауып, оның үстіне біріктіріледі. Өсімдіктердің сыртқы жапырақтары біріктіріліп, жасуша бетін жабады.
Өсімдіктердің терең парақтары сіңірілген бөлшектің айналасында мембрана құрайды - ол түзіледі фагосома.Фагосома лизосомалармен қосылып, нәтижесінде олардың комплексі - гетеролизосома (гетеросома,немесе фаголизосома).Онда бөлшектің ұсталған компоненттерінің лизисі жүреді. Лизис өнімдерінің бір бөлігі гетеросомадан жойылады және жасушамен пайдаланылады, ал кейбіреулері лизосомалық ферменттердің әсеріне бейім болмауы мүмкін. Бұл қалдықтар қалдық денелерді құрайды.
Потенциалды түрде барлық жасушалар фагоцитозға қабілетті, бірақ денеде бұл бағытта маманданғандар аз. Бұл нейтрофильді лейкоциттер мен макрофагтар.
Экзоцитоз.
Бұл жасушадан заттарды шығару. Біріншіден, ірі молекулалы қосылыстар Гольджи кешенінде тасымалдау көпіршіктері түрінде бөлінеді. Соңғылары микротүтікшелердің қатысуымен жасуша бетіне бағытталған. Көпіршікті мембрана плазмалеммаға енген, ал көпіршіктің мазмұны жасушаның сыртында болады (сурет) Көпіршіктің плазмалеммамен қосылуы ешқандай қосымша сигналдарсыз жүруі мүмкін. Бұл экзоцитоз деп аталады құрылтайшы.Өзінің зат алмасу өнімдерінің көпшілігі жасушадан осылай шығарылады. Бірқатар жасушалар, алайда, арнайы қосылыстарды - дененің басқа бөліктерінде қолданылатын құпияларды синтездеуге арналған. Құпиясы бар тасымалдау везикуласы плазмалық мембранамен қосылуы үшін сыртқы сигналдар қажет. Сонда ғана біріктіру орын алып, құпия ашылады. Бұл экзоцитоз деп аталады реттеледі... Секрецияның бөлінуіне ықпал ететін сигналдық молекулалар деп аталады либериндер (релизинг факторлары),және жоюдың алдын алу – статиндер.
Рецепторлардың қызметі.
Олар негізінен плазмалемманың бетінде орналасқан және олардың лигандтарымен байланысуға қабілетті гликопротеидтермен қамтамасыз етіледі. Лиганд өзінің рецепторына құлыптың кілті сияқты сәйкес келеді. Лигандтың рецептормен байланысуы полипептид конформациясының өзгеруін тудырады. Трансмембраналық ақуыздың мұндай өзгеруімен жасушадан тыс және жасушаішілік орта арасында байланыс орнатылады.
Рецепторлардың түрлері.
Белок-иондық каналдармен байланысқан рецепторлар. Олар иондардың өту арнасын уақытша ашатын немесе жабатын сигналдық молекуламен әрекеттеседі. (Мысалы, нейротрансмиттер ацетилхолиннің рецепторы иондық арнаны құрайтын 5 суббірліктен тұратын ақуыз. Ацетилхолин болмаған кезде арна жабылады, ал бекітілгеннен кейін ол ашылып, натрий иондарының өтуіне мүмкіндік береді).
Каталитикалық рецепторлар. Олар жасушадан тыс бөліктен (рецептордың өзі) және пролинкиназа ферменті қызметін атқаратын жасушаішілік цитоплазмалық бөліктен тұрады (мысалы, өсу гормонының рецепторлары).
G-белоктармен байланысқан рецепторлар. Бұл лигандпен әрекеттесетін рецептордан және мембранамен байланысқан ферментке (аденилатциклаза) немесе иондық арнаға сигнал беретін G-белоктан (гуанозинтрифосфатпен байланысқан реттеуші ақуыз) тұратын трансмембраналық ақуыздар. Нәтижесінде циклдік АМФ немесе кальций иондары белсендіріледі. (Аденилатциклаза жүйесі осылай жұмыс істейді. Мысалы, бауыр жасушаларында инсулин гормонының рецепторы болады. Рецептордың жасушадан жоғары бөлігі инсулинмен байланысады. Бұл жасушаішілік бөліктің - аденилатциклаза ферментінің белсендірілуін тудырады. Ол әртүрлі жасушаішілік процестердің жылдамдығын реттейтін, сол немесе басқа метаболикалық ферменттердің белсендірілуін немесе тежелуін тудыратын АТФ-дан циклдік АМФ синтездейді.
Физикалық факторларды қабылдайтын рецепторлар. Мысалы, родопсиннің фоторецепторлы ақуызы. Жарықты жұтқанда конформациясын өзгертіп, жүйке импульсін қоздырады.
Ол липидті қабаттан тұрады, оның липидтері қатаң бағытталған – липидтердің гидрофобты бөлігі (құйрық) қабаттың ішке қараған, ал гидрофильді бөлігі (бас) сыртқа қарайды. Липидтерден басқа плазмалық мембрананың құрылысына мембраналық белоктардың үш түрі қатысады: перифериялық, интегралдық және жартылай интегралдық.
Мембраналарды зерттеу бағыттарының бірі қазіргі уақытта мембраналарды құрайтын әртүрлі құрылымдық және реттеуші липидтердің де, жеке интегралдық және жартылай интегралдық белоктардың да қасиеттерін егжей-тегжейлі зерттеу болып табылады.
Интегралдық мембраналық ақуыздар
Мембрананың өзін ұйымдастыруда негізгі рөлді интегралды және жартылай интегралды белоктар атқарады, олар глобулярлы құрылымға ие және липидті фазамен гидрофильді - гидрофобты әрекеттесу арқылы байланысады. Интегралдық белоктардың глобулдары мембрананың бүкіл қалыңдығына енеді, ал олардың гидрофобты бөлігі глобулдың ортасында орналасады және липидті фазаның гидрофобты аймағына батырылады.
Жартылай интегралдық мембраналық ақуыздар
Жартылай интегралды белоктарда гидрофобты аминқышқылдары глобулдың бір полюсінде шоғырланған, соған сәйкес шарлар бір (сыртқы немесе ішкі) мембрана бетінен сыртқа шығып, мембранаға жартылай ғана батырылған.
Мембраналық ақуыздардың қызметі
Плазмалық мембрананың интегралды және жартылай интегралдық белоктарына бұрын екі функция берілген: жалпы құрылымдық және арнайы. Осыған сәйкес олардың арасында құрылымдық және қызметтік белоктар ажыратылды. Дегенмен, мембраналардың ақуыздық фракцияларын оқшаулау әдістерін жетілдіру және жеке белоктарды неғұрлым егжей-тегжейлі талдау қазіргі уақытта ешқандай ерекше функцияларды атқармайтын барлық мембраналар үшін әмбебап құрылымдық ақуыздардың жоқтығын көрсетеді. Керісінше, белгілі бір функциялары бар мембраналық ақуыздар өте әртүрлі. Бұл рецепторлық функцияларды орындайтын белоктар, әртүрлі қосылыстардың белсенді және пассивті тасымалдаушылары болып табылатын белоктар және ең соңында көптеген ферменттік жүйелердің бөлігі болып табылатын белоктар. Сайттан алынған материал
Мембраналық ақуыздардың қасиеттері
Барлық осы интегралды және жартылай интегралдық мембраналық белоктардың ортақ қасиеті функционалдық жағынан ғана емес, сонымен қатар химиялық жағынан да ерекшеленеді, олардың іргелі қозғалу қабілеті, сұйық липидті фазадағы мембраналық жазықтықта «қалқымалы». Жоғарыда айтылғандай, кейбір жасушалардың плазмалық мембраналарында мұндай қозғалыстардың болуы тәжірибе жүзінде дәлелденді. Бірақ бұл мембраналық ақуыздарда анықталған қозғалыстың жалғыз түрінен алыс. Бүйірлік ығысудан басқа жеке интегралдық және жартылай интегралдық белоктар мембраналық жазықтықта көлденең және тіпті тік бағытта айнала алады, сонымен қатар молекуланың липидтік фазаға батыру дәрежесін өзгерте алады.
Опсин.Белок түйіршіктерінің осы алуан түрлі және күрделі қозғалыстарының барлығы фоторецепторлық жасушалардың мембраналарына тән опсин протеинінің мысалында әсіресе жақсы суреттелген (3-сурет). Өздеріңіз білетіндей, қараңғыда опсин қос цисс байланысы бар каротиноидты тормен байланысты; сетчатки және опсин кешені родопсинді немесе көрнекі күлгін құрайды. Родопсин молекуласы мембрананың көлденең жазықтығында бүйірлік қозғалысқа және айналуға қабілетті (3-сурет, А). Жарық әсер еткенде тор қабық фотоизомеризациядан өтіп, транс формаға айналады. Бұл жағдайда торлы қабықтың конформациясы өзгереді және ол опсиннен бөлінеді, ол өз кезегінде айналу жазықтығын көлденеңінен вертикальға өзгертеді (сурет 3, В). Мұндай трансформациялардың салдары жүйке импульсінің пайда болуына әкелетін иондар үшін мембрананың өткізгіштігінің өзгеруі болып табылады.
Бір қызығы, жарық энергиясының әсерінен индукцияланған опсин глобулдарының конформациясындағы өзгерістер көздің торлы қабығының жасушаларында болатын жүйке импульсін тудыруға қызмет етіп қана қоймайды, сонымен қатар арнайы күлгін бактерияларда кездесетін қарапайым фотосинтетикалық жүйе болып табылады.
Биологиялық мембраналар жасушаның құрылымдық ұйымының негізін құрайды. Плазмалық мембрана (плазмалемма) тірі жасушаның цитоплазмасын қоршап тұрған мембрана. Мембраналар липидтер мен белоктардан тұрады. Липидтер (негізінен фосфолипидтер) қос қабатты құрайды, онда молекулалардың гидрофобты «құйрықтары» мембрананың ішкі жағына, ал гидрофильділері - оның беттеріне бағытталған. Ақуыз молекулалары мембрананың сыртқы және ішкі бетінде орналасуы мүмкін, олар липидті қабатқа ішінара батып кетуі немесе оның ішіне және ішіне еніп кетуі мүмкін. Батырылған мембрана ақуыздарының көпшілігі ферменттер болып табылады. Бұл плазмалық мембрана құрылымының сұйық-сүйек-мозаикалық моделі. Белок және липид молекулалары қозғалғыш, бұл мембрананың динамизмін қамтамасыз етеді. Сондай-ақ мембраналардың құрамында гликолипидтер және мембрананың сыртқы бетінде орналасқан гликопротеидтер (гликокаликс) түріндегі көмірсулар болады. Әрбір жасушаның мембрана бетіндегі белоктар мен көмірсулардың жиынтығы спецификалық болып табылады және жасуша типінің өзіндік көрсеткіші болып табылады.
Мембраналық функциялар:
- Бөлу. Ол жасушаның ішкі мазмұны мен сыртқы орта арасындағы тосқауылдың қалыптасуынан тұрады.
- Цитоплазма мен сыртқы орта арасындағы зат алмасуды қамтамасыз ету. Жасушаға су, иондар, бейорганикалық және органикалық молекулалар түседі (тасымалдау қызметі). Жасушада түзілген өнімдер (секреторлық қызмет) сыртқы ортаға шығарылады.
- Көлік. Мембрана арқылы тасымалдау әртүрлі жолдармен жүруі мүмкін. Пассивті тасымалдау энергия шығынынсыз, тасымалдаушы белоктардың көмегімен қарапайым диффузия, осмос немесе жеңілдетілген диффузия арқылы жүзеге асырылады. Белсенді тасымалдау тасымалдаушы белоктардың көмегімен жүзеге асырылады және ол энергияны қажет етеді (мысалы, натрий-калий сорғы). Сайттан алынған материал
Эндоцитоз нәтижесінде жасушаға биополимерлердің үлкен молекулалары түседі. Ол фагоцитоз және пиноцитоз болып екіге бөлінеді. Фагоцитоз - жасушаның үлкен бөлшектерді ұстап алуы және сіңіруі. Бұл құбылысты алғаш рет И.И. Мечников. Алдымен заттар плазмалық мембранаға, спецификалық рецепторлық ақуыздарға жабысады, содан кейін қабықша салбырап, ойыс түзеді.
Асқорыту вакуольі пайда болады. Ол жасушаға түскен заттарды қорытады. Адамдар мен жануарларда лейкоциттер фагоцитозға қабілетті. Ақ қан жасушалары бактерияларды және басқа бөлшектерді сіңіреді.
Пиноцитоз – құрамында еріген заттары бар сұйық тамшыларды ұстап алу және сіңіру процесі. Заттар мембраналық ақуыздарға (рецепторларға) жабысады, ал ерітінді тамшысы мембранамен қоршалып, вакуоль түзеді. Пиноцитоз және фагоцитоз АТФ энергиясының жұмсалуымен жүреді.
- Секреторлық. Секреция – жасушада синтезделген заттарды жасушаның сыртқы ортаға шығаруы. Гормондар, полисахаридтер, белоктар, май тамшылары мембранамен шектелген көпіршіктерде қоршалып, плазмалеммаға барады. Қабықшалар біріктіріліп, везикуланың мазмұны жасушаны қоршап тұрған ортаға шығарылады.
- Ұлпадағы жасушалардың қосылуы (бүктелген өсінділерге байланысты).
- Рецептор. Мембранада рецепторлардың көп саны – арнайы белоктар болады, олардың рөлі сигналдарды жасушаның сыртынан ішкі жағына жіберу болып табылады.
1. Кедергі- қоршаған ортамен реттелетін, селективті, пассивті және белсенді зат алмасуды қамтамасыз етеді.
Жасуша мембраналары бар селективті өткізгіштік: глюкоза, амин қышқылдары, май қышқылдары, глицерин және иондар олар арқылы баяу таралады, мембраналардың өзі бұл процесті белсенді түрде реттейді - кейбір заттар арқылы рұқсат етіледі, ал басқалары болмайды.
2. Көлік- мембрана арқылы заттар жасуша ішіне және одан тыс тасымалданады. Мембрана арқылы тасымалдау мыналарды қамтамасыз етеді: қоректік заттардың жеткізілуін, метаболизмнің соңғы өнімдерін жоюды, әртүрлі заттардың секрециясын, иондық градиенттерді құруды, жасушалық ферменттердің жұмысына қажетті жасушадағы тиісті рН және иондық концентрацияны сақтау.
Заттардың жасушаға түсуінің немесе оларды жасушадан сыртқа шығарудың төрт негізгі механизмі бар:
а) пассивті (диффузия, осмос) (энергия шығынын қажет етпейді)
Диффузия
Бір заттың молекулаларының немесе атомдарының басқа бір заттың молекулалары немесе атомдары арасында таралуы, олардың концентрацияларының барлық алып жатқан көлемдегі өздігінен теңестірілуіне әкеледі. Кейбір жағдайларда заттардың біреуінің концентрациясы теңестірілген және олар бір заттың екіншісінде диффузиясы туралы айтады. Бұл жағдайда заттың ауысуы концентрациясы жоғары аймақтан концентрациясы төмен аймаққа (концентрация градиентінің векторы бойымен) жүреді. (2.4-сурет).
Күріш. 2.4. Диффузия процесінің диаграммасы
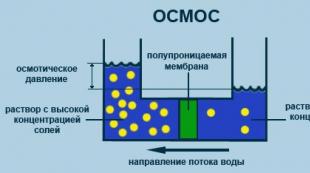
Осмос
Еріткіш молекулаларының жартылай өткізгіш мембранасы арқылы еріген заттың концентрациясы төмен көлемнен еріген заттың жоғары концентрациясына қарай бір жақты диффузия процесі (2.5-сурет).

Күріш. 2.5. Осмос процесінің диаграммасы
б) Белсенді тасымалдау (энергияны тұтынуды қажет етеді)
Натрий-калий сорғысы- концентрация градиенті мен трансмембраналық потенциалдар айырмасын қамтамасыз ететін натрий иондарының (жасушадан) және калий иондарының (жасушаға) белсенді конъюгацияланған трансмембраналық тасымалдау механизмі. Соңғысы жасушалар мен мүшелердің көптеген функцияларына негіз болады: безді жасушалардың секрециясы, бұлшықеттердің жиырылуы, жүйке импульстарының өткізілуі және т.б. (2.6-сурет).

Күріш. 2.6. Калий-натрий сорғысының схемасы
Бірінші кезеңде Na+/K+-ATPase ферменті мембрананың ішкі жағынан үш Na+ ионын бекітеді. Бұл иондар АТФазаның белсенді орталығының конформациясын өзгертеді. Осыдан кейін фермент бір АТФ молекуласын гидролиздеуге қабілетті. Гидролизден кейін бөлінетін энергия тасымалдаушының конформациясын өзгертуге жұмсалады, соның арқасында мембрананың сыртқы жағында үш Na+ иондары және PO 4 3– ион (фосфат) пайда болады. Мұнда Na+ иондары бөлінеді, ал PO 4 3– екі K+ ионымен ауыстырылады. Осыдан кейін фермент өзінің бастапқы конформациясына оралады, ал мембрананың ішкі жағында К+ иондары пайда болады. Мұнда K+ иондары бөлініп, тасымалдаушы қайтадан жұмысқа дайын болады.
Нәтижесінде жасушадан тыс ортада Na+ иондарының жоғары концентрациясы, ал жасуша ішінде К+ жоғары концентрациясы түзіледі. Бұл концентрация айырмашылығы жасушаларда жүйке импульсін өткізу кезінде қолданылады.
в) эндоцитоз (фагоцитоз, пиноцитоз)
Фагоцитоз(жасушамен қоректену) - эукариоттық жасушалар, бактериялар, вирустар, өлі жасушалардың қалдықтары және т.б. сияқты қатты заттарды жасушаның сіңіру процесі. Жұтылған объектінің айналасында үлкен жасушаішілік вакуоль (фагосома) түзіледі. Фагосомалардың мөлшері 250 нм және одан да көп. Фагосоманы бастапқы лизосомамен біріктіру арқылы екінші реттік лизосома түзіледі. Қышқыл ортада гидролиздік ферменттер екінші реттік лизосомадағы макромолекулаларды ыдыратады. Бөлінетін өнімдер (амин қышқылдары, моносахаридтер және басқа да пайдалы заттар) содан кейін лизосомалық мембрана арқылы жасуша цитоплазмасына тасымалданады. Фагоцитоз өте кең таралған. Жоғары ұйымдасқан жануарлар мен адамдарда фагоцитоз процесі қорғаныс қызметін атқарады. Лейкоциттер мен макрофагтардың фагоцитарлық белсенділігі ағзаны патогенді микробтардан және оған түсетін басқа қажетсіз бөлшектерден қорғауда үлкен маңызға ие. Фагоцитозды алғаш рет орыс ғалымы И.И.Мечников сипаттаған (2.7-сурет)
Пиноцитоз(клетка арқылы ішу) – құрамында еритін заттар, соның ішінде ірі молекулалары (белоктар, полисахаридтер және т.б.) бар ортадан сұйық фазаны жасушаның сіңіру процесі. Пиноцитоз кезінде жасуша ішіндегі мембранадан ұсақ көпіршіктер – эндосомалар бөлінеді. Олар фагосомаларға қарағанда кішірек (олардың өлшемі 150 нм-ге дейін) және әдетте ірі бөлшектерді қамтымайды. Эндосома пайда болғаннан кейін оған біріншілік лизосома жақындайды және бұл екі мембраналық көпіршіктер біріктіріледі. Пайда болған органоид екінші реттік лизосома деп аталады. Пиноцитоз процесі барлық эукариоттық жасушаларда үнемі жүреді. (сур. 7)
Рецепторлы эндоцитоз - жасуша қабықшасының жасушаға еніп, қырлы шұңқырлар түзетін белсенді спецификалық процесс. Қапталған шұңқырдың жасушаішілік жағында бейімделгіш белоктар жиынтығы болады. Жасуша бетіндегі арнайы рецепторлармен байланысатын макромолекулалар пиноцитоз арқылы жасушаларға түсетін заттарға қарағанда ішке өте жоғары жылдамдықпен өтеді.

Күріш. 2.7. Эндоцитоз
г) экзоцитоз (теріс фагоцитоз және пиноцитоз)
Жасуша ішілік көпіршіктер (мембраналық көпіршіктер) жасушаның сыртқы мембранасымен біріктірілетін жасушалық процесс. Экзоцитоз кезінде секреторлық көпіршіктердің (экзоцитарлы көпіршіктердің) мазмұны сыртқа шығарылады және олардың мембранасы жасуша қабығымен біріктіріледі. Бұл әдіспен жасушадан барлық дерлік жоғары молекулалық қосылыстар (белоктар, пептидтік гормондар және т.б.) бөлінеді. (2.8-сурет)

Күріш. 2.8. Экзоцитоз схемасы
3. Биопотенциалдардың түзілуі және өткізілуі- мембрананың көмегімен жасушада иондардың тұрақты концентрациясы сақталады: жасуша ішіндегі К+ ионының концентрациясы сыртқа қарағанда әлдеқайда жоғары, ал Na+ концентрациясы әлдеқайда төмен, бұл өте маңызды, өйткені бұл мембранадағы потенциалдар айырмасының сақталуын және жүйке импульсінің пайда болуын қамтамасыз етеді.
4. Механикалық- жасушаның автономиясын, оның жасушаішілік құрылымдарын, сонымен қатар басқа жасушалармен (тіндерде) байланысын қамтамасыз етеді.
5. Энергия- хлоропласттарда фотосинтез және митохондрияларда жасушалық тыныс алу кезінде олардың мембраналарында белоктар да қатысатын энергия тасымалдау жүйелері жұмыс істейді;
6. Рецептор- мембранадағы кейбір белоктар рецепторлар (жасуша белгілі бір сигналдарды қабылдайтын молекулалар) болып табылады.
7. Ферментативті- мембраналық ақуыздар көбінесе ферменттер болып табылады. Мысалы, ішек эпителий жасушаларының плазмалық мембраналарында ас қорыту ферменттері болады.
8. Матрица- мембраналық белоктардың белгілі бір өзара орналасуы мен бағдарлануын, олардың оптималды әрекеттесуін қамтамасыз етеді;
9. Торларды таңбалау- мембранада маркер ретінде әрекет ететін антигендер бар - жасушаны анықтауға мүмкіндік беретін «белгілер». Бұл «антенналар» рөлін атқаратын гликопротеидтер (яғни, тармақталған олигосахаридтердің бүйірлік тізбектері бар белоктар). Маркерлердің көмегімен жасушалар басқа жасушаларды тани алады және олармен бірге әрекет ете алады, мысалы, органдар мен тіндердің пайда болуы кезінде. Ол сондай-ақ иммундық жүйеге бөгде антигендерді тануға мүмкіндік береді.
Жасуша қосындылары
Жасушалық қосындыларға көмірсулар, майлар және белоктар жатады. Бұл заттардың барлығы жасуша цитоплазмасында әртүрлі мөлшердегі және пішіндегі тамшылар мен түйіршіктер түрінде жиналады. Олар жасушада периодты түрде синтезделеді және зат алмасу процесінде қолданылады.
Цитоплазма
Ол плазмалық мембранасы және ядросы жоқ тірі жасушаның (протопласт) бөлігі. Цитоплазманың құрамына: цитоплазмалық матрица, цитоскелет, органеллалар және қосындылар (кейде вакуольдердің қосындылары мен мазмұны цитоплазманың тірі затына жатқызылмайды) кіреді. Сыртқы ортадан плазмалық мембранамен шектелген цитоплазма жасушалардың ішкі жартылай сұйық ортасы болып табылады. Эукариоттық жасушалардың цитоплазмасында ядро және әртүрлі органоидтар орналасқан. Ол сонымен қатар әртүрлі қосындылардан тұрады - жасушалық белсенділіктің өнімдері, вакуольдер, сондай-ақ жасушаның қаңқасын құрайтын ең кішкентай түтікшелер мен жіптер. Цитоплазманың негізгі затының құрамында белоктар басым болады.
Цитоплазмалық функциялар
1) онда негізгі зат алмасу процестері жүреді.
2) ядро мен барлық органоидтарды бір бүтінге біріктіреді, олардың өзара әрекеттесуін қамтамасыз етеді.
3) қозғалғыштық, қозғыштық, зат алмасу және көбею.
Ұтқырлық әртүрлі формада болады:
Жасуша цитоплазмасының жасушаішілік қозғалысы.
Амеба тәрізді қозғалыс. Қозғалыстың бұл түрі цитоплазманың осы немесе басқа тітіркендіргішке қарай немесе одан алыстап псевдоподия түзуінде көрінеді. Қозғалыстың бұл формасы амебаға, қанның лейкоциттеріне, сондай-ақ кейбір ұлпа жасушаларына тән.
Жыпылықтау қозғалысы. Ол ұсақ протоплазмалық өсінділердің – кірпікшелер мен жілікшелердің (кірпікшелер, көп жасушалы жануарлардың эпителий жасушалары, сперматозоидтар және т.б.) соғуы түрінде көрінеді.
Келісімшарттық қозғалыс. Ол цитоплазмада ерекше органоидты миофибрилдердің болуына байланысты қамтамасыз етіледі, олардың қысқаруы немесе ұзаруы жасушаның жиырылуына және босаңсуына ықпал етеді. Жиырылу қабілеті бұлшықет жасушаларында көбірек дамыған.
Тітіркену метаболизм мен энергияны өзгерту арқылы жасушалардың тітіркенуге жауап беру қабілетінде көрінеді.
Цитоскелет
Эукариоттық жасушаның ерекше белгілерінің бірі оның цитоплазмасында микротүтікшелер және белок талшықтары шоғырлары түріндегі қаңқа түзілімдерінің болуы. Сыртқы цитоплазмалық мембранамен және ядролық қабықпен тығыз байланысқан цитоскелет элементтері цитоплазмада күрделі өрімдерді құрайды.
Цитоскелет микротүтікшелерден, микрофиламенттерден және микротрабекулярлық жүйеден түзіледі. Цитоскелет жасушаның пішінін анықтайды, жасуша қозғалысына, жасушаның өзінің бөлінуіне және қозғалуына, органеллалардың жасуша ішілік тасымалдануына қатысады.
Микротүтікшелербарлық эукариоттық жасушаларда болады және диаметрі 30 нм-ден аспайтын, қабырғасының қалыңдығы 5 нм болатын қуыс тармақталмаған цилиндрлер. Олардың ұзындығы бірнеше микрометрге дейін жетуі мүмкін. Оңай бөлшектеңіз және қайта жинаңыз. Микротүтікшелердің қабырғасы негізінен спираль тәріздес бүктелген тубулин ақуызының суббірліктерінен тұрады. (2.09-сурет)
Микротүтікшелердің қызметі:
1) қосалқы функцияны орындайды;
2) бөлінетін шпиндельді құрайды; хромосомалардың жасуша полюстеріне дивергенциясын қамтамасыз ету; жасуша органеллаларының қозғалысына жауапты;
3) жасуша ішілік тасымалдауға, секрецияға, жасуша қабырғасының түзілуіне қатысады;
4) кірпікшелердің, жілікшелердің, базальды денелердің және центриолалардың құрылымдық құрамдас бөлігі болып табылады.
Микрофиламенттербұлшықет актиніне жақын актин протеинінен тұратын диаметрі 6 нм жіптермен ұсынылған. Актин жасушадағы жалпы ақуыздың 10-15% құрайды. Жануарлар жасушаларының көпшілігінде плазмалық мембрананың астында актин жіпшелері мен байланысты белоктардың тығыз желісі түзіледі.
Жасушада актиннен басқа миозин жіпшелері де кездеседі. Алайда олардың саны әлдеқайда аз. Актин мен миозиннің әсерлесуіне байланысты бұлшықеттің жиырылуы жүреді. Микрофиламенттер бүкіл жасушаның немесе оның ішіндегі жеке құрылымдардың қозғалысымен байланысты. Кейбір жағдайларда қозғалыс тек актин жіптерімен, басқаларында миозинмен бірге актинмен қамтамасыз етіледі.
Микрофиламенттердің қызметі
1) механикалық беріктік
2) жасушаның пішінін өзгертуіне және қозғалуына мүмкіндік береді.

Күріш. 2.09. Цитоскелет
Органеллалар (немесе органеллалар)
болып бөлінеді мембраналық емес, бір мембраналық және қос мембраналық.
TO мембраналық емес органоидтарЭукариоттық жасушаларға өздерінің жабық мембранасы жоқ органеллалар жатады, атап айтқанда: рибосомаларжәне тубулиндік микротүтікшелер негізінде құрылған органеллалар - жасуша орталығы (центриолдар)және Қозғалыс органеллалары (жүгі және кірпікшелер).Біржасушалы организмдердің көпшілігінде және жоғары (жер үсті) өсімдіктердің басым көпшілігінде центриолдар болмайды.
TO бір мембраналы органоидтарбайланыстыру: эндоплазмалық ретикулум, Гольджи аппараты, лизосомалар, пероксисомалар, сферосомалар, вакуольдер және басқалары.Барлық бір мембраналы органоидтар бір жасушалық жүйеге өзара байланысқан. Өсімдік жасушаларында арнайы лизосомалар, жануарлар жасушаларында арнайы вакуольдар болады: асқорыту, шығару, жиырылғыш, фагоцитарлық, аутофагоцитарлық және т.б.
TO екі мембраналы органоидтарбайланыстыру митохондриялар мен пластидтер.
Мембраналық емес органоидтар
A) Рибосомалар- барлық организмдердің жасушаларында кездесетін органоидтар. Бұл диаметрі шамамен 20 нм болатын глобулярлы бөлшектермен ұсынылған шағын органеллалар. Рибосомалар мөлшері бірдей емес екі бөлімшеден тұрады – үлкен және кіші. Рибосомаларға белоктар және рибосомалық РНҚ (рРНҚ) жатады. Рибосомалардың екі негізгі түрі бар: эукариоттық (80S) және прокариоттық (70S).
Жасушадағы локализацияға байланысты цитоплазмада белоктарды синтездейтін бос рибосомалар және оған бекінген рибосомалар – ЭПР мембраналарының сыртқы бетімен үлкен суббірліктер арқылы байланысқан, Гольджи кешеніне енетін белоктарды синтездейтін рибосомалар орналасады, содан кейін олар арқылы секрецияланады. жасуша. Белок биосинтезі кезінде рибосомалар комплекстерге – полирибосомаларға (полисомалар) бірігуі мүмкін.
Эукариоттық рибосомалар ядрошықта түзіледі. Алдымен ядролық ДНҚ-да рРНҚ синтезделеді, содан кейін ол цитоплазмадан келетін рибосомалық ақуыздармен жабылып, қажетті мөлшерге дейін ыдырап, рибосоманың суббірліктерін түзеді. Ядрода толық қалыптасқан рибосомалар жоқ. Суббірліктердің тұтас рибосомаға қосылуы цитоплазмада, әдетте белок биосинтезі кезінде жүреді.
Рибосомалар барлық организмдердің жасушаларында кездеседі. Әрқайсысы кіші және үлкен екі бөлшектен тұрады. Рибосомаларға белоктар мен РНҚ кіреді.
Функциялар
ақуыз синтезі.
Синтезделген белоктар алдымен эндоплазмалық тордың арналары мен қуыстарында жинақталады, содан кейін жасушаның органеллалары мен бөліктеріне тасымалданады. Оның мембраналарында орналасқан EPS және рибосомалар белоктардың биосинтезі мен тасымалдануының біртұтас аппаратын білдіреді. (2.10-2.11-сурет).

Күріш. 2.10. Рибосоманың құрылымы

Күріш. 2.11. Рибосоманың құрылымы
В) Жасуша орталығы (центриолдар)
Центриоль - цилиндр (ұзындығы 0,3 мкм және диаметрі 0,1 мкм), оның қабырғасы бір-бірімен белгілі бір аралықта айқаспалы байланыс арқылы қосылған үш біріктірілген микротүтіктердің (9 үштік) тоғыз тобынан құралған. Центриолдар жиі жұпталады, мұнда олар бір-біріне тік бұрышта орналасқан. Егер центриоль кірпікшенің немесе жілікшенің түбінде жатса, онда ол негізгі дене деп аталады.
Барлық дерлік жануарлар жасушаларында жасуша орталығының ортаңғы элементі болып табылатын жұп центриолдар болады.
Бөліну алдында центриолдар қарама-қарсы полюстерге ауытқиды және олардың әрқайсысының жанында аналық центриоль пайда болады. Жасушаның әртүрлі полюстерінде орналасқан центриолдардан бір-біріне қарай өсетін микротүтікшелер түзіледі.
Функциялар
1) еншілес жасушалар арасында генетикалық материалдың біркелкі таралуына ықпал ететін митоздық шпиндель түзеді;
2) цитоскелеттің ұйымдасу орталығы болып табылады. Шпиндель жіптерінің бір бөлігі хромосомаларға бекітіледі.
Центриолдар – цитоплазманың өздігінен көбейетін органоидтары. Олар бұрыннан барлардың қайталануы нәтижесінде пайда болады. Бұл центриолдар бір-бірінен ажыраған кезде пайда болады. Жетілмеген центриолда 9 жалғыз микротүтікше бар; шамасы, әрбір микротүтікше жетілген центриолға тән триплеттерді жинауға арналған матрица болып табылады. (2.12-сурет).
Цетриолдар төменгі сатыдағы өсімдіктердің (балдырлардың) жасушаларында кездеседі.

Күріш. 2.12. Жасуша орталығының центриолдары
Бір мембраналы органоидтар
D) Эндоплазмалық ретикулум (ЭПС)
Цитоплазманың бүкіл ішкі аймағы көптеген ұсақ арналар мен қуыстармен толтырылған, олардың қабырғалары құрылымы жағынан плазмалық мембранаға ұқсас мембраналар болып табылады. Бұл арналар тармақталып, бір-бірімен байланысып, эндоплазмалық ретикулум деп аталатын желіні құрайды. Эндоплазмалық ретикулум құрылымы бойынша гетерогенді. Оның екі түрі белгілі - түйіршіктіжәне тегіс.
Түйіршікті тордың арналары мен қуыстарының мембраналарында көптеген ұсақ дөңгелек денелер бар - рибосомабұл мембраналарға өрескел көрініс береді. Тегіс эндоплазмалық тордың жарғақшалары өз бетінде рибосомаларды тасымалдамайды. EPS көптеген әртүрлі функцияларды орындайды.
Функциялар
Түйіршікті эндоплазмалық тордың негізгі қызметі рибосомаларда жүзеге асырылатын ақуыз синтезіне қатысу. Тегіс эндоплазмалық тордың мембраналарында липидтер мен көмірсулар синтезделеді. Барлық осы синтез өнімдері арналар мен қуыстарда жинақталады, содан кейін жасушаның әртүрлі органеллаларына тасымалданады, онда олар тұтынылады немесе жасушалық қосындылар ретінде цитоплазмада жиналады. EPS жасушаның негізгі органеллаларын байланыстырады (2.13-сурет).

Күріш. 2.13. Эндоплазмалық тордың (ЭПС) немесе ретикулумның құрылымы
E) Гольджи аппараты
Бұл органоидтың құрылымы пішінінің әртүрлілігіне қарамастан, өсімдік және жануарлар организмдерінің жасушаларында ұқсас. Оның көптеген маңызды функциялары бар.
Бір мембраналық органоид. Бұл жиектері ұзартылған тегістелген «цистерналар» дестесі, олармен шағын бір мембраналық көпіршіктер жүйесі (Гольджи көпіршіктері) байланысты. Гольджи көпіршіктері негізінен EPS іргелес жағында және дестелердің шеткі жағында шоғырланған. Олар белоктар мен липидтерді Гольджи аппаратына тасымалдайды деп есептеледі, оның молекулалары цистернадан цистернаға ауыса отырып, химиялық модификацияға ұшырайды.
Бұл заттардың барлығы алдымен жиналып, химиялық күрделене түседі, содан кейін үлкенді-кішілі көпіршіктер түрінде цитоплазмаға түседі де, не жасушаның тіршілігі барысында оның өзінде пайдаланылады, не одан бөлініп, организмде қолданылады. (2.14-2.15-сурет).

Күріш. 2.14. Гольджи аппаратының құрылымы
Функциялар:
Белоктардың, липидтердің, көмірсулардың модификациясы және жинақталуы;
Келетін органикалық заттардың мембраналық көпіршіктерге (везикулаларға) орау;
Лизосомалардың түзілу орны;
Секреторлық қызмет сондықтан Гольджи аппараты секреторлық жасушаларда жақсы дамыған.

Күріш. 2.15. Гольджи кешені
E) Лизосомалар
Олар кішкентай дөңгелек денелер. Лизосома ішінде белоктарды, майларды, көмірсуларды, нуклеин қышқылдарын ыдырататын ферменттер бар. Лизосомалар цитоплазмаға енген тағамдық бөлшекке жақындап, онымен қосылып, бір асқорыту вакуольі пайда болады, оның ішінде лизосомалық ферменттермен қоршалған тағамдық бөлшек болады.
Лизосомалық ферменттер дөрекі EPS-те синтезделеді, Гольджи аппаратына беріледі, онда олар модификацияланады және лизосомалардың мембраналық көпіршіктеріне оралады. Лизосомада 20-дан 60-қа дейін гидролиздік ферменттер болуы мүмкін. Ферменттердің көмегімен заттардың ыдырауы деп аталады лизис.
Біріншілік және екіншілік лизосомаларды ажыратыңыз. Гольджи аппаратынан бүршіктенген бастапқы лизосомалар деп аталады.
Біріншілік лизосомалардың эндоцитарлы вакуольмен қосылуы нәтижесінде түзілетін екіншілік лизосомалар деп аталады. Бұл жағдайда олар жасушаға фагоцитоз немесе пиноцитоз арқылы түскен заттарды қорытады, сондықтан оларды ас қорыту вакуольдері деп атауға болады.
Лизосомалардың қызметі:
1) эндоцитоз кезінде жасуша басып алған заттардың немесе бөлшектердің (бактериялардың, басқа жасушалардың) қорытылуы;
2) аутофагия – жасушаға қажет емес құрылымдардың бұзылуы, мысалы, ескі органоидтарды жаңаларымен ауыстыру немесе жасушаның өзінде түзілетін белоктар мен басқа заттардың қорытылуы;
3) автолиз - жасушаның өлуіне әкелетін өздігінен қорытылуы (кейде бұл процесс патологиялық емес, ағзаның дамуымен немесе кейбір арнайы жасушалардың дифференциациясымен бірге жүреді) (2.16-2.17-сурет).
Мысал: Төбешік бақаға айналғанда, құйрық жасушаларындағы лизосомалар оны қорытады: құйрық жойылып, осы процесте пайда болған заттарды дененің басқа жасушалары сіңіріп, пайдаланады.

Күріш. 2.16. Лизосома түзілуі

Күріш. 2.17. Лизосомалардың қызметі
G) Пероксисомалар
Құрамы бойынша лизосомаларға ұқсас органоидтар, диаметрі 1,5 мкм-ге дейінгі көпіршіктер, құрамында шамамен 50 ферменті бар біртекті матрицасы бар.
Каталаза сутегі асқын тотығы 2H 2 O 2 → 2H 2 O + O 2 ыдырауын тудырады және липидтердің асқын тотығуын болдырмайды.
Пероксисомалар бұрын барлардан бүршіктену арқылы түзіледі, яғни. құрамында ДНҚ жоқтығына қарамастан, өздігінен көбейетін органоидтарға жатады. Оларға ферменттердің түсуіне байланысты өседі, дөрекі ЭПС және гиалоплазмада пероксисомалық ферменттер түзіледі. (2.18-сурет).

Күріш. 2.18. Пероксисома (кристалдық нуклеоидтың ортасында)
H) Вакуольдер
Бір мембраналы органоидтар. Вакуольдер – органикалық және бейорганикалық заттардың сулы ерітінділерімен толтырылған «ыдыстар». Вакуольдердің түзілуіне EPS және Гольджи аппараты қатысады.
Өсімдіктің жас жасушаларында көптеген ұсақ вакуольдар болады, содан кейін олар жасушалар өсіп, дифференциялану кезінде бір-бірімен қосылып, бір үлкен орталық вакуольді құрайды.
Орталық вакуоль жетілген жасуша көлемінің 95% -ын алады, ал ядро мен органеллалар жасуша мембранасына кері итеріледі. Өсімдік вакуолін шектейтін мембрана деп аталады тонопласт.
Өсімдік вакуолін толтыратын сұйықтық жасуша шырыны деп аталады. Жасуша шырынының құрамына суда еритін органикалық және бейорганикалық тұздар, моносахаридтер, дисахаридтер, аминқышқылдары, соңғы немесе улы зат алмасу өнімдері (гликозидтер, алкалоидтар), кейбір пигменттер (антоцианиндер) кіреді.
Органикалық заттардан қант пен белоктар көбірек сақталады. Қанттар – көбінесе ерітінділер түрінде, белоктар ЭПР көпіршіктері және Гольджи аппараты түрінде келеді, одан кейін вакуольдер сусызданып, алейрон дәніне айналады.
Жануарлар жасушаларында екінші реттік лизосомалар тобына жататын және құрамында гидролиздік ферменттері бар ұсақ ас қорыту және аутофагиялық вакуольдер болады. Бір жасушалы жануарларда осморегуляция және экскреция қызметін атқаратын жиырылғыш вакуольдер де болады.
Функциялар
Өсімдіктерде
1) сұйықтықтың жиналуы және тургордың сақталуы;
2) қоректік заттардың және минералды тұздардың қорын жинақтау;
3) гүлдер мен жемістерді бояу және осылайша жемістер мен тұқымдарды тозаңдатқыштар мен таратушыларды тарту.
Жануарларда:
4) ас қорыту вакуольдері – органикалық макромолекулаларды бұзады;
5) жиырылғыш вакуольдер жасушаның осмостық қысымын реттейді және жасушадан қажет емес заттарды шығарады
6) фагоцитоз кезінде антигендердің иммундық жасушалары арқылы фагоцитарлық вакуоль түзіледі
7) автофагоцитарлық вакуольдер фагоцитоз кезінде өз ұлпаларының иммундық жасушаларымен түзіледі
Екі мембраналық органеллалар (митохондриялар мен пластидтер)
Бұл органеллалар жартылай автономды болып табылады, өйткені олардың ДНҚ-сы және ақуыз синтездейтін аппараты бар. Митохондриялар барлық дерлік эукариоттық жасушаларда кездеседі. Пластидтер тек өсімдік жасушаларында болады.
I) Митохондрия
Бұл жасушадағы зат алмасу процестерін энергиямен қамтамасыз ететін органоидтар. Гиалоплазмада митохондриялар әдетте диффузиялық түрде таралады, бірақ мамандандырылған жасушаларда олар энергияны қажет ететін аймақтарда шоғырланған. Мысалы, бұлшықет жасушаларында митохондриялардың көп мөлшері жиырылғыш фибрилдер бойымен, сперматозоидтар бойында, бүйрек түтіктерінің эпителийінде, синапс аймағында және т.б. шоғырланған. Митохондриялардың бұл орналасуы АТФ аз жоғалуын қамтамасыз етеді. оның диффузиясы кезінде.
Сыртқы мембрана митохондрияларды цитоплазмадан бөліп тұрады, өз бетінше жабылады және инвагинациялар түзбейді. Ішкі мембрана митохондриялардың ішкі мазмұнын - матрицаны шектейді. Тән ерекшелігі - көптеген инвагинациялардың - кристалдардың пайда болуы, соның арқасында ішкі мембраналардың ауданы ұлғаяды. Кристалардың саны мен даму дәрежесі ұлпаның функционалдық белсенділігіне байланысты. Митохондриялардың өзіндік генетикалық материалы бар (2.19-сурет).
Митохондриялық ДНҚ – жабық дөңгелек қос тізбекті молекула, адам жасушаларында оның мөлшері 16569 жұп нуклеотидті құрайды, бұл ядрода локализацияланған ДНҚ-дан шамамен 105 есе кіші. Митохондриялардың өздерінің ақуыз синтездейтін жүйесі бар, ал митохондриялық мРНҚ-дан трансляцияланатын белоктардың саны шектеулі. Митохондриялық ДНҚ барлық митохондриялық ақуыздарды кодтай алмайды. Митохондриядағы ақуыздардың көпшілігі ядроның генетикалық бақылауында.

Күріш. 2.19. Митохондриялық құрылым
Митохондриялық функциялар
1) АТФ түзілуі
2) ақуыз синтезі
3) арнайы синтездерге қатысу, мысалы, стероидты гормондардың синтезі (бүйрек үсті бездері)
4) жұмсалған митохондриялар да экскреция өнімдерін, зиянды заттарды жинақтай алады, т.б. басқа жасуша органеллаларының қызметін атқара алады
K) Пластидтер
Пластидтер– Өсімдіктерге ғана тән органоидтар.
Пластидтердің үш түрі бар:
1) хлоропласттар(жасыл пластидтер);
2) хромопластар(сары, қызғылт сары немесе қызыл пластидтер)
3) лейкопластар(түссіз пластидтер).
Әдетте жасушада пластидтердің бір түрі ғана кездеседі.
Хлоропласттар
Бұл органоидтар жапырақтардың жасушаларында және басқа да жасыл өсімдік мүшелерінде, сондай-ақ әртүрлі балдырларда кездеседі. Жоғары сатыдағы өсімдіктерде әдетте бір жасушада бірнеше ондаған хлоропластар болады. Хлоропластардың жасыл түсі олардағы хлорофилл пигментінің құрамына байланысты.
Хлоропласт өсімдік жасушаларының негізгі органоиды болып табылады, онда фотосинтез жүреді, яғни күн сәулесінің энергиясын пайдаланып бейорганикалық заттардан (CO 2 және H 2 O) органикалық заттардың (көмірсулардың) түзілуі. Хлоропластар құрылымы жағынан митохондрияларға ұқсас.
Хлоропластар күрделі құрылымға ие. Олар гиалоплазмадан екі қабықпен – сыртқы және ішкі мембранамен бөлінген. Ішкі мазмұн деп аталады строма... Ішкі мембрана жалпақ көпіршіктер түріндегі күрделі, қатаң реттелген мембраналар жүйесін құрайды. тилакоидтар.
Тилакоидтар үйінділерде жиналады - дәндермонеталардың бағандарына ұқсайды . Граналар бір-бірімен стромальды тилакоидтар арқылы жалғасады, олар арқылы пластид бойымен өтеді (2.20-2.22-сурет).Хлорофилл мен хлоропластар тек жарық әсерінен түзіледі.

Күріш. 2.20. Хлоропластар жарық микроскопында

Күріш. 2.21. Электрондық микроскоптағы хлоропласт құрылымы

Күріш. 2.22. Хлоропластардың схемалық құрылысы
Функциялар
1) фотосинтез(жарық энергиясы есебінен бейорганикалық заттардан органикалық заттардың түзілуі). Бұл процесте хлорофилл негізгі рөл атқарады. Ол жарық энергиясын сіңіреді және оны фотосинтетикалық реакцияларды жүргізуге бағыттайды. Митохондриялар сияқты хлоропласттарда АТФ синтезделеді.
2) аминқышқылдары мен май қышқылдарының синтезіне қатысады;
3) крахмалдың уақытша қорларының қоймасы ретінде қызмет етеді.
Лейкопласттар- күн сәулесінен жасырылған мүшелердің жасушаларында (тамырлар, тамырлар, түйнектер, тұқымдар) кездесетін ұсақ түссіз пластидтер. Олардың құрылымы хлоропласттардың құрылымына ұқсас. (2.23-сурет).
Алайда, хлоропласттардан айырмашылығы, лейкопластардың ішкі мембраналық жүйесі нашар дамыған, өйткені олар қоректік заттардың – крахмалдың, белоктардың және липидтердің синтезі мен жинақталуына қатысады. Жарықта лейкопластар хлоропласттарға айналуы мүмкін.

Күріш. 2.23. Лейкопласт құрылымы
Хромопластар- каротиноидтар тобына жататын пигменттерден туындайтын сарғыш, қызыл және сары түсті пластидтер. Хромопластар көптеген өсімдіктердің жапырақшаларының жасушаларында, жетілген жемістерде, сирек тамыржемістерде, сондай-ақ күзгі жапырақтарда кездеседі. Хромопластарда ішкі мембраналық жүйе, әдетте, жоқ (Cурет 24).

Күріш. 2.24. Хромопласт құрылымы
Хромопластардың маңызы әлі толық зерттелмеген. Олардың көпшілігі ескірген пластидтер. Олар, әдетте, хлоропласттардан дамиды, ал пластидтерде хлорофилл және ішкі мембрана құрылымы бұзылып, каротиноидтар жиналады. Бұл күзде жемістер пісіп, жапырақтары сарғайған кезде болады. Хромопластардың биологиялық маңызы мынада, олар гүлдер мен жемістердің ашық түстерін тудырады, бұл жәндіктерді айқас тозаңдану үшін және басқа жануарларды жемістерді тарату үшін тартады. Лейкопластар хромопластарға да айнала алады.
Пластидтердің қызметі
Қарапайым бейорганикалық қосылыстардан хлорофиллдегі органикалық заттардың синтезі: көмірқышқыл газы мен күн сәулесінің кванттарының қатысуымен су - фотосинтез, Фотосинтездің жеңіл фазасындағы АТФ синтезі
Рибосомаларда белоктардың синтезі (хлоропласттың ішкі мембраналарының арасында ДНҚ, РНҚ және рибосомалар болады, сондықтан хлоропласттарда, сондай-ақ митохондрияларда осы органеллалардың белсенділігіне қажетті ақуыз синтезделеді).
Хромопластардың болуы гүлдердің, жемістердің, күзгі жапырақтардың сары, қызғылт сары және қызыл түсімен түсіндіріледі.
Лейкопласттарда сақтаушы заттар болады (сабақта, тамырда, түйнектерде).
Хлоропластар, хромопластар және лейкопластар жасуша аралық ауысуға қабілетті. Сонымен, күзде жемістер піскенде немесе жапырақтардың түсін өзгерткенде, хлоропласттар хромопластарға айналады, ал лейкопластар хлоропласттарға айналуы мүмкін, мысалы, картоп түйнектері жасыл болған кезде.
Эволюциялық мағынада пластидтердің бастапқы, бастапқы түрі хлоропластар болып табылады, олардан қалған екі типтегі пластидтер пайда болды. Пластидтердің митохондриялармен көптеген ұқсастықтары бар, оларды цитоплазманың басқа компоненттерінен ажыратады. Бұл, ең алдымен, өзінің рибосомалары мен ДНҚ-ның болуына байланысты екі мембрананың қабығы және салыстырмалы генетикалық автономия. Органеллалардың бұл ерекшелігі пластидтер мен митохондриялардың прекурсорлары эволюция процесінде эукариоттық жасушаға айналған және бірте-бірте хлоропластар мен митохондрияларға айналған бактериялар деген пікірге негіз болды. (2.25-сурет).

Күріш. 2.25. Симбиогенез теориясы бойынша митохондриялар мен хлоропластардың түзілуі
Ұяшық- тірі жүйенің элементар бірлігі. Тірі жасушаның белгілі бір қызметті атқаруға жауапты әртүрлі құрылымдары тұтас организмнің мүшелері сияқты органеллалар деп аталады. Жасушадағы спецификалық қызметтер органеллалар, белгілі бір пішіні бар жасушаішілік құрылымдар, мысалы, жасуша ядросы, митохондриялар, т.б.
Жасуша құрылымдары:
Цитоплазма... Жасушаның плазмалық мембрана мен ядроның арасында орналасқан міндетті бөлігі. ЦитозолБұл әртүрлі тұздар мен органикалық заттардың тұтқыр сулы ерітіндісі, ақуыз жіпшелері жүйесі - цитоскелеттер арқылы өтеді. Жасушаның химиялық және физиологиялық процестерінің көпшілігі цитоплазмада өтеді. Құрылысы: Цитозол, цитоскелет. Қызметі: әртүрлі органоидтарды, жасушаның ішкі ортасын қамтиды
Плазмалық мембрана... Жануарлардың, өсімдіктердің әрбір жасушасы қоршаған ортадан немесе басқа жасушалардан плазмалық мембранамен шектелген. Бұл мембрананың қалыңдығы соншалықты кішкентай (шамамен 10 нм), оны тек электронды микроскоппен көруге болады.

Липидтеролар мембранада қос қабат түзеді, ал белоктар оның бүкіл қалыңдығынан өтеді, липидті қабатта әртүрлі тереңдікте батырылады немесе мембрананың сыртқы және ішкі беттерінде орналасады. Барлық басқа органоидтардың мембраналарының құрылысы плазмалық мембранаға ұқсас. Құрылымы: екі қабатты липидтер, белоктар, көмірсулар. Функциялары: шектеу, жасуша пішінін сақтау, зақымданудан қорғау, заттардың түсуі мен шығарылуын реттеу.
Лизосомалар... Лизосомалар - мембраналық органоидтар. Олардың пішіні сопақша және диаметрі 0,5 мкм. Олардың құрамында органикалық заттарды бұзатын ферменттер жиынтығы бар. Лизосома қабықшасы өте күшті және өзінің ферменттерінің жасуша цитоплазмасына енуіне кедергі жасайды, бірақ егер лизосома қандай да бір сыртқы әсерлерден зақымданса, онда бүкіл жасуша немесе оның бір бөлігі жойылады.
Лизосомалар өсімдіктердің, жануарлардың және саңырауқұлақтардың барлық жасушаларында кездеседі.
Әртүрлі органикалық бөлшектерді қорытып, лизосомалар жасушадағы химиялық және энергетикалық процестерге қосымша «шикізат» береді. Аштық кезінде лизосома жасушалары жасушаны өлтірмей кейбір органоидтарды қорытады. Бұл ішінара ас қорыту жасушаны біраз уақыт қоректік заттардың қажетті минимумымен қамтамасыз етеді. Кейде лизосомалар жануарлардың дамуында маңызды рөл атқаратын тұтас жасушалар мен жасушалар тобын қорытады. Мысал ретінде құйрық бақаға айналғанда құйрықты жоғалтуды келтіруге болады. Құрылысы: сопақша пішінді көпіршіктер, сыртында мембрана, ішінде ферменттер. Қызметі: органикалық заттарды бөлу, өлі органоидтарды жою, жұмсалған жасушаларды жою.
Гольджи кешені... Эндоплазмалық ретикулумның қуыстары мен түтікшелерінің люмендеріне түсетін биосинтез өнімдері Гольджи аппаратында шоғырланып, тасымалданады. Бұл органоидтың өлшемі 5-10 мкм.

Құрылым: мембранамен қоршалған қуыстар (везикулалар). Қызметі: органикалық заттардың жинақталуы, буылуы, шығарылуы, лизосомалардың түзілуі
Эндоплазмалық ретикулум... Эндоплазмалық ретикулум - жасуша цитоплазмасында органикалық заттардың синтезі мен тасымалдануына арналған жүйе, ол байланысқан қуыстардың ашық құрылымы болып табылады.
Диаметрі 20 нм шар тәрізді болып келетін жасушаның ең кішкентай органеллалары – эндоплазмалық тордың мембраналарына рибосомалардың көп саны бекітілген. және РНҚ мен ақуыздан тұрады. Белок синтезі рибосомаларда жүреді. Содан кейін жаңадан синтезделген белоктар қуыстар мен түтікшелер жүйесіне еніп, олар бойымен жасуша ішінде қозғалады. Қабықшалардан қуыстар, түтікшелер, түтікшелер, рибосома мембраналарының бетінде. Қызметі: рибосомалардың көмегімен органикалық заттарды синтездеу, заттарды тасымалдау.
Рибосомалар... Рибосомалар эндоплазмалық тордың мембраналарына бекінген немесе цитоплазмада еркін орналасады, олар топ болып орналасады, оларда белоктар синтезделеді. Ақуыз құрамы, рибосомалық РНҚ Қызметі: ақуыз биосинтезін қамтамасыз етеді (ақуыз молекуласының жиналуы).
Митохондрия... Митохондриялар энергетикалық органеллалар болып табылады. Митохондриялардың пішіні әртүрлі, олар қалған, таяқша тәрізді, орташа диаметрі 1 мкм жіп тәрізді болуы мүмкін. және ұзындығы 7 мкм. Митохондриялардың саны жасушаның функционалдық белсенділігіне байланысты және жәндіктердің ұшатын бұлшықеттерінде ондаған мыңға жетуі мүмкін. Митохондриялар сыртынан сыртқы қабықпен шектелген, оның астында көптеген өсінділер – кристалдар түзетін ішкі мембрана орналасқан.

Митохондриялардың ішінде РНҚ, ДНҚ және рибосомалар бар. Оның мембраналарына арнайы ферменттер салынған, олардың көмегімен тағамдық заттардың энергиясы митохондриядағы жасушаның және жалпы организмнің тіршілігіне қажетті АТФ энергиясына айналады.
Мембрана, матрица, өсінділер – кристалдар. Қызметі: АТФ молекуласының синтезі, өз белоктарының, нуклеин қышқылдарының, көмірсулардың, липидтердің синтезі, өз рибосомаларының түзілуі.
Пластидтер... Тек өсімдік жасушасында: лекопластар, хлоропласттар, хромопласттар. Қызметі: резервтік органикалық заттардың жинақталуы, тозаңдандыратын жәндіктердің тартылуы, АТФ және көмірсулардың синтезі. Хлоропласттардың пішіні диаметрі 4–6 мкм болатын диск немесе шар тәрізді. Қос мембраналы - сыртқы және ішкі. Хлоропласттың ішінде бір-бірімен және хлоропласттың ішкі қабығымен байланысқан рибосома ДНҚ және арнайы мембраналық құрылымдар – дәндер болады. Әрбір хлоропластта жарықты жақсы түсіру үшін тізілген 50-ге жуық дәндер бар. Хлорофилл гран мембраналарында орналасқан, оның арқасында күн сәулесінің энергиясы АТФ химиялық энергиясына айналады. АТФ энергиясы хлоропласттарда органикалық қосылыстардың, ең алдымен көмірсулардың синтезіне жұмсалады.
Хромопластар... Хромопластарда кездесетін қызыл және сары пигменттер өсімдіктің әртүрлі бөліктеріне қызыл және сары түс береді. сәбіз, қызанақ жемістері.
Лейкопласттар қоректік зат – крахмалдың жинақталу орны болып табылады. Әсіресе картоп түйнектерінің жасушаларында лейкопласттар көп. Жарықта лейкопластар хлоропласттарға айналуы мүмкін (оның нәтижесінде картоп жасушалары жасылға айналады). Күзде хлоропластар хромопласттарға айналады, ал жасыл жапырақтар мен жемістер сары және қызыл түске айналады.
Жасуша орталығы... Бір-біріне перпендикуляр орналасқан екі цилиндрден, центриолдардан тұрады. Функциялар: шпиндельдерді бөлуге арналған жіп тірегі

Цитоплазмада жасушалық қосындылар пайда болады, содан кейін жасуша тіршілігі процесінде жойылады.

Түйіршіктер түріндегі тығыз қосындыларда резервтік қоректік заттар (крахмал, белоктар, қанттар, майлар) немесе жасушаның қалдықтары бар, оларды әлі жою мүмкін емес. Өсімдік жасушаларының барлық пластидтері резервтік қоректік заттарды синтездеу және жинақтау қабілетіне ие. Өсімдік жасушаларында резервтік қоректік заттардың жиналуы вакуольдерде жүреді.
Дәндер, түйіршіктер, тамшыларФункциялары: органикалық заттар мен энергияны сақтайтын тұрақты емес түзілістер
Негізгі... Екі мембрананың ядролық мембранасы, ядро шырыны, ядрошық. Қызметі: жасушада тұқым қуалайтын ақпаратты сақтау және оның көбеюі, РНҚ синтезі – ақпараттық, тасымалдау, рибосомалық. Ядро қабықшасында споралар болады, олар арқылы ядро мен цитоплазма арасында белсенді зат алмасу жүреді. Ядрода тек берілген жасушаның барлық белгілері мен қасиеттері туралы, онда жүруі керек процестер (мысалы, ақуыз синтезі) туралы ғана емес, сонымен бірге жалпы организмнің ерекшеліктері туралы тұқым қуалайтын ақпарат сақталады. Ақпарат хромосомалардың негізгі бөлігі болып табылатын ДНҚ молекулаларында жазылады. Ядрода ядрошық бар. Ядро құрамында тұқым қуалайтын ақпаратты қамтитын хромосомалардың болуына байланысты жасушаның барлық тіршілік әрекеті мен дамуын басқаратын орталық қызметін атқарады.