Гликолиздің организм үшін маңызы. Гликолиз. Мен қадаммын. Триозафосфат изомерлерінің түзілуі
Гликолиз - көптеген тірі организмдер үшін әртүрлі көмірсулардың катаболизмінің негізгі процесі. Ол фотосинтез жүрмейтін жасушаларда ATP молекулалары түрінде энергияны өндіруге мүмкіндік береді. Анаэробты гликолиз оттегі бар немесе жоқ кезде жүреді.
Процесс ерекшеліктері
Бұл процестің химиялық мәнін көптеген зерттеулер ашыту мен тыныс алудың бастапқы кезеңдерінің ұқсастықтары бар екенін көрсетті. Осы жаңалықтың арқасында ғалымдар тірі дүниедегі бірлікті түсіндірді. Анаэробты гликолиз үшкарбон қышқылдарының циклі өтіп, бос электрондардың тасымалдануы аяқталғаннан кейін жүреді. Пируват митохондрияға еніп, көмірқышқыл газына дейін тотығады. Бұл гексозадан бос энергияның бөлінуіне әкеледі. Гликолиздік реакциялардың үдеткіштері болып табылатын ферменттер цитозолда еритін түрде көптеген жасушаларда болады. Оттегінің қатысуымен болатын гексозаның тотығу процесінің үдеткіштері митохондриялық мембраналарда шоғырланған.
Алты көміртегі атомын екі молекулаға қамтитын көмірсу молекуласын бөлу үшін он белсенді ферменттің болуы қажет. Зерттеушілер оларды таза күйінде бөліп алып, физикалық және химиялық сипаттамаларын зерттей алды.

Процесс қадамдары
Тірі организмдердің жасушаларында гликолиз процесі жүреді. Ол пирожүзім қышқылының түзілуімен бірге жүреді және бірнеше кезеңнен тұрады. Тыныс алуды бұзу үшін глюкозаны белсендіру керек. Ұқсас процесс көміртегі атомы АТФ-мен әрекеттесу кезінде фосфорланған кезде орын алады.
глюкоза + АТФ = глюкоза-6-фосфат + АДФ
Бұл химиялық әрекеттесуді жүзеге асыру үшін магний катиондары мен гексокиназа (фермент) қолданылады. Содан кейін реакция өнімі фруктоза-6-фосфатқа изомерленеді. Катализатор ретінде фосфоглюкоизомераза ферменті қолданылады.
Глюкозаның глюколизі АТФ қатысатын басқа кезеңмен сипатталады. Фосфор қышқылы фруктозаның бірінші көміртегі атомына қосылады. Гликолиздің келесі кезеңдері пайда болған фруктоза-1,6-дифосфаттың триозаға ажырауымен, PHA (3-фосфоглицеральдегид) түзілуімен байланысты.
Гликолиз энергияның бөлінуімен байланысты көп сатылы процесс. Глюкозаның бір молекуласы бөлінгенде екі PHA молекуласы алынады, сондықтан олар қайталанады.
Гликолиз - бұл теңдеумен жинақталған процесс:
C6H12O6 + 2ATP + 2NAD + 2Pn + 4ADP → 2PVK 2NADH + 2H+ + 4ATP + 2ADP

Гликолиздің реттелуі
Гликолиз - тірі организм үшін маңызды процесс. Ол екі функцияны орындауға бағытталған:
- гексоза молекулаларының ыдырауы кезінде АТФ генерациясы;
- келесі синтез процесі үшін құрылыс блоктарын тасымалдау.
Бұл процестің реттелуі тірі жасушаның осы қажеттіліктерін толық қанағаттандыруға бағытталған. Ферменттер катализатор ретінде әрекет ететін реакциялар қайтымсыз.
Реттеуші функцияны жүзеге асыруда фосфофруктокиназа ерекше мәнге ие. Бұл фермент реттеуші функцияны орындайды және процестің жылдамдығын сипаттайды.

Физиологиялық маңызы
Гликолизді глюкоза катаболизмінің әмбебап тәсілі деп санауға болады. Оны про- және эукариоттық организмдер белсенді пайдаланады. Гликолизді катализдейтін ферменттер суда ериді және цитозольде жиналады. Кейбір жануарлар жасушалары мен тіндері гексозаны тек гликолиз арқылы катаболиздей алады. Мысалы, бүйрек арналарының жасушалары мен мидың нейрондары ұқсас қабілеттерге ие.
Май тінінде және бауырда гликолиздің физиологиялық рөлінде кейбір айырмашылықтар бар. Май тінінде және бауырда ас қорыту кезінде бұл процесс майлардың синтезінде қолданылатын субстраттардың көзі болып табылады.
Кейбір өсімдік ұлпалары крахмалды түйнектерде сақтайды. Су өсімдіктері энергияны гликолиз арқылы алады.
Анаэробты жағдайда пируват этанол мен лактатқа айналады. Процесс энергияның көп мөлшерінің бөлінуімен бірге жүреді.
Адипоциттерде гликолиздің маңызды физиологиялық маңызы бар. Оның көмегімен тотығу процесінің орнына липогенез пайда болады, ол тотығу стрессін төмендетеді.

Медициналық маңызы
Анаэробты жағдайда түзілетін лактаттың жиналуымен қанда лактоацидоз дамиды. Бұл қан ортасының реакциясының төмендеуіне әкеледі, бұл жасуша метаболизмінің күрт бұзылуымен бірге жүреді. Ұқсас процесс тіндердің оттегімен қамтамасыз етілуінің бұзылуына байланысты патологиялық процестерде орын алады. Мысалы, миокард инфарктісімен, қан кетумен, өкпе эмболиясымен. Бұл процесс қант диабетімен байланысты, онда аэробты гликолиздің орнына анаэробты процесс жүреді.
Инсулин гликолиздің үдеткіші екенін ескере отырып, 1 типті қант диабетінде гликолиз баяулайды. Сондықтан бұл процесс үшін қолданылатын ферменттерді ынталандыратын дәрілер ауруды емдеу функциясын орындайды.

Қорытынды
Гликолиз – организмдердің толық тіршілігіне қажетті процесс. Қатерлі ісік кезінде глюкозаны тұтыну он есе артады, сондықтан ісік жасушаларының өміршеңдігі гликолизге байланысты.
Бұл процестің ағымының ерекшеліктерін егжей-тегжейлі зерттегеннен кейін ғалымдар гликолизді жасушаларды тамақтандыру үшін ғана емес, сонымен қатар белгілі бір ауруларды емдеу үшін де қолдана алды.
IN анаэробты процесспирожүзім қышқылы сүт қышқылына (лактат) дейін тотықсызданады, сондықтан микробиологияда анаэробты гликолизді сүт қышқылының ашытуы деп атайды. лактат метаболизмге жатады тұйықсодан кейін ештеңеге айналмайды, лактатты пайдаланудың жалғыз жолы - оны қайтадан пируватқа тотықтыру.
Көптеген дене жасушалары глюкозаның анаэробты тотығуына қабілетті. Үшін эритроциттерол энергияның жалғыз көзі. Жасушалар қаңқа бұлшықеттеріглюкозаның оттегісіз ыдырауына байланысты олар күшті, жылдам, қарқынды жұмыстарды орындауға қабілетті, мысалы, спринтинг, күш спортындағы стресс. Физикалық жүктемеден басқа жасушаларда глюкозаның оттегісіз тотығуы гипоксия кезінде күшейеді - әртүрлі типтегі анемия, сағ қан айналымы бұзылыстарысебебіне қарамастан тін.
гликолиз
Глюкозаның анаэробты конверсиясы жүреді цитозолжәне 11 ферментативті реакцияның екі кезеңін қамтиды.
Гликолиздің бірінші сатысы
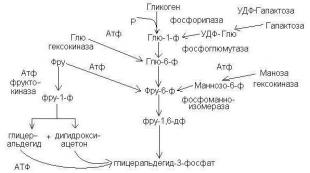
Гликолиздің бірінші сатысы дайындық, мұнда АТФ энергиясы жұмсалады, глюкоза белсендіріледі және одан глюкоза түзіледі. триоза фосфаттары.
Бірінші реакциягликолиз сақинаға қосылмаған 6-шы көміртек атомының фосфорлануына байланысты глюкозаның реактивті қосылысқа айналуына дейін төмендейді. Бұл реакция глюкозаның кез келген конверсиясында гексокиназа катализдейтін бірінші реакция болып табылады.
Екінші реакцияСақинадан тағы бір көміртек атомын оның келесі фосфорлануы (фермент) үшін алып тастау қажет глюкозафосфат изомераза). Нәтижесінде фруктоза-6-фосфат түзіледі.
Үшінші реакция– фермент фосфофруктокиназафруктоза-6-фосфатты симметриялы дерлік фруктоза-1,6-дифосфат молекуласын түзе отырып фосфорлайды. Бұл реакция гликолиз жылдамдығын реттеуде орталық болып табылады.
IN төртінші реакцияфруктоза 1,6-дифосфат екіге бөлінеді фруктоза-1,6-дифосфат-альдолаза екі фосфорланған триоза изомерлері – альдозалардың түзілуімен глицеральдегид(GAF) және кетоздар дигидроксиацетон(DAF).
Бесінші реакциядайындық кезеңі – глицеральдегидфосфат пен дигидроксиацетонфосфаттың қатысуымен бір-біріне ауысуы. триозафосфат изомераза. Реакцияның тепе-теңдігі дигидроксиацетонфосфаттың пайдасына ығысады, оның үлесі 97%, глицеральдегидфосфаттың үлесі 3%. Бұл реакция өзінің қарапайымдылығына қарамастан глюкозаның тағдырын анықтайды:
- жасушада энергияның жетіспеушілігімен және глюкозаның тотығуы белсендірілгенде, дигидроксиацетонфосфат гликолиздің екінші сатысында одан әрі тотығатын глицеральдегидфосфатқа айналады,
- АТФ жеткілікті мөлшерімен, керісінше, глицеральдегидфосфаты дигидроксиацетонфосфатқа изомерленеді, ал соңғысы майлардың синтезіне жіберіледі.
Гликолиздің екінші кезеңі

Гликолиздің екінші кезеңі энергияның бөлінуіглицеральдегидфосфаттың құрамында болады және оны пішінде сақтайды ATP.
алтыншы реакциягликолиз (фермент глицеральдегидфосфатдегидрогеназа) - глицеральдегидфосфаттың тотығуы және оған фосфор қышқылының қосылуы 1,3-дифосфоглицерин қышқылы мен НАДН-ның жоғары энергиялы қосылысының түзілуіне әкеледі.
IN жетінші реакция(фермент фосфоглицераткиназа) 1,3-дифосфоглицераттың құрамындағы фосфоэфир байланысының энергиясы АТФ түзілуіне жұмсалады. Реакция қосымша атау алды - , ол тотығу фосфорланудан айырмашылығы (митохондриялық мембранадағы сутегі иондарының электрохимиялық градиентінен) АТФ-да (реакция субстратынан) макроэргиялық байланыстарды алу үшін энергия көзін көрсетеді.
Сегізінші реакция- әсерінен алдыңғы реакцияда синтезделген 3-фосфоглицерат фосфоглицерат мутазы 2-фосфоглицератқа изомерленеді.
Тоғызыншы реакция– фермент энолаза 2-фосфоглицерин қышқылынан су молекуласын ажыратады және фосфоэнолпируват құрамында макроэргиялық фосфоэфирлік байланыстың түзілуіне әкеледі.
Оныншы реакциягликолиз басқа субстраттың фосфорлану реакциясы- пируваткиназа арқылы жоғары энергиялы фосфатты фосфоэнолпируваттан АДФ-ге беруден және пирожүзім қышқылының түзілуінен тұрады.
жалпы шолу
Гликолиздік жол 10 ретті реакциядан тұрады, олардың әрқайсысы басқа ферментпен катализденеді.
Гликолиз процесін шартты түрде екі кезеңге бөлуге болады. 2 АТФ молекуласының энергия шығынымен жүретін бірінші кезең глюкоза молекуласының глицеральдегид-3-фосфаттың 2 молекуласына ыдырауы болып табылады. Екінші кезеңде АТФ синтезімен жүретін глицеральдегид-3-фосфаттың NAD-тәуелді тотығуы жүреді. Гликолиздің өзі толық анаэробты процесс, яғни реакциялар жүруі үшін оттегінің болуын қажет етпейді.
Гликолиз - барлық дерлік тірі организмдерде белгілі ең көне метаболикалық процестердің бірі. Болжам бойынша, гликолиз бастапқы прокариоттарда 3,5 миллиард жыл бұрын пайда болған.
Локализация
Эукариоттық организмдердің жасушаларында глюкозаның ПВХ-ға дейін ыдырауын катализдейтін он фермент цитозолда, энергия алмасуына қатысы бар барлық басқа ферменттер митохондриялар мен хлоропластарда орналасады. Глюкоза жасушаға екі жолмен түседі: натрийге тәуелді симпорт (негізінен энтероциттерге және бүйрек түтікшелі эпителийіне) және тасымалдаушы белоктардың көмегімен глюкозаның жеңілдетілген диффузиясы. Бұл тасымалдаушы ақуыздардың жұмысы гормондармен және ең алдымен инсулинмен бақыланады. Ең бастысы, инсулин бұлшықет пен май тінінде глюкозаның тасымалдануын ынталандырады.
Нәтиже
Гликолиздің нәтижесі глюкозаның бір молекуласының пирожүзім қышқылының (ПВА) екі молекуласына айналуы және NAD∙H коферментінің түріндегі екі қалпына келтіретін эквиваленттің түзілуі болып табылады.
Гликолиздің толық теңдеуі:
Глюкоза + 2NAD + + 2ADP + 2P n \u003d 2NAD ∙ H + 2PVC + 2ATP + 2H 2 O + 2H +.Жасушада оттегі болмаған немесе жетіспейтін жағдайда пирожүзім қышқылы сүт қышқылына дейін тотықсызданады, онда гликолиздің жалпы теңдеуі келесідей болады:
Глюкоза + 2ADP + 2F n \u003d 2 лактат + 2ATP + 2H 2 O.Осылайша, глюкозаның бір молекуласының анаэробты ыдырауы кезінде жалпы таза АТФ шығымы АДФ субстратының фосфорлану реакцияларында алынған екі молекуланы құрайды.
Аэробты организмдерде гликолиздің соңғы өнімдері жасушалық тыныс алумен байланысты биохимиялық циклдерде әрі қарай өзгерістерге ұшырайды. Нәтижесінде бір глюкоза молекуласының барлық метаболиттерінің толық тотығуынан кейін жасушалық тыныс алудың соңғы сатысында – митохондриялық тыныс алу тізбегінде оттегінің қатысуымен жүретін тотығу фосфорлануы – әрбір глюкоза үшін қосымша 34 немесе 36 АТФ молекуласы қосымша синтезделеді. молекуласы.
Жол
Бірінші реакциягликолиз болып табылады фосфорлану 1 АТФ молекуласының энергия шығынымен тінге тән гексокиназа ферментінің қатысуымен пайда болатын глюкоза молекулалары; глюкозаның белсенді түрі түзіледі - глюкоза-6-фосфат (G-6-F):
Реакцияның жүруі үшін ортада АТФ молекулалық кешені байланысатын Mg 2+ иондарының болуы қажет. Бұл реакция қайтымсыз және бірінші болып табылады гликолиздің негізгі реакциясы.
Глюкозаның фосфорлануы екі мақсатты көздейді: біріншіден, глюкозаның бейтарап молекуласын өткізетін плазмалық мембрана теріс зарядты G-6-P молекулаларының өтуіне мүмкіндік бермейтіндіктен, фосфорланған глюкоза жасуша ішінде құлыпталады. Екіншіден, фосфорлану кезінде глюкоза биохимиялық реакцияларға қатыса алатын және метаболикалық циклдерге қатыса алатын белсенді түрге айналады.
Гексокиназаның бауыр изоферменті – глюкокиназаның қандағы глюкоза деңгейін реттеуде маңызы зор.
келесі реакцияда ( 2 ) G-6-P фосфоглюкоизомераза ферментімен айналады фруктоза-6-фосфат (F-6-F):

Бұл реакция үшін энергия қажет емес, реакция толығымен қайтымды. Бұл кезеңде фруктозаны фосфорлану арқылы гликолиз процесіне де қосуға болады.
Содан кейін бірден екі реакция бірінен соң бірі жүреді: фруктоза-6-фосфаттың қайтымсыз фосфорлануы ( 3 ) және нәтижесінде алынған альдолдың қайтымды бөлінуі фруктоза-1,6-бисфосфат (F-1,6-bF) екі триозаға ( 4 ).
F-6-F фосфорлануы басқа АТФ молекуласының энергиясын жұмсаумен фосфофруктокиназа арқылы жүзеге асады; бұл екінші негізгі реакциягликолиз, оның реттелуі жалпы гликолиздің қарқындылығын анықтайды.

Альдолдың бөлінуі F-1,6-bFфруктоза-1,6-бисфосфат альдолазаның әсерінен жүреді:

Төртінші реакция нәтижесінде дигидроксиацетонфосфатыЖәне глицеральдегид-3-фосфат, ал біріншісі бірден дерлік әрекет астында фосфотриоза изомеразаекіншісіне барады 5 ), одан әрі түрлендіруге қатысатын:

Глицеральдегидфосфаттың әрбір молекуласы NAD+ қатысуымен тотығады. глицеральдегидфосфатдегидрогеназабұрын 1,3-дифосфоглицерат (6 ):

Қайдан келеді 1,3-дифосфоглицерат, 1 позицияда макроэргиялық байланыс бар, фосфоглицераткиназа ферменті фосфор қышқылының қалдығын АДФ молекуласына тасымалдайды (реакция 7 ) - АТФ молекуласы түзіледі:

Бұл субстраттың фосфорлануының алғашқы реакциясы. Осы сәттен бастап глюкозаның ыдырау процесі энергия тұрғысынан тиімсіз болуын тоқтатады, өйткені бірінші кезеңдегі энергия шығындары өтеледі: 2 АТФ молекуласы синтезделеді (әрбір 1,3-дифосфоглицератқа бір) жұмсалған екінің орнына. реакциялар 1 Және 3 . Бұл реакцияның жүруі үшін цитозолда АДФ болуы қажет, яғни жасушада АТФ артық болғанда (және АДФ жетіспесе) оның жылдамдығы төмендейді. Метаболизденбейтін АТФ жасушада жиналмай, жай ғана жойылатындықтан, бұл реакция гликолиздің маңызды реттеушісі болып табылады.
Содан кейін кезекпен: фосфоглицерин мутазасы түзіледі 2-фосфоглицерат (8 ):

Энолаза формалары фосфоэнолпируват (9 ):

Ақырында, АДФ субстратының фосфорлануының екінші реакциясы пируват пен АТФ-ның энолдық формасының түзілуімен жүреді. 10 ):

Реакция пируваткиназаның әсерінен жүреді. Бұл гликолиздің соңғы негізгі реакциясы. Пируваттың энол түрінің пируватқа изомерленуі ферментативті емес жолмен жүреді.
Құрылғаннан бері F-1,6-bFэнергияның бөлінуімен реакциялар ғана жүреді 7 Және 10 мұнда АДФ субстрат фосфорлануы жүреді.
Әрі қарай дамыту
Гликолиз кезінде түзілетін пируват пен NAD∙H-тің соңғы тағдыры организмге және жасуша ішіндегі жағдайларға, әсіресе оттегінің немесе басқа электрон акцепторларының болуы немесе болмауына байланысты.
Анаэробты организмдерде пируват пен NAD∙H одан әрі ашытылады. Сүт қышқылын ашу кезінде, мысалы, бактерияларда пируват лактатдегидрогеназа ферментінің әсерінен сүт қышқылына дейін тотықсызданады. Ашытқыда ұқсас процесс алкогольдік ашыту болып табылады, онда соңғы өнімдер этанол мен көмірқышқыл газы болады. Бутирикалық және цитратты ашыту да белгілі.
Бутирикалық ашыту:
Глюкоза → май қышқылы + 2 CO 2 + 2 H 2 O.
Алкогольді ашыту:
Глюкоза → 2 этанол + 2 СО 2.
Лимонды ашыту:
Глюкоза → лимон қышқылы + 2 H 2 O.
Ашыту тамақ өнеркәсібінде өте маңызды.
Аэробтарда пируват әдетте трикарбон қышқылының айналымына (Кребс циклі) енеді, ал NAD∙H ақырында тотығу фосфорлану процесі арқылы митохондриялардағы тыныс алу тізбегіндегі оттегімен тотығады.
Адамның зат алмасуы негізінен аэробты болғанына қарамастан, қарқынды жұмыс істейтін қаңқа бұлшықеттерінде анаэробты тотығу байқалады. Оттегінің шектеулі қол жетімділігі жағдайында пируват сүт қышқылына айналады, өйткені көптеген микроорганизмдерде сүт қышқылы ашыту кезінде болады:
ПВХ + NAD∙H + H + → лактат + NAD + .
Ерекше қарқынды физикалық белсенділіктен кейін біраз уақыттан кейін пайда болатын бұлшықет ауыруы оларда сүт қышқылының жиналуымен байланысты.
Сүт қышқылының түзілуі метаболизмнің тұйық тармағы болып табылады, бірақ метаболизмнің соңғы өнімі емес. Лактатдегидрогеназаның әсерінен сүт қышқылы қайтадан тотығады, пируват түзеді, ол әрі қарай трансформацияға қатысады.
Гликолиздің реттелуі
Жергілікті және жалпы реттеуді ажыратыңыз.
Жергілікті реттеу жасуша ішіндегі әртүрлі метаболиттердің әсерінен ферменттердің белсенділігін өзгерту арқылы жүзеге асады.
Гликолиздің реттелуі тұтастай алғанда, бірден бүкіл организм үшін гормондардың әсерінен жүреді, олар екіншілік хабаршылардың молекулалары арқылы әсер етіп, жасушаішілік метаболизмді өзгертеді.
Инсулин гликолизді ынталандыруда маңызды рөл атқарады. Глюкагон мен адреналин гликолиздің ең маңызды гормондық ингибиторлары болып табылады.
Инсулин гликолизді ынталандырады:
- гексокиназа реакциясын белсендіру;
- фосфофруктокиназаны ынталандыру;
- пируваткиназаны ынталандыру.
Басқа гормондар да гликолизге әсер етеді. Мысалы, соматотропин гликолиз ферменттерін тежейді, ал қалқанша безінің гормондары стимулятор болып табылады.
Гликолиз бірнеше негізгі қадамдар арқылы реттеледі. Гексокиназамен катализденетін реакциялар ( 1 ), фосфофруктокиназа ( 3 ) және пируваткиназа ( 10 ) бос энергияның айтарлықтай төмендеуімен сипатталады және іс жүзінде қайтымсыз, бұл оларға гликолизді реттеудің тиімді нүктелері болуға мүмкіндік береді.
Гексокиназаның реттелуі
Гексокиназареакция өнімі – глюкоза-6-фосфатпен тежеледі, ол ферментпен аллостериялық байланысады, оның белсенділігін өзгертеді.
Жасушадағы G-6-P негізгі бөлігі гликогеннің ыдырауы арқылы түзілетіндіктен, гексокиназа реакциясы, шын мәнінде, гликолиздің пайда болуы үшін қажет емес, ал гликолизді реттеуде глюкозаның фосфорлануы қажет емес. ерекше маңызы бар. Гексокиназа реакциясы қандағы және жасушадағы глюкоза концентрациясын реттеудегі маңызды қадам болып табылады.
Фосфорлану кезінде глюкоза тасымалдаушы молекулалар арқылы мембрана арқылы тасымалдану мүмкіндігін жоғалтады, бұл оның жасушада жиналуына жағдай жасайды. Гексокиназа G-6-P тежелуі глюкозаның жасушаға түсуін шектейді, оның шамадан тыс жиналуын болдырмайды.
Бауырдың глюкокиназасы (гексокиназаның IV изотипі) глюкоза-6-фосфатпен тежелмейді, бауыр жасушалары G-6-P жоғары болса да глюкозаны жинақтауды жалғастырады, одан кейін гликоген синтезделеді. Басқа изотиптермен салыстырғанда глюкокиназа Михаэлис константасының жоғары мәніне ие, яғни фермент тамақтан кейін әрдайым дерлік пайда болатын глюкозаның жоғары концентрациясы жағдайында ғана толық қуатта жұмыс істейді.
Глюкоза-6-фосфат глюкоза-6-фосфатазаның әсерінен глюкозаға қайта айналуы мүмкін. Глюкокиназа және глюкоза-6-фосфатаза ферменттері қандағы глюкозаның қалыпты деңгейін ұстап тұруға қатысады.
Фосфофруктокиназаның реттелуі
Фосфофруктокиназа реакциясының қарқындылығы гликолиздің бүкіл өтуіне шешуші әсер етеді, ал фосфофруктокиназаны ынталандыру реттеудегі ең маңызды қадам болып саналады.
Фосфофруктокиназа (ФФК) – тетрамерлі фермент, ол екі конформациялық күйде (R және T), тепе-теңдікте және кезектесіп бірінен екіншісіне өтеді. АТФ бір мезгілде ПФК субстраты және аллостериялық ингибиторы болып табылады.
ҚФФ қосалқы бөлімшелерінің әрқайсысында екі АТФ байланыстыру орны бар: субстрат алаңы және тежелу орны. Субстрат алаңы кез келген тетрамер конформациясында АТФ-ны бекітуге бірдей қабілетті. Ал ингибирлеу орны АТФ-ны тек фермент Т-конформациялық күйде болғанда ғана байланыстырады.FPA үшін тағы бір субстрат - бұл ферментке жақсырақ R күйінде қосылатын фруктоза 6-фосфат. АТФ жоғары концентрациясында тежелу орны басып қалады, ферменттердің конформациялары арасындағы ауысулар мүмкін емес болады, ал фермент молекулаларының көпшілігі Р-6-Р-ны тіркей алмай, Т күйінде тұрақтанады. Дегенмен, АТФ фосфофруктокиназаның тежелуі ферменттің R-конформацияларына қосылатын АМФ арқылы басылады, осылайша F-6-P байланыстыру үшін ферменттің күйін тұрақтандырады.
Гликолиз мен глюконеогенездің ең маңызды аллостериялық реттеушісі болып табылады фруктоза 2,6-бисфосфат, бұл циклдердің аралық буыны емес. Фруктоза-2,6-бисфосфат фосфофруктокиназаны аллостериялық белсендіреді.
Фруктоза-2,6-бифосфаттың синтезі арнайы бифункционалды фермент - фосфофруктокиназа-2/фруктоза-2,6-бифосфатаза (ФФК-2/Ф-2,6-ВПаза) арқылы катализденеді. Фосфорланбаған түрінде белок фосфофруктокиназа-2 ретінде белгілі және фруктоза 6-фосфатқа каталитикалық белсенділік танытып, фруктоза 2-6-бисфосфатты түзеді. Нәтижесінде ҚФФ белсенділігі айтарлықтай ынталандырылады және фруктоза-1,6-бифосфатазаның белсенділігі күшті тежеледі. Яғни, ҚФФ-2 белсенділігі жағдайында гликолиз бен глюконеогенез арасындағы осы реакцияның тепе-теңдігі біріншіге қарай ығысады – фруктоза-1,6-бисфосфат синтезделеді.
Фосфорланған күйде бифункционалды ферменттің киназалық белсенділігі болмайды, керісінше оның молекуласында P2,6BP гидролизін P6P және бейорганикалық фосфатқа түсіретін учаске белсендіріледі. Бифункционалды ферменттің фосфорлануының метаболикалық әсері ПФК-ның аллостериялық стимуляциясы тоқтап, F-1,6-ВПазаның аллостериялық тежелуі жойылады және тепе-теңдік глюконеогенезге қарай ығысады. F6F, содан кейін глюкоза өндіріледі.
Бифункционалды ферменттің өзара конверсиясы cAMP-тәуелді протеинкиназа (ПК) арқылы жүзеге асады, ол өз кезегінде қанда айналатын пептидтік гормондармен реттеледі.
Қандағы глюкозаның концентрациясы төмендеген кезде инсулиннің түзілуі де тежеледі, ал глюкагонның бөлінуі, керісінше, ынталандырылады және оның қандағы концентрациясы күрт көтеріледі. Глюкагон (және басқа контринсулярлық гормондар) бауыр жасушаларының плазмалық мембранасындағы рецепторлармен байланысып, мембраналық аденилатциклазаның белсендірілуін тудырады. Аденилатциклаза АТФ циклдік АМФ-ға айналуын катализдейді. cAMP протеинкиназаның реттеуші суббірлігімен байланысады, бұл бірқатар ферменттерді, соның ішінде бифункционалды FFK-2/P-2,6-BPase-ны фосфорилдендіретін оның каталитикалық суббірліктерінің босатылуын және белсендірілуін тудырады. Сонымен бірге бауырда глюкозаны тұтыну тоқтап, глюконеогенез және гликогенолиз белсендіріліп, нормогликемияны қалпына келтіреді.
пируваткиназа
Гликолиздің реттелуі жүзеге асырылатын келесі қадам соңғы реакция – пируваткиназаның әсер ету кезеңі болып табылады. Пируваткиназа үшін реттеуші қасиеттері бар бірқатар изоферменттер де сипатталған.
Бауыр пируваткиназасы(L-типі) фосфорланумен, алстерикалық эффекторлармен және ген экспрессиясының реттелуімен реттеледі. Фермент АТФ және ацетил-КоА арқылы тежеліп, фруктоза-1,6-бисфосфатпен белсендіріледі. АТФ пируваткиназасының тежелуі АТФ ПФК әрекетіне ұқсас жүреді. АТФ ферментті тежеу орнымен байланысуы оның фосфоэнолпируватқа жақындығын төмендетеді. Бауыр пируваткиназасы протеинкиназамен фосфорланады және тежеледі, сондықтан да гормоналды бақылауда болады. Сонымен қатар, бауыр пируваткиназасының белсенділігі де сандық түрде реттеледі, яғни оның синтезінің деңгейін өзгерту арқылы. Бұл баяу, ұзақ мерзімді реттеу. Рациондағы көмірсулардың көбеюі пируваткиназаны кодтайтын гендердің экспрессиясын ынталандырады, нәтижесінде жасушадағы ферменттің деңгейі жоғарылайды.
М-типті пируваткиназамида, бұлшықеттерде және басқа глюкозаны қажет ететін тіндерде табылған ақуыз киназамен реттелмейді. Бұл іргелі болып табылады, өйткені бұл тіндердің метаболизмі тек ішкі қажеттіліктермен анықталады және қандағы глюкоза деңгейіне байланысты емес.
Бұлшықет пируваткиназасы қандағы глюкоза деңгейін төмендету немесе гормоналды босату сияқты сыртқы әсерлерге ұшырамайды. Бауыр изоферментінің фосфорлануына және тежелуіне әкелетін жасушадан тыс жағдайлар М-типті пируваткиназаның белсенділігін өзгертпейді. Яғни, жолақты бұлшықеттердегі гликолиздің қарқындылығы тек жасуша ішіндегі жағдайлармен анықталады және жалпы реттелуге байланысты емес.
Мағынасы
Гликолиз – ерекше маңызы бар катаболикалық жол. Ол жасушалық реакцияларды, соның ішінде ақуыз синтезін энергиямен қамтамасыз етеді. Гликолиздің аралық өнімдері майларды синтездеуде қолданылады. Пируват аланинді, аспартатты және басқа қосылыстарды синтездеу үшін де қолданылуы мүмкін. Гликолиздің арқасында митохондриялық өнімділік және оттегінің қолжетімділігі қысқа мерзімді төтенше жүктемелер кезінде бұлшықет күшін шектемейді.
да қараңыз
Сілтемелер
- гликолиз (ағылшынша)
Анаэробты гликолиз – адам және жануарлар ұлпаларында оттегін пайдаланбай жүретін глюкозаның бірізді түрленуінің күрделі ферментативті процесі (28-сурет).
Пирувин қышқылының сүт қышқылына қайтымды айналуын лактатдегидрогеназа катализдейді:
Гликолиздің жалпы нәтижесі келесі теңдеумен өрнектеледі: C 6 H 12 O 6 + 2H 3 RO 4 + 2ADP \u003d 2C 3 H 6 O 3 + 2ATP + 2H 2 O
Сонымен, анаэробты гликолиз кезіндегі АТФ таза шығымы 1 моль глюкозаға 2 моль АТФ құрайды. Дәл анаэробты гликолиздің арқасында адам және жануар организмі оттегі тапшылығы жағдайында белгілі бір уақыт аралығында бірқатар физиологиялық функцияларды орындай алады.
Бактериялардағы бұл процесс сүтқышқылды ашыту деп аталады: ол ашытылған сүт өнімдерін дайындаудың негізінде жатыр. Анаэробты гликолиз жасушалардың цитозолында жүреді, бұл үшін қажетті барлық ферменттер бар және митохондриялық тыныс алу тізбегін қажет етпейді. Анаэробты гликолиз процесінде АТФ субстраттың фосфорлану реакциялары есебінен түзіледі.
Анаэробты жағдайда ашытқыларда ұқсас процесс жүреді - алкогольдік ашыту, бұл жағдайда пирожүзім қышқылы ацетальдегидті қалыптастыру үшін декарбоксилделеді, содан кейін этил спиртіне дейін тотықсызданады:
CH 3 -CO-COOH → CH 3 -CHO + CO 2;
CH 3 -CHO + OVERHAND + H + → CH 3 -CH 2 -OH + OVER +.

28-сурет. Глюкозаның анаэробты гликолизінің диаграммасы
10.6. Глюкозаның аэробты ыдырауы
Глюкозаның аэробты ыдырауы үш кезеңнен тұрады:
1) глюкозаның пирожүзім қышқылына (пируватқа) айналуы – аэробты гликолиз. Бұл бөлік жоғарыда қарастырылған анаэробты гликолиз процесіне ұқсас, оның соңғы сатысын қоспағанда (пируваттың сүт қышқылына айналуы);
2) катаболизмнің жалпы жолы;
3) митохондриялық электрондарды тасымалдау тізбегі – ұлпаның тыныс алу процесі.
Катаболизмнің жалпы жолы
Катаболизмнің жалпы жолы екі кезеңнен тұрады.
1 кезең – пирожүзім қышқылының тотығу декарбоксилденуі. Бұл көп ферменттік жүйе – пируватдегидрогеназа кешенімен катализденетін күрделі көп сатылы процесс; митохондрияда локализацияланған (ішкі мембрана және матрица) және жалпы жалпы схемамен көрсетілуі мүмкін:
CH 3 -CO-COOH + HS-KoA + OVER + → CH 3 -CO-SkoA + AVERHAND + H + + CO 2.
2-ші кезең – Кребс циклі (цитраттық цикл, немесе үшкарбон және дикарбон қышқылдарының циклі) (29-сурет); митохондрияларда (матрицада) локализацияланған. Бұл циклде ацетил-КоА құрамына кіретін ацетил қалдығы бірқатар бастапқы сутегі донорларын құрайды. Әрі қарай сутегі дегидрогеназалардың қатысуымен тыныс алу тізбегіне енеді. Цитрат циклі мен тыныс алу тізбегінің конъюгацияланған әрекеті нәтижесінде ацетил қалдығы CO 2 және H 2 O тотығады. Аэробты ыдырау кезіндегі глюкозаның барлық түрлену тізбегі үшін жалпы теңдеу келесідей:
C 6 H 12 O 2 + 6O 2 → 6CO 2 + 6H 2 O
Аэробты ыдыраудың энергетикалық әсері 1 глюкоза молекуласының ыдырауы кезінде 38 АТФ молекуласының синтезі болып табылады. Сонымен, энергия жағынан глюкозаның көмірқышқыл газы мен суға дейін толық тотығуы анаэробты гликолизге қарағанда тиімдірек процесс болып табылады. Оттегі анаэробты гликолизді тежейді, сондықтан артық оттегі болған жағдайда өсімдік және жануарлар тіндерінде анаэробты гликолизден (ашыту) тыныс алуға (аэробты гликолиз) өту байқалады, яғни. жасушаларды энергия алудың тиімді және үнемді тәсіліне ауыстыру (Пастер эффектісі). Ағзаны энергиямен қамтамасыз етудегі анаэробты гликолиздің рөлі әсіресе қысқа мерзімді қарқынды жұмыс кезінде, митохондрияға оттегінің тасымалдану механизмінің күші аэробты гликолизді қамтамасыз ету үшін жеткіліксіз болған кезде үлкен. Сонымен, ~ 30 секундқа (200 м-ге) жүгіру толығымен анаэробты гликолизмен қамтамасыз етіледі, ал тыныс алудың жоғарылауымен анаэробты гликолиз жылдамдығы төмендейді, ал аэробты ыдырау жылдамдығы артады. 4-5 минуттан кейін. жүгіру (1,5 км) – энергияның жартысы анаэробты, жартысы аэробты процесс арқылы беріледі. 30 минуттан кейін. (10 км жүгіру) - энергия толығымен дерлік аэробтық процесспен қамтамасыз етіледі.
Эритроциттерде митохондрия мүлде болмайды, олардың АТФ-ға қажеттілігі анаэробты гликолиз арқылы толық қанағаттандырылады.
(грек тілінен glykys – тәтті және lysis – ыдырау, ыдырау) – тірі организмдерде энергия түзудің үш негізгі (гликолиз, Кребс циклі және Энтнер-Доудоров жолы) жолдарының бірі. Бұл жануарлар тіндеріндегі көмірсулардың (негізінен глюкоза мен гликогеннің) анаэробты (яғни бос O 2 қатысуын қажет етпейтін) ферменттік гидролитикалық емес ыдырау процесі, аденозин үшфосфор қышқылының (АТФ) синтезімен бірге жүреді және сүт қышқылының түзілуі. Гликолиз бұлшықет жасушалары, сперматозоидтар, өсіп келе жатқан тіндер (соның ішінде ісіктер) үшін маңызды, өйткені оттегі болмаған кезде энергияның сақталуын қамтамасыз етеді. Бірақ гликолиз О 2 (аэробты гликолиз) болған кезде де белгілі – эритроциттерде, көз торында, ұрық тіндерінде туылғаннан кейін бірден және ішектің шырышты қабатында. Гликолизді зерттеуге Г. және К.Кори, сондай-ақ биохимияның О.Мейерхоф және Г.Эмбден сияқты ізашарлары үлкен үлес қосты. Гликолиз биохимиялық реакциялардың толық шифрланған алғашқы тізбегі болды (19 ғасырдың аяғынан 1940 жылдарға дейін). Кейбір жасушалардағы (эритроциттер, май тіндері) гексомонофосфатты шунт немесе пентозофосфат жолы да энергия жеткізуші рөлін атқара алады.
Гликолиз процесіне глюкозадан басқа глицерин, кейбір аминқышқылдары және басқа субстраттар қатысуы мүмкін. Гликолиздің негізгі субстраты гликоген болып табылатын бұлшықет тінінде процесс 2 және 3 реакциялардан басталады ( см. схемасы) және гликогенолиз деп аталады. Гликогенолиз және гликолиз үшін ортақ аралық зат глюкоза-6-фосфат болып табылады. Гликоген түзілудің кері жолы гликогенез деп аталады.
Гликолиз кезінде түзілетін өнімдер кейінгі тотығу трансформациялары үшін субстраттар болып табылады ( см. Үшкарбон қышқылының циклі немесе Кребс циклі). Гликолизге ұқсас процестер өсімдік, ашытқы және бактерия жасушаларында болатын сүт қышқылы, май, спирт, глицерин ашыту болып табылады. Гликолиздің жеке кезеңдерінің қарқындылығы қышқылдық – рН – рН (оңтайлы рН 7–8), температура мен ортаның иондық құрамына байланысты. Гликолиз реакцияларының реттілігі ( см.схемасы) жақсы зерттелген және аралық өнімдер анықталған. Жасуша шырынында болатын еритін гликолиз ферменттері кристалды немесе тазартылған түрде оқшауланған.
Гликолиздің жеке сатыларын жүзеге асыратын ферменттер:
1. Гексокиназа CF2.7.1.1 (немесе глюкокиназа CF2.7.1.2)
2. Гликоген фосфорилаза CP2.4.1.1
3. Фосфоглюкомутаза CP2.7.5.1
4. Глюкозафосфат изомераза CP5.3.1.9
5. Фосфофруктокиназа KF2.7.1.11
6. Фруктоза бисфосфат альдолаза EC4.1.2.13
7. Триозафосфат изомераза CP5.3.1.1
8, 9. Глицеральдегидфосфатдегидрогеназа CP1.2.1.12
10. Фосфоглицераткиназа CP2.7.2.3
11. Фосфоглицеромутаза EC2.7.5.3
12. Энолаза KF4.2.1.11
13. Пируваткиназа KF2.7.1.40
14. Лактатдегидрогеназа CP1.1.1.27
Гликолиз қанттардың фосфор туындыларының түзілуінен басталады, бұл субстраттың циклдік формасының ациклді, неғұрлым реактивті түрге айналуына ықпал етеді. Гликолиз жылдамдығын реттейтін реакциялардың бірі - фосфорилаза ферментімен катализденетін 2-реакция. Гликолиздегі орталық реттеуші рөл фосфофруктокиназа ферментіне жатады (реакция 5), оның белсенділігі АТФ және цитратпен тежеледі, бірақ оның ыдырау өнімдерімен ынталандырылады. Гликолиздің орталық буыны гликолитикалық тотықсыздану (реакциялар 8-10) болып табылады, ол 3-фосфоглицеральдегидтің 3-фосфоглицерин қышқылына дейін тотығуымен және никотинамид аденин динуклеотиді (NAD) коферментінің тотықсыздануымен жүретін тотығу-тотықсыздану процесі. Бұл түрлендірулер фосфоглицераткиназаның қатысуымен 3-фосфоглицеральдегиддегидрогеназа (ФФГ) арқылы жүзеге асады. Бұл гликолиздегі жалғыз тотығу сатысы, бірақ ол бос оттегіні қажет етпейді, тек NAD+ болуы қажет, содан кейін ол NAD-H 2-ге дейін төмендейді.
Тотығу-тотықсыздану (тотықсыздану процесі) нәтижесінде энергия бөлінеді, ол субстраттың фосфорлану процесінде жинақталады (энергияға бай АТФ қосылысы түрінде). АТФ түзілуін қамтамасыз ететін екінші реакция 13-реакция – пирожүзім қышқылының түзілуі. Анаэробты жағдайда гликолиз лактатдегидрогеназаның әсерінен және тотықсызданған NAD-тың қатысуымен сүт қышқылының түзілуімен (14-ші реакция) аяқталады, ол NAD-қа дейін тотығады (NAD-H 2) және қайтадан тотығу сатысында қолдануға болады. . Аэробты жағдайда пирожүзім қышқылы митохондрияларда Кребс циклі кезінде тотығады.
Сонымен, глюкозаның 1 молекуласы ыдырағанда 2 молекула сүт қышқылы және 4 молекула АТФ түзіледі. Сонымен бірге гликолиздің бірінші кезеңдерінде (1, 5 реакцияларды қараңыз) 1 глюкоза молекуласына 2 АТФ молекуласы жұмсалады. Гликогенолиз процесінде 3 АТФ молекуласы түзіледі, себебі. глюкоза-6-фосфат алу үшін АТФ жұмсаудың қажеті жоқ. Гликолиздің алғашқы тоғыз реакциясы оның эндергоникалық (энергияны сіңірумен) фазасы, ал соңғы реакциялары оның экзергоникалық (энергия бөлінуімен) фазасы. Гликолиз процесінде глюкозаның толық тотығуымен (СО 2 және H 2 O-ға дейін) алуға болатын теориялық энергияның тек 7%-ға жуығы бөлінеді. Дегенмен, АТФ түріндегі энергияны сақтаудың жалпы тиімділігі 35–40% құрайды, ал практикалық жағдайларда жасушалар одан да жоғары болуы мүмкін.
Глицеральдегидфосфатдегидрогеназа мен лактатдегидрогеназа іштей байланысқан (біреуіне NAD+ қажет, екіншісі НАД+ құрайды), бұл коферменттің айналымын қамтамасыз етеді. Бұл, мүмкін, терминал дегидрогеназаның негізгі биохимиялық маңызы.
1, 5 және 13-тен басқа барлық гликолиз реакциялары қайтымды. Дегенмен, глюкозаны (1-реакция) немесе фруктоза монофосфатты (5-реакция) тиісті ферменттердің қатысуымен фосфор қышқылын гидролиздік ыдырату арқылы олардың фосфор туындыларынан алуға болады; 13 реакциясы іс жүзінде қайтымсыз, шамасы, фосфор тобының гидролизінің жоғары энергиясына байланысты (шамамен 13 ккал/моль). Сондықтан гликолиз өнімдерінен глюкозаның түзілуі басқа жолмен жүреді.
О 2 болғанда гликолиз жылдамдығы төмендейді (Пастер эффектісі). Кейбір қарқынды гликолиздеуші тіндерде гликолиз арқылы тіннің тыныс алуын басу мысалдары бар (Крабтри эффектісі). Анаэробты және аэробты тотығу процестерінің өзара байланысының механизмдері толық зерттелмеген. Гликолиз және гликогенез процестерінің бір мезгілде реттелуі ағзаның қажеттіліктеріне байланысты осы жолдардың әрқайсысында көміртегі ағынын бірегей түрде анықтайды. Бақылау екі деңгейде жүзеге асырылады - гормондық (жоғары сатыдағы жануарларда екінші хабаршылардың қатысуымен реттеуші каскадтар арқылы) және метаболикалық (барлық ағзаларда).
Игорь Рапанович