პლაზმური მემბრანის ფუნქციები. პლაზმური მემბრანა პლაზმის სტრუქტურა და ფუნქცია
პლაზმური მემბრანა განსაკუთრებულ პოზიციას იკავებს, რადგან ის ზღუდავს უჯრედს გარეთ და პირდაპირ არის დაკავშირებული უჯრედგარე გარემოსთან. მისი სისქე დაახლოებით 10 ნმ-ია და ყველაზე სქელი უჯრედის მემბრანაა. ძირითადი კომპონენტებია ცილები (60%-ზე მეტი), ლიპიდები (დაახლოებით 40%) და ნახშირწყლები (დაახლოებით 1%). ყველა სხვა უჯრედის მემბრანის მსგავსად, ის სინთეზირდება EPS არხებში.
პლაზმური ფუნქციები.
ტრანსპორტი.
პლაზმური მემბრანა ნახევრად გამტარია, ე.ი. მასში სხვადასხვა სიჩქარით შერჩევით სხვადასხვა მოლეკულა გადის. მემბრანის გასწვრივ ნივთიერებების ტრანსპორტირების ორი გზა არსებობს: პასიური და აქტიური ტრანსპორტი.
პასიური ტრანსპორტი.პასიური ტრანსპორტი ან დიფუზია არ საჭიროებს ენერგიის შეყვანას. დაუხტვილი მოლეკულები დიფუზირდება კონცენტრაციის გრადიენტის გასწვრივ, დამუხტული მოლეკულების ტრანსპორტირება დამოკიდებულია წყალბადის პროტონების კონცენტრაციის გრადიენტზე და ტრანსმემბრანული პოტენციალის სხვაობაზე, რომლებიც გაერთიანებულია ელექტროქიმიურ პროტონულ გრადიენტში. როგორც წესი, მემბრანის შიდა ციტოპლაზმური ზედაპირი ატარებს უარყოფით მუხტს, რაც ხელს უწყობს დადებითად დამუხტული იონების შეღწევას უჯრედში. არსებობს ორი სახის დიფუზია: მარტივი და მსუბუქი.
მარტივი დიფუზია ტიპიურია მცირე ნეიტრალური მოლეკულებისთვის (H 2 O, CO 2, O 2), ასევე ჰიდროფობიური დაბალი მოლეკულური წონის ორგანული ნივთიერებებისთვის. ამ მოლეკულებს შეუძლიათ გაიარონ მემბრანის ფორებში ან არხებში მემბრანის ცილებთან ყოველგვარი ურთიერთქმედების გარეშე, სანამ კონცენტრაციის გრადიენტი შენარჩუნებულია.
გამარტივებული დიფუზია დამახასიათებელია ჰიდროფილური მოლეკულებისთვის, რომლებიც მემბრანის გასწვრივ ტრანსპორტირდება აგრეთვე კონცენტრაციის გრადიენტის გასწვრივ, მაგრამ პრინციპის მიხედვით სპეციალური მემბრანული მატარებელი ცილების დახმარებით. ფორმები.
ხელშემწყობი დიფუზია ძალზე შერჩევითია, ვინაიდან გადამზიდავ ცილას აქვს ტრანსპორტირებული ნივთიერების დამაკავშირებელი ცენტრი და ტრანსფერს თან ახლავს ცილის კონფორმაციული ცვლილებები. გაადვილებული დიფუზიის ერთ-ერთი შესაძლო მექანიზმი ასეთია: სატრანსპორტო ცილა (ტრანსლოკაზა) აკავშირებს ნივთიერებას, შემდეგ უახლოვდება მემბრანის მოპირდაპირე მხარეს, ათავისუფლებს ამ ნივთიერებას, იღებს საწყის კონფორმაციას და კვლავ მზად არის სატრანსპორტო ფუნქციის შესასრულებლად. ცოტა რამ არის ცნობილი იმის შესახებ, თუ როგორ მოძრაობს თავად ცილა. გადაცემის კიდევ ერთი შესაძლო მექანიზმი მოიცავს რამდენიმე მატარებელი ცილის მონაწილეობას. ამ შემთხვევაში, თავდაპირველად შეკრული ნაერთი თავად გადადის ერთი ცილიდან მეორეზე, თანმიმდევრულად აკავშირებს ამა თუ იმ ცილას, სანამ ის მემბრანის მოპირდაპირე მხარეს არ იქნება.
Აქტიური ტრანსპორტი.ასეთი ტრანსპორტირება ხდება მაშინ, როდესაც გადატანა ხორციელდება კონცენტრაციის გრადიენტის საწინააღმდეგოდ. ის მოითხოვს ენერგიის დახარჯვას უჯრედის მიერ. აქტიური ტრანსპორტი ემსახურება უჯრედის შიგნით ნივთიერებების დაგროვებას. ენერგიის წყარო ხშირად არის ATP. აქტიური ტრანსპორტირებისთვის, ენერგიის წყაროს გარდა, აუცილებელია მემბრანის ცილების მონაწილეობა. ცხოველთა უჯრედებში ერთ-ერთი აქტიური სატრანსპორტო სისტემა პასუხისმგებელია Na და K + იონების ტრანსპორტირებაზე უჯრედის მემბრანაში. ამ სისტემას ე.წ ნა + - K * -ტუმბო.იგი პასუხისმგებელია უჯრედშიდა გარემოს შემადგენლობის შენარჩუნებაზე, რომელშიც K + იონების კონცენტრაცია უფრო მაღალია, ვიდრე Na * იონების.
ორივე იონის კონცენტრაციის გრადიენტი შენარჩუნებულია K + უჯრედის შიგნით და Na + გარეთ გადატანით. ორივე ტრანსპორტი ხდება კონცენტრაციის გრადიენტის საწინააღმდეგოდ. იონების ეს განაწილება განსაზღვრავს უჯრედებში წყლის შემცველობას, ნერვული უჯრედების და კუნთოვანი უჯრედების აგზნებადობას და ნორმალური უჯრედების სხვა თვისებებს. Na + -K + - ტუმბო არის ცილა - ტრანსპორტირება ATPase.ამ ფერმენტის მოლეკულა არის ოლიგომერი და გადის მემბრანაში. ტუმბოს მუშაობის სრული ციკლის დროს 3 Na + იონი გადადის უჯრედიდან უჯრედშორის ნივთიერებაში, ხოლო საპირისპირო მიმართულებით - 2 K + იონი, ხოლო ატფ-ის მოლეკულის ენერგია გამოიყენება. არსებობს კალციუმის იონების გადაცემის სატრანსპორტო სისტემები (Ca 2+ -ATPase), პროტონული ტუმბოები (H + -ATPase) და ა.შ.
მემბრანის მეშვეობით ნივთიერების აქტიური გადატანა, რომელიც განხორციელდა სხვა ნივთიერების კონცენტრაციის გრადიენტის ენერგიის გამო, ე.წ. სიმპტომი... ამ შემთხვევაში, სატრანსპორტო ატფ-აზას აქვს ორივე ნივთიერების დამაკავშირებელი ადგილები. ანტიპორტიარის ნივთიერების მოძრაობა მისი კონცენტრაციის გრადიენტის წინააღმდეგ. ამ შემთხვევაში, სხვა ნივთიერება მოძრაობს საპირისპირო მიმართულებით მისი კონცენტრაციის გრადიენტის გასწვრივ. სიმპტომი და ანტიპორტი (კოტრანსპორტი) შეიძლება მოხდეს ნაწლავიდან ამინომჟავების შეწოვისას და პირველადი შარდიდან გლუკოზის რეაბსორბციის დროს Na +, K + -ATPase მიერ შექმნილი Na + იონების კონცენტრაციის გრადიენტის ენერგიის გამოყენებით.
ტრანსპორტის კიდევ 2 ტიპია ენდოციტოზი და ეგზოციტოზი.
ენდოციტოზი- უჯრედის მიერ დიდი ნაწილაკების დაჭერა. ენდოციტოზის რამდენიმე გზა არსებობს: პინოციტოზი და ფაგოციტოზი. ჩვეულებრივ ქვეშ პინოციტოზიგაიგეთ უჯრედის მიერ თხევადი კოლოიდური ნაწილაკების დაჭერა, ქვეშ ფაგოციტოზი- კორპუსების დაჭერა (უფრო მკვრივი და დიდი ნაწილაკები სხვა უჯრედებამდე). პინო- და ფაგოციტოზის მექანიზმი განსხვავებულია.
ზოგადად, გარედან მყარი ნაწილაკების ან თხევადი წვეთების უჯრედში შეღწევას ჰეტეროფაგია ეწოდება. ეს პროცესი ყველაზე გავრცელებულია პროტოზოებში, მაგრამ ძალიან მნიშვნელოვანია ადამიანებში (ისევე როგორც სხვა ძუძუმწოვრებში). ჰეტეროფაგია მნიშვნელოვან როლს ასრულებს სხეულის დაცვაში (სეგმენტირებული ნეიტროფილები - გრანულოციტები; მაკროფაგოციტები), ძვლოვანი ქსოვილის რემოდელირება (ოსტეოკლასტები), ფარისებრი ჯირკვლის ფოლიკულების მიერ თიროქსინის წარმოქმნა, პროქსიმალურ ნეფრონში ცილის და სხვა მაკრომოლეკულების რეაბსორბცია. და სხვა პროცესები.
პინოციტოზი.
იმისათვის, რომ გარე მოლეკულები შევიდნენ უჯრედში, ისინი ჯერ უნდა იყოს შეკრული გლიკოკალიქსის რეცეპტორებით (მემბრანის ზედაპირულ ცილებთან შეკრული მოლეკულების ნაკრები) (ნახ.).
ასეთი შებოჭვის ადგილზე, კლატრინის ცილის მოლეკულები გვხვდება პლაზმალემის ქვეშ. პლაზმალემა, გარედან მიმაგრებულ მოლეკულებთან და ციტოპლაზმის ქვეშ მდებარე კლატრინთან ერთად, იწყებს შეჭრას. პროტრუზია უფრო ღრმა ხდება, მისი კიდეები უახლოვდება და შემდეგ იხურება. შედეგად, ბუშტი იშლება პლაზმალემისგან, რომელიც ატარებს დატყვევებულ მოლეკულებს. კლატრინი მის ზედაპირზე გამოიყურება როგორც არათანაბარი საზღვარი ელექტრონულ მიკროგრაფებზე, ამიტომ ასეთ ბუშტებს უწოდებენ შემოსაზღვრულ.
კლატრინი ხელს უშლის ბუშტუკების უჯრედშიდა მემბრანებთან მიმაგრებას. ამრიგად, შემოსაზღვრული ვეზიკულები შეიძლება თავისუფლად გადაიტანონ უჯრედში ციტოპლაზმის ზუსტად იმ ნაწილებში, სადაც მათი შიგთავსი უნდა იყოს გამოყენებული. ასე, კერძოდ, სტეროიდული ჰორმონები მიეწოდება ბირთვს. თუმცა, ჩვეულებრივ, შემოსაზღვრული ვეზიკულები პლაზმურიდან გაყოფის შემდეგ მალევე იშლება საზღვარი. კლატრინი გადადის პლაზმალემაში და შეუძლია კვლავ მონაწილეობა მიიღოს ენდოციტოზის რეაქციებში.
ციტოპლაზმის უჯრედის ზედაპირზე უფრო მუდმივი ვეზიკულები - ენდოსომებია. შემოსაზღვრული ვეზიკულები ათავისუფლებს კლატრინს და ერწყმის ენდოსომებს, ზრდის ენდოსომების მოცულობას და ზედაპირს. შემდეგ ენდოსომების ჭარბი ნაწილი იშლება ახალი ვეზიკულის სახით, რომელშიც არ არის უჯრედში შესული ნივთიერებები, ისინი რჩება ენდოსომაში. ახალი ვეზიკულა მიმართულია უჯრედის ზედაპირზე და ერწყმის მემბრანას. შედეგად, პლაზმალემის დაქვეითება, რომელიც ხდება შემოსაზღვრული ვეზიკულის გაყოფის დროს, აღდგება, ხოლო მისი რეცეპტორები ასევე უბრუნდებიან პლაზმალემას.
ენდოსომები ჩაეფლო ციტოპლაზმაში და შერწყმულია ლიზოსომის მემბრანებთან. ასეთი მეორადი ლიზოსომის შიგნით შემომავალი ნივთიერებები განიცდიან სხვადასხვა ბიოქიმიურ ტრანსფორმაციას. პროცესის დასრულების შემდეგ, ლიზოსომის მემბრანა შეიძლება დაიშალოს ფრაგმენტებად და ლიზოსომის დაშლის პროდუქტები და შიგთავსი ხელმისაწვდომი გახდეს უჯრედშიდა მეტაბოლური რეაქციებისთვის. მაგალითად, ამინომჟავები უკავშირდება tRNA-ს და მიეწოდება რიბოზომებს, ხოლო გლუკოზა შეიძლება შევიდეს გოლგის კომპლექსში ან აგრანულარული EPS-ის ტუბულებში.
მიუხედავად იმისა, რომ ენდოსომებს არ აქვთ კლატრინის საზღვარი, ყველა მათგანი არ ერწყმის ლიზოსომებს. ზოგიერთი მათგანი მიმართულია ერთი უჯრედის ზედაპირიდან მეორეზე (თუ უჯრედები ქმნიან ეპითელურ შრეს). იქ ენდოსომის მემბრანა ერწყმის პლაზმოლემას და შიგთავსი გამოიყოფა. შედეგად, ნივთიერებები უჯრედში გადადის ერთი გარემოდან მეორეში ცვლილებების გარეშე. ამ პროცესს ე.წ ტრანსციტოზი... ცილის მოლეკულები, განსაკუთრებით იმუნოგლობულინები, ასევე შეიძლება ტრანსპორტირება ტრანსციტოზით.
ფაგოციტოზი.
თუ დიდ ნაწილაკს აქვს მოლეკულური დაჯგუფებები მის ზედაპირზე, რომელთა ამოცნობა შესაძლებელია უჯრედის რეცეპტორებით, ის აკავშირებს. არავითარ შემთხვევაში, უცხო ნაწილაკები ყოველთვის არ ფლობენ ასეთ ჯგუფებს. თუმცა, სხეულში მოხვედრისას ისინი გარშემორტყმულია იმუნოგლობულინების (ოპსონინების) მოლეკულებით, რომლებიც ყოველთვის გვხვდება სისხლში და უჯრედგარე გარემოში. იმუნოგლობულინები ყოველთვის აღიარებულია ფაგოციტური უჯრედების მიერ.
მას შემდეგ, რაც ოფსონინები, რომლებიც ფარავს უცხო ნაწილაკს, აკავშირებენ ფაგოციტის რეცეპტორებს, მისი ზედაპირის კომპლექსი აქტიურდება. აქტინის მიკროფილამენტები იწყებენ ურთიერთქმედებას მიოზინთან და იცვლება უჯრედის ზედაპირის კონფიგურაცია. ნაწილაკების ირგვლივ გადაჭიმულია ფაგოციტების ციტოპლაზმის გამონაზარდები. ისინი ფარავს ნაწილაკების ზედაპირს და აერთიანებს მასზე. გამონაზარდების გარე ფოთლები ერწყმის, უჯრედის ზედაპირს ხურავს.
გამონაზარდების ღრმა ფურცლები ქმნიან მემბრანას შთანთქმის ნაწილაკების ირგვლივ - ის იქმნება ფაგოსომა.ფაგოსომა ერწყმის ლიზოსომებს, რაც იწვევს მათ კომპლექსს - ჰეტეროლისომა (ჰეტეროსომა,ან ფაგოლიზოსომა).მასში ხდება ნაწილაკების დატყვევებული კომპონენტების ლიზისი. ლიზისის ზოგიერთი პროდუქტი ამოღებულია ჰეტეროსომიდან და გამოიყენება უჯრედის მიერ, ზოგი კი შესაძლოა არ ექვემდებარებოდეს ლიზოსომური ფერმენტების მოქმედებას. ეს ნარჩენები ქმნიან ნარჩენ სხეულებს.
პოტენციურად ყველა უჯრედს აქვს ფაგოციტოზის უნარი, მაგრამ სხეულში მხოლოდ რამდენიმე სპეციალიზდება ამ მიმართულებით. ეს არის ნეიტროფილური ლეიკოციტები და მაკროფაგები.
ეგზოციტოზი.
ეს არის ნივთიერებების ამოღება უჯრედიდან. პირველი, მსხვილმოლეკულური ნაერთები გამოიყოფა გოლჯის კომპლექსში სატრანსპორტო ბუშტების სახით. ეს უკანასკნელი მიკროტუბულების მონაწილეობით მიმართულია უჯრედის ზედაპირზე. ვეზიკულის მემბრანა ჩადგმულია პლაზმალემაში, ხოლო ვეზიკულის შიგთავსი უჯრედის გარეთაა (ნახ.) ვეზიკულის შერწყმა პლაზმალემასთან შეიძლება მოხდეს დამატებითი სიგნალების გარეშე. ამ ეგზოციტოზს ე.წ კონსტიტუციური.ასე გამოიყოფა უჯრედიდან საკუთარი მეტაბოლიზმის პროდუქტების უმეტესი ნაწილი. თუმცა, რიგი უჯრედები შექმნილია სპეციალური ნაერთების სინთეზისთვის - საიდუმლოებები, რომლებიც გამოიყენება სხეულის სხვა ნაწილებში. იმისათვის, რომ სატრანსპორტო ვეზიკულა საიდუმლოებით შეერწყას პლაზმურ მემბრანას, საჭიროა გარე სიგნალები. მხოლოდ ამის შემდეგ მოხდება შერწყმა და საიდუმლო გათავისუფლდება. ამ ეგზოციტოზს ე.წ რეგულირდება... სასიგნალო მოლეკულებს, რომლებიც ხელს უწყობენ სეკრეციის გამოყოფას, ე.წ ლიბერინები (განთავისუფლების ფაქტორები),და აღმოფხვრის თავიდან აცილება - სტატინები.
რეცეპტორის ფუნქციები.
ისინი ძირითადად უზრუნველყოფილნი არიან გლიკოპროტეინებით, რომლებიც მდებარეობს პლაზმალემის ზედაპირზე და შეუძლიათ მათ ლიგანდებთან შეკავშირება. ლიგანდი შეესაბამება მის რეცეპტორს, როგორც საკეტის გასაღები. ლიგანდის რეცეპტორთან შეერთება იწვევს პოლიპეპტიდის კონფორმაციის ცვლილებას. ტრანსმემბრანული ცილის ასეთი ცვლილებით მყარდება კომუნიკაცია უჯრედშორის და უჯრედშიდა გარემოს შორის.
რეცეპტორების ტიპები.
პროტეინის იონურ არხებთან დაკავშირებული რეცეპტორები. ისინი ურთიერთქმედებენ სასიგნალო მოლეკულასთან, რომელიც დროებით ხსნის ან ხურავს არხს იონების გავლისთვის. (მაგალითად, აცეტილქოლინის ნეიროტრანსმიტერის რეცეპტორი არის ცილა, რომელიც შედგება 5 ქვედანაყოფისგან, რომლებიც ქმნიან იონურ არხს. აცეტილქოლინის არარსებობის შემთხვევაში, არხი იკეტება და მიმაგრების შემდეგ იხსნება და საშუალებას აძლევს ნატრიუმის იონებს გაიარონ).
კატალიზური რეცეპტორები. ისინი შედგება უჯრედგარე ნაწილისგან (თავად რეცეპტორი) და უჯრედშიდა ციტოპლაზმური ნაწილისგან, რომელიც ფუნქციონირებს როგორც ფერმენტ პროლინ კინაზა (მაგალითად, ზრდის ჰორმონის რეცეპტორები).
რეცეპტორები, რომლებიც დაკავშირებულია G- პროტეინებთან. ეს არის ტრანსმემბრანული ცილები, რომლებიც შედგება რეცეპტორისგან, რომელიც ურთიერთქმედებს ლიგანდთან და G- პროტეინთან (გუანოზინტრიფოსფატთან დაკავშირებული მარეგულირებელი ცილა), რომელიც გადასცემს სიგნალს მემბრანასთან დაკავშირებულ ფერმენტზე (ადენილატ ციკლაზა) ან იონურ არხზე. შედეგად, ციკლური AMP ან კალციუმის იონები გააქტიურებულია. (ასე მუშაობს ადენილატციკლაზას სისტემა. მაგალითად, ღვიძლის უჯრედებში არის რეცეპტორი ჰორმონის ინსულინის. რეცეპტორის ზეუჯრედული ნაწილი აკავშირებს ინსულინს. ეს იწვევს უჯრედშიდა ნაწილის - ფერმენტ ადენილატ ციკლაზას გააქტიურებას. იგი ასინთეზებს ციკლურ AMP-ს ATP-დან, რომელიც არეგულირებს სხვადასხვა უჯრედშიდა პროცესების სიჩქარეს, რაც იწვევს ამ ან სხვა მეტაბოლური ფერმენტების აქტივაციას ან ინჰიბირებას).
რეცეპტორები, რომლებიც აღიქვამენ ფიზიკურ ფაქტორებს. მაგალითად, ფოტორეცეპტორი ცილა როდოპსინი. სინათლის შთანთქმისას ის იცვლის თავის კონფორმაციას და აღაგზნებს ნერვულ იმპულსს.
იგი შედგება ბილიპიდური შრისგან, რომლის ლიპიდები მკაცრად ორიენტირებულია - ლიპიდების ჰიდროფობიური ნაწილი (კუდი) მიმართულია ფენის შიგნით, ხოლო ჰიდროფილური ნაწილი (თავი) გარეთაა. გარდა ლიპიდებისა, პლაზმური მემბრანის აგებაში მონაწილეობს მემბრანის სამი ტიპი: პერიფერიული, ინტეგრალური და ნახევრად ინტეგრალური.
მემბრანების შესწავლის ერთ-ერთი სფერო ამჟამად არის მემბრანების შემადგენელი სხვადასხვა სტრუქტურული და მარეგულირებელი ლიპიდების და ცალკეული ინტეგრალური და ნახევრად ინტეგრალური ცილების თვისებების დეტალური შესწავლა.
მემბრანის ინტეგრალური ცილები
თავად მემბრანის ორგანიზებაში მთავარ როლს ასრულებენ ინტეგრალური და ნახევრად ინტეგრალური პროტეინები, რომლებსაც აქვთ გლობულური სტრუქტურა და ლიპიდურ ფაზას უკავშირდება ჰიდროფილურ - ჰიდროფობიური ურთიერთქმედებით. ინტეგრალური ცილების გლობულები შეაღწევს მემბრანის მთელ სისქეს და მათი ჰიდროფობიური ნაწილი მდებარეობს გლობულის შუაში და ჩაეფლო ლიპიდური ფაზის ჰიდროფობიურ ზონაში.
ნახევრად ინტეგრალური მემბრანის ცილები
ნახევრად ინტეგრალურ პროტეინებში, ჰიდროფობიური ამინომჟავები კონცენტრირებულია გლობულის ერთ-ერთ პოლუსზე და, შესაბამისად, გლობულები მხოლოდ ნახევრად არის ჩაძირული მემბრანაში, გამოდის გარედან ერთი (გარე ან შიდა) მემბრანის ზედაპირიდან.
მემბრანის ცილების ფუნქციები
პლაზმური მემბრანის ინტეგრალურ და ნახევრად ინტეგრალურ ცილებს ადრე ენიჭებოდათ ორი ფუნქცია: ზოგადი სტრუქტურული და სპეციფიკური. შესაბამისად, მათ შორის გამოიყო სტრუქტურული და ფუნქციური ცილები. თუმცა, მემბრანების ცილოვანი ფრაქციების იზოლირების მეთოდების გაუმჯობესება და ცალკეული ცილების უფრო დეტალური ანალიზი ახლა მიუთითებს უნივერსალური სტრუქტურული ცილების არარსებობაზე ყველა მემბრანისთვის, რომელიც არ ახორციელებს რაიმე სპეციფიკურ ფუნქციას. ამის საპირისპიროდ, მემბრანის ცილები სპეციფიკური ფუნქციებით ძალიან მრავალფეროვანია. ეს არის ცილები, რომლებიც ასრულებენ რეცეპტორების ფუნქციებს, ცილები, რომლებიც არიან სხვადასხვა ნაერთების აქტიური და პასიური მატარებლები, და ბოლოს, ცილები, რომლებიც მრავალი ფერმენტული სისტემის ნაწილია. მასალა საიტიდან
მემბრანის ცილების თვისებები
ყველა ამ ინტეგრალური და ნახევრად ინტეგრალური მემბრანული ცილების საერთო თვისება, რომელიც განსხვავდება არა მხოლოდ ფუნქციური, არამედ ქიმიურადაც, არის მათი ფუნდამენტური უნარი გადაადგილება, "ცურვა" მემბრანის სიბრტყეში თხევადი ლიპიდური ფაზაში. როგორც ზემოთ აღინიშნა, ზოგიერთი უჯრედის პლაზმურ მემბრანებში ასეთი მოძრაობების არსებობა ექსპერიმენტულად დადასტურდა. მაგრამ ეს შორს არის მემბრანის ცილებში გამოვლენილი მოძრაობის ერთადერთი ტიპისაგან. გვერდითი გადაადგილების გარდა, ცალკეულ ინტეგრალურ და ნახევრად ინტეგრალურ ცილებს შეუძლიათ მემბრანის სიბრტყეში ბრუნვა ჰორიზონტალური და თუნდაც ვერტიკალური მიმართულებით, ასევე შეუძლიათ შეცვალონ მოლეკულის ჩაძირვის ხარისხი ლიპიდურ ფაზაში.
ოპსინი.ცილის გლობულების ყველა ეს მრავალფეროვანი და რთული მოძრაობა განსაკუთრებით კარგად არის ასახული ოპსინის ცილის მაგალითით, რომელიც სპეციფიკურია ფოტორეცეპტორული უჯრედების მემბრანებისთვის (ნახ. 3). მოგეხსენებათ, ოპსინი სიბნელეში ასოცირდება კაროტინოიდ ბადურასთან, რომელიც შეიცავს ორმაგ ცისუს კავშირს; ბადურის და ოპსინის კომპლექსი ქმნის როდოპსინს, ანუ ვიზუალურ მეწამულს. როდოპსინის მოლეკულას შეუძლია გვერდითი მოძრაობა და ბრუნვა მემბრანის ჰორიზონტალურ სიბრტყეში (ნახ. 3, A). სინათლის ზემოქმედებისას, ბადურა განიცდის ფოტოიზომერიზაციას და ტრანსფორმირდება ტრანს ფორმაში. ამ შემთხვევაში იცვლება ბადურას კონფორმაცია და ის გამოიყოფა ოპსინისგან, რაც თავის მხრივ ცვლის ბრუნვის სიბრტყეს ჰორიზონტალურიდან ვერტიკალურზე (ნახ. 3, B). ასეთი გარდაქმნების შედეგია იონების მემბრანის გამტარიანობის ცვლილება, რაც იწვევს ნერვული იმპულსის გაჩენას.
საინტერესოა, რომ სინათლის ენერგიით გამოწვეული ოპსინის გლობულების კონფორმაციის ცვლილებები არა მხოლოდ ნერვული იმპულსების გამომუშავებას ემსახურება, როგორც ეს ხდება თვალის ბადურის უჯრედებში, არამედ არის უმარტივესი ფოტოსინთეზური სისტემა, რომელიც გვხვდება სპეციალურ მეწამულ ბაქტერიებში.
ბიოლოგიური მემბრანები ქმნიან უჯრედის სტრუქტურული ორგანიზაციის საფუძველს. პლაზმური მემბრანა (პლაზმალემა) არის მემბრანა, რომელიც აკრავს ცოცხალი უჯრედის ციტოპლაზმას. მემბრანები შედგება ლიპიდებისა და ცილებისგან. ლიპიდები (ძირითადად ფოსფოლიპიდები) ქმნიან ორმაგ ფენას, რომელშიც მოლეკულების ჰიდროფობიური „კუდები“ მიმართულია მემბრანის შიგნითაკენ, ხოლო ჰიდროფილური – მისი ზედაპირებისკენ. ცილის მოლეკულები შეიძლება განთავსდეს მემბრანის გარე და შიდა ზედაპირზე, მათ შეუძლიათ ნაწილობრივ ჩაიძირონ ლიპიდურ შრეში ან შეაღწიონ მასში და მეშვეობით. ჩაძირული მემბრანის ცილების უმეტესობა ფერმენტებია. ეს არის პლაზმური მემბრანის სტრუქტურის სითხე-ძვლოვან-მოზაიკური მოდელი. ცილის და ლიპიდური მოლეკულები მობილურია, რაც უზრუნველყოფს მემბრანის დინამიზმს. მემბრანები ასევე შეიცავს ნახშირწყლებს გლიკოლიპიდების და გლიკოპროტეინების (გლიკოკალიქსის) სახით, რომლებიც მდებარეობს მემბრანის გარე ზედაპირზე. თითოეული უჯრედის მემბრანის ზედაპირზე ცილებისა და ნახშირწყლების ნაკრები სპეციფიკურია და წარმოადგენს უჯრედის ტიპის ერთგვარ მაჩვენებელს.
მემბრანის ფუნქციები:
- გამოყოფა. იგი შედგება ბარიერის ფორმირებაში უჯრედის შიდა შიგთავსსა და გარე გარემოს შორის.
- ციტოპლაზმასა და გარე გარემოს შორის ნივთიერებების გაცვლის უზრუნველყოფა. წყალი, იონები, არაორგანული და ორგანული მოლეკულები შედის უჯრედში (სატრანსპორტო ფუნქცია). უჯრედში წარმოქმნილი პროდუქტები (სეკრეტორული ფუნქცია) გამოიყოფა გარე გარემოში.
- ტრანსპორტი. მემბრანის გასწვრივ ტრანსპორტირება შეიძლება განხორციელდეს სხვადასხვა გზით. პასიური ტრანსპორტი ხორციელდება ენერგიის მოხმარების გარეშე, მარტივი დიფუზიით, ოსმოსით ან გაადვილებული დიფუზიით გადამზიდი ცილების დახმარებით. აქტიური ტრანსპორტი ხდება გადამზიდავი ცილების დახმარებით და მას სჭირდება ენერგია (მაგალითად, ნატრიუმ-კალიუმის ტუმბო). მასალა საიტიდან
ბიოპოლიმერების დიდი მოლეკულები უჯრედში ხვდება ენდოციტოზის შედეგად. იგი იყოფა ფაგოციტოზად და პინოციტოზად. ფაგოციტოზი არის უჯრედის მიერ დიდი ნაწილაკების დაჭერა და შეწოვა. ფენომენი პირველად აღწერა I.I. მეჩნიკოვი. თავდაპირველად, ნივთიერებები ეკვრის პლაზმურ მემბრანას, სპეციფიკურ რეცეპტორულ ცილებს, შემდეგ მემბრანა იშლება და ქმნის დეპრესიას.
წარმოიქმნება საჭმლის მომნელებელი ვაკუოლი. ის ითვისებს უჯრედში შესულ ნივთიერებებს. ადამიანებში და ცხოველებში ლეიკოციტებს შეუძლიათ ფაგოციტოზი. სისხლის თეთრი უჯრედები შთანთქავს ბაქტერიებს და სხვა ნაწილაკებს.
პინოციტოზი არის თხევადი წვეთების დაჭერა და შთანთქმის პროცესი მასში გახსნილი ნივთიერებებით. ნივთიერებები ეკვრის მემბრანის პროტეინებს (რეცეპტორებს), ხოლო ხსნარის წვეთი გარშემორტყმულია მემბრანით, რომელიც ქმნის ვაკუოლს. პინოციტოზი და ფაგოციტოზი წარმოიქმნება ATP ენერგიის ხარჯვით.
- სეკრეტორული. სეკრეცია - უჯრედის მიერ უჯრედში სინთეზირებული ნივთიერებების გათავისუფლება გარე გარემოში. ჰორმონები, პოლისაქარიდები, ცილები, ცხიმის წვეთები ჩასმულია მემბრანით შემოსაზღვრულ ვეზიკულებში და მიდიან პლაზმალემაში. მემბრანები ერწყმის და ვეზიკულის შიგთავსი გამოიყოფა უჯრედის მიმდებარე გარემოში.
- უჯრედების შეერთება ქსოვილში (დაკეცილი გამონაზარდების გამო).
- რეცეპტორი. მემბრანები შეიცავს რეცეპტორების დიდ რაოდენობას – სპეციალურ ცილებს, რომელთა როლი არის სიგნალების გადაცემა გარედან უჯრედის შიგნით.
1. ბარიერი- უზრუნველყოფს გარემოსთან დარეგულირებულ, შერჩევით, პასიურ და აქტიურ მეტაბოლიზმს.
უჯრედის მემბრანებს აქვთ შერჩევითი გამტარიანობა: გლუკოზა, ამინომჟავები, ცხიმოვანი მჟავები, გლიცეროლი და იონები ნელ-ნელა დიფუზირდება მათში, თავად გარსები აქტიურად არეგულირებენ ამ პროცესს - ზოგიერთი ნივთიერება ნებადართულია, ზოგი კი არა.
2. ტრანსპორტი- მემბრანის მეშვეობით ნივთიერებები უჯრედში და გარეთ გადადის. მემბრანების მეშვეობით ტრანსპორტი უზრუნველყოფს: საკვები ნივთიერებების მიწოდებას, მეტაბოლური პროდუქტების მოცილებას, სხვადასხვა ნივთიერებების სეკრეციას, იონური გრადიენტების შექმნას, უჯრედში შესაბამისი pH და იონური კონცენტრაციის შენარჩუნებას, რაც აუცილებელია უჯრედული ფერმენტების მუშაობისთვის.
არსებობს უჯრედში ნივთიერებების შეყვანის ან უჯრედიდან გარედან მათი ამოღების ოთხი ძირითადი მექანიზმი:
ა) პასიური (დიფუზია, ოსმოზი) (არ საჭიროებს ენერგიის მოხმარებას)
დიფუზია
ერთი ნივთიერების მოლეკულების ან ატომების გავრცელება მოლეკულებს ან მეორის ატომებს შორის, რაც იწვევს მათი კონცენტრაციების სპონტანურ გათანაბრებას მთელ დაკავებულ მოცულობაში. ზოგიერთ სიტუაციაში, ერთ-ერთ ნივთიერებას უკვე აქვს გათანაბრებული კონცენტრაცია და ისინი საუბრობენ ერთი ნივთიერების მეორეში გავრცელებაზე. ამ შემთხვევაში მატერიის გადატანა ხდება მაღალი კონცენტრაციის ფართობიდან დაბალი კონცენტრაციის ზონაში (კონცენტრაციის გრადიენტის ვექტორის გასწვრივ (ნახ. 2.4).
ბრინჯი. 2.4. დიფუზიის პროცესის დიაგრამა
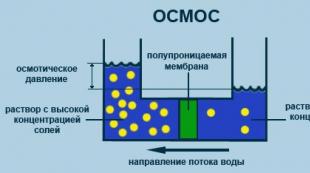
ოსმოზი
ცალმხრივი დიფუზიის პროცესი გამხსნელის მოლეკულების ნახევრად გამტარი მემბრანის მეშვეობით გახსნილი ნივთიერების უფრო მაღალი კონცენტრაციისკენ გახსნილი ნივთიერების დაბალი კონცენტრაციის მოცულობიდან. (ნახ. 2.5).

ბრინჯი. 2.5. ოსმოსის პროცესის დიაგრამა
ბ) აქტიური ტრანსპორტი (მოითხოვს ენერგიის მოხმარებას)
ნატრიუმ-კალიუმის ტუმბო- ნატრიუმის იონების (უჯრედიდან) და კალიუმის იონების (უჯრედში) აქტიური კონიუგირებული ტრანსმემბრანული ტრანსპორტირების მექანიზმი, რომელიც უზრუნველყოფს კონცენტრაციის გრადიენტს და ტრანსმემბრანული პოტენციალის განსხვავებას. ეს უკანასკნელი ემსახურება უჯრედებისა და ორგანოების მრავალი ფუნქციის საფუძველს: ჯირკვლის უჯრედების სეკრეციას, კუნთების შეკუმშვას, ნერვული იმპულსების გამტარობას და ა.შ. (ნახ. 2.6).

ბრინჯი. 2.6. კალიუმ-ნატრიუმის ტუმბოს სქემა
პირველ ეტაპზე ფერმენტი Na + / K + -ATPase ამაგრებს სამ Na + იონს მემბრანის შიდა მხრიდან. ეს იონები ცვლის ატფ-აზას აქტიური ცენტრის კონფორმაციას. ამის შემდეგ ფერმენტს შეუძლია ერთი ATP მოლეკულის ჰიდროლიზება. ჰიდროლიზის შემდეგ გამოთავისუფლებული ენერგია იხარჯება მატარებლის კონფორმაციის შეცვლაზე, რის გამოც სამი Na + იონი და PO 4 3– იონი (ფოსფატი) ჩნდება მემბრანის გარე მხარეს. აქ Na + იონები იყოფა და PO 4 3– ჩანაცვლებულია ორი K + იონით. ამის შემდეგ ფერმენტი უბრუნდება თავდაპირველ კონფორმაციას და K + იონები ჩნდება მემბრანის შიდა მხარეს. აქ K + იონები იყოფა და გადამზიდავი კვლავ მზად არის სამუშაოდ.
შედეგად, უჯრედგარე გარემოში იქმნება Na + იონების მაღალი კონცენტრაცია, ხოლო უჯრედის შიგნით იქმნება K + მაღალი კონცენტრაცია. კონცენტრაციის ეს განსხვავება გამოიყენება უჯრედებში ნერვული იმპულსის გატარების დროს.
გ) ენდოციტოზი (ფაგოციტოზი, პინოციტოზი)
ფაგოციტოზი(უჯრედის მიერ ჭამა) - უჯრედის მიერ მყარი საგნების შეწოვის პროცესი, როგორიცაა ევკარიოტული უჯრედები, ბაქტერიები, ვირუსები, მკვდარი უჯრედების ნაშთები და ა.შ. შთანთქმის ობიექტის გარშემო წარმოიქმნება დიდი უჯრედშიდა ვაკუოლი (ფაგოსომა). ფაგოსომების ზომა არის 250 ნმ და მეტი. ფაგოსომის პირველად ლიზოსომასთან შერწყმის შედეგად წარმოიქმნება მეორადი ლიზოსომა. მჟავე გარემოში ჰიდროლიზური ფერმენტები ანადგურებს მაკრომოლეკულებს მეორად ლიზოსომაში. დაშლის პროდუქტები (ამინომჟავები, მონოსაქარიდები და სხვა სასარგებლო ნივთიერებები) შემდეგ ტრანსპორტირდება ლიზოსომური მემბრანის მეშვეობით უჯრედის ციტოპლაზმაში. ფაგოციტოზი ძალიან გავრცელებულია. მაღალ ორგანიზებულ ცხოველებსა და ადამიანებში ფაგოციტოზის პროცესი დამცავ როლს ასრულებს. ლეიკოციტების და მაკროფაგების ფაგოციტურ აქტივობას დიდი მნიშვნელობა აქვს ორგანიზმის დაცვაში პათოგენური მიკრობებისა და მასში შემავალი სხვა არასასურველი ნაწილაკებისგან. ფაგოციტოზი პირველად აღწერა რუსმა მეცნიერმა ი.ი.მეჩნიკოვმა (ნახ. 2.7)
პინოციტოზი(უჯრედის მიერ დალევა) - ხსნადი ნივთიერებების, მათ შორის დიდი მოლეკულების (ცილები, პოლისაქარიდები და ა.შ.) შემცველი გარემოდან თხევადი ფაზის უჯრედის მიერ შეწოვის პროცესი. პინოციტოზის დროს უჯრედის შიგნით არსებული მემბრანისგან იშლება მცირე ზომის ვეზიკულები - ენდოსომები. ისინი უფრო მცირეა ვიდრე ფაგოსომები (მათი ზომა 150 ნმ-მდეა) და ჩვეულებრივ არ შეიცავს დიდ ნაწილაკებს. ენდოსომის წარმოქმნის შემდეგ პირველადი ლიზოსომა უახლოვდება მას და ეს ორი მემბრანული ვეზიკულა ერწყმის ერთმანეთს. მიღებულ ორგანელას მეორადი ლიზოსომა ეწოდება. პინოციტოზის პროცესი მუდმივად ხორციელდება ყველა ევკარიოტული უჯრედის მიერ. (ნახ. 7)
რეცეპტორებით გამოწვეული ენდოციტოზი - აქტიური სპეციფიური პროცესი, რომლის დროსაც უჯრედის მემბრანა იშლება უჯრედში, ქმნის კიდეებს ორმოებს. მოპირკეთებული ფოსოს უჯრედშიდა მხარე შეიცავს ადაპტაციურ ცილებს. მაკრომოლეკულები, რომლებიც აკავშირებენ უჯრედის ზედაპირზე არსებულ სპეციფიკურ რეცეპტორებს, შიგნით გადიან ბევრად უფრო მაღალი სიჩქარით, ვიდრე ნივთიერებები, რომლებიც უჯრედებში შედიან პინოციტოზის გზით.

ბრინჯი. 2.7. ენდოციტოზი
დ) ეგზოციტოზი (უარყოფითი ფაგოციტოზი და პინოციტოზი)
უჯრედული პროცესი, რომლის დროსაც უჯრედშიდა ვეზიკულები (მემბრანული ვეზიკულები) ერწყმის გარე უჯრედის მემბრანას. ეგზოციტოზის დროს სეკრეტორული ვეზიკულების შიგთავსი (ეგზოციტური ვეზიკულები) გამოიყოფა გარეთ და მათი მემბრანა ერწყმის უჯრედის მემბრანას. ამ მეთოდით უჯრედიდან გამოიყოფა თითქმის ყველა მაკრომოლეკულური ნაერთი (ცილები, პეპტიდური ჰორმონები და სხვ.). (ნახ. 2.8)

ბრინჯი. 2.8. ეგზოციტოზის სქემა
3. ბიოპოტენციალების წარმოქმნა და გატარება- მემბრანის დახმარებით უჯრედში იონების მუდმივი კონცენტრაცია შენარჩუნებულია: უჯრედის შიგნით K + იონის კონცენტრაცია გაცილებით მაღალია, ვიდრე გარეთ, ხოლო Na +-ის კონცენტრაცია გაცილებით დაბალია, რაც ძალიან მნიშვნელოვანია, ვინაიდან ეს უზრუნველყოფს მემბრანაზე პოტენციური სხვაობის შენარჩუნებას და ნერვული იმპულსის წარმოქმნას.
4. მექანიკური- უზრუნველყოფს უჯრედის ავტონომიას, მის უჯრედშიდა სტრუქტურებს, ასევე კავშირს სხვა უჯრედებთან (ქსოვილებში).
5. ენერგია- ქლოროპლასტებში ფოტოსინთეზის და მიტოქონდრიებში უჯრედული სუნთქვის დროს მათ მემბრანებში მოქმედებს ენერგიის გადაცემის სისტემები, რომელშიც ასევე მონაწილეობენ ცილები;
6. რეცეპტორი- მემბრანის ზოგიერთი ცილა არის რეცეპტორები (მოლეკულები, რომელთა მეშვეობითაც უჯრედი აღიქვამს გარკვეულ სიგნალებს).
7. ფერმენტული- მემბრანის ცილები ხშირად ფერმენტებია. მაგალითად, ნაწლავის ეპითელური უჯრედების პლაზმური გარსები შეიცავს საჭმლის მომნელებელ ფერმენტებს.
8. მატრიცა- უზრუნველყოფს მემბრანის ცილების გარკვეულ ურთიერთგანლაგებას და ორიენტაციას, მათ ოპტიმალურ ურთიერთქმედებას;
9. გალიის მარკირება- მემბრანაზე არის ანტიგენები, რომლებიც მოქმედებენ როგორც მარკერები - "ეტიკეტები", რომლებიც საშუალებას გაძლევთ ამოიცნოთ უჯრედი. ეს არის გლიკოპროტეინები (ანუ ცილები, მათზე მიმაგრებული განშტოებული ოლიგოსაქარიდული გვერდითი ჯაჭვებით), რომლებიც ასრულებენ „ანტენების“ როლს. მარკერების დახმარებით უჯრედებს შეუძლიათ სხვა უჯრედების ამოცნობა და მათთან ერთად მოქმედება, მაგალითად, ორგანოებისა და ქსოვილების ფორმირებისას. ის ასევე საშუალებას აძლევს იმუნურ სისტემას ამოიცნოს უცხო ანტიგენები.
ფიჭური ჩანართები
უჯრედული ჩანართები მოიცავს ნახშირწყლებს, ცხიმებს და ცილებს. ყველა ეს ნივთიერება გროვდება უჯრედის ციტოპლაზმაში სხვადასხვა ზომისა და ფორმის წვეთებისა და მარცვლის სახით. ისინი პერიოდულად სინთეზირდება უჯრედში და გამოიყენება მეტაბოლურ პროცესში.
ციტოპლაზმა
ეს არის ცოცხალი უჯრედის (პროტოპლასტის) ნაწილი პლაზმური მემბრანისა და ბირთვის გარეშე. ციტოპლაზმის შემადგენლობა მოიცავს: ციტოპლაზმურ მატრიქსს, ციტოჩონჩხს, ორგანელებს და ჩანართებს (ზოგჯერ ვაკუოლების ჩანართები და შიგთავსი არ კლასიფიცირდება ციტოპლაზმის ცოცხალ ნივთიერებად). გარე გარემოდან პლაზმური მემბრანით გამოყოფილი ციტოპლაზმა არის უჯრედების შიდა ნახევრად თხევადი გარემო. ბირთვი და სხვადასხვა ორგანელები განლაგებულია ევკარიოტული უჯრედების ციტოპლაზმაში. იგი ასევე შეიცავს მრავალფეროვან ჩანართებს - უჯრედული აქტივობის პროდუქტებს, ვაკუოლებს, ასევე უმცირეს მილაკებსა და ძაფებს, რომლებიც ქმნიან უჯრედის ჩონჩხს. ციტოპლაზმის ძირითადი ნივთიერების შემადგენლობაში ჭარბობს ცილები.
ციტოპლაზმური ფუნქციები
1) მასში მიმდინარეობს ძირითადი მეტაბოლური პროცესები.
2) აერთიანებს ბირთვს და ყველა ორგანელას ერთ მთლიანობაში, უზრუნველყოფს მათ ურთიერთქმედებას.
3) მობილურობა, გაღიზიანებადობა, მეტაბოლიზმი და რეპროდუქცია.
მობილურობა მოდის სხვადასხვა ფორმით:
უჯრედის ციტოპლაზმის უჯრედშიდა მოძრაობა.
ამეების ფორმის მოძრაობა. მოძრაობის ეს ფორმა გამოიხატება ციტოპლაზმის მიერ ფსევდოპოდიის წარმოქმნით ამა თუ იმ სტიმულისკენ ან მისგან მოშორებით. მოძრაობის ეს ფორმა თანდაყოლილია ამებაში, სისხლის ლეიკოციტებში, ასევე ქსოვილის ზოგიერთ უჯრედში.
მბჟუტავი მოძრაობა. იგი ვლინდება პაწაწინა პროტოპლაზმური წარმონაქმნების - წამწამებისა და ფლაგელების (ცილატები, მრავალუჯრედიანი ცხოველების ეპითელური უჯრედები, სპერმატოზოიდები და ა.შ.) ცემის სახით.
კონტრაქტის მოძრაობა. იგი უზრუნველყოფილია ციტოპლაზმაში სპეციალური ორგანოიდური მიოფიბრილების არსებობის გამო, რომელთა შემცირება ან გახანგრძლივება ხელს უწყობს უჯრედის შეკუმშვას და მოდუნებას. შეკუმშვის უნარი ყველაზე მეტად განვითარებულია კუნთოვან უჯრედებში.
გაღიზიანება გამოიხატება უჯრედების უნარში, რეაგირება მოახდინონ გაღიზიანებაზე მეტაბოლიზმისა და ენერგიის შეცვლით.
ციტოჩონჩხი
ევკარიოტული უჯრედის ერთ-ერთი გამორჩეული მახასიათებელია მის ციტოპლაზმაში ჩონჩხის წარმონაქმნების არსებობა მიკროტუბულებისა და ცილის ბოჭკოების ჩალიჩების სახით. ციტოჩონჩხის ელემენტები, რომლებიც მჭიდრო კავშირშია გარე ციტოპლაზმურ მემბრანასთან და ბირთვულ გარსთან, ქმნიან კომპლექსურ ქსოვილებს ციტოპლაზმაში.
ციტოჩონჩხი იქმნება მიკროტუბულებით, მიკროფილამენტებით და მიკროტრაბეკულური სისტემით. ციტოჩონჩხი განსაზღვრავს უჯრედის ფორმას, მონაწილეობს უჯრედის მოძრაობაში, თავად უჯრედის გაყოფასა და მოძრაობაში, ორგანელების უჯრედშიდა ტრანსპორტში.
მიკროტუბულებიშეიცავს ყველა ეუკარიოტულ უჯრედს და წარმოადგენს ღრუ განშტოებული ცილინდრებს, რომელთა დიამეტრი არ აღემატება 30 ნმ, ხოლო კედლის სისქე 5 ნმ. მათი სიგრძე შეიძლება მიაღწიოს რამდენიმე მიკრომეტრს. ადვილად იშლება და ხელახლა იკრიბება. მიკროტუბულების კედელი ძირითადად შედგება სპირალურად დაკეცილი ტუბულინის ცილის ქვედანაყოფებისგან (ნახ. 2.09)
მიკროტუბულების ფუნქციები:
1) შეასრულოს დამხმარე ფუნქცია;
2) ჩამოყალიბდეს დაშლის spindle; უზრუნველყოს ქრომოსომების განსხვავება უჯრედის პოლუსებთან; პასუხისმგებელნი არიან უჯრედის ორგანელების მოძრაობაზე;
3) მონაწილეობა მიიღოს უჯრედშიდა ტრანსპორტში, სეკრეციაში, უჯრედის კედლის ფორმირებაში;
4) არის წამწამების, დროშების, ბაზალური სხეულების და ცენტრიოლების სტრუქტურული კომპონენტი.
მიკროფილამენტებიწარმოდგენილია ძაფებით 6 ნმ დიამეტრით, რომელიც შედგება აქტინის ცილისგან, რომელიც ახლოს არის კუნთების აქტინთან. აქტინი შეადგენს უჯრედის მთლიანი ცილის 10-15%-ს. ცხოველური უჯრედების უმეტესობაში აქტინის ძაფებისა და ასოცირებული ცილების მკვრივი ქსელი იქმნება თავად პლაზმური მემბრანის ქვეშ.
აქტინის გარდა, უჯრედში ასევე გვხვდება მიოზინის ძაფები. თუმცა მათი რიცხვი გაცილებით ნაკლებია. აქტინისა და მიოზინის ურთიერთქმედების გამო ხდება კუნთების შეკუმშვა. მიკროფილამენტები დაკავშირებულია მთელი უჯრედის ან მასში არსებული მისი ცალკეული სტრუქტურების მოძრაობასთან. ზოგ შემთხვევაში მოძრაობა უზრუნველყოფილია მხოლოდ აქტინის ძაფებით, ზოგ შემთხვევაში - აქტინით მიოსინთან ერთად.
მიკროფილამენტების ფუნქციები
1) მექანიკური სიმტკიცე
2) საშუალებას აძლევს უჯრედს შეცვალოს ფორმა და გადაადგილდეს.

ბრინჯი. 2.09. ციტოჩონჩხი
ორგანელები (ან ორგანელები)
იყოფა არამემბრანული, ერთმემბრანული და ორმემბრანული.
TO არამემბრანული ორგანელებიეუკარიოტული უჯრედები მოიცავს ორგანელებს, რომლებსაც არ აქვთ საკუთარი დახურული მემბრანა, კერძოდ: რიბოზომებიდა ტუბულინის მიკროტუბულების საფუძველზე აგებული ორგანელები - უჯრედის ცენტრი (ცენტრიოლები)და მოძრაობის ორგანელები (ფლაგელა და ცილიები).ერთუჯრედიანი ორგანიზმების უმეტესობის უჯრედებში და უმაღლესი (ხმელეთის) მცენარეების აბსოლუტურ უმრავლესობაში ცენტრიოლები არ არის.
TO ერთმემბრანიანი ორგანელებიეხება: ენდოპლაზმური ბადე, გოლჯის აპარატი, ლიზოსომები, პეროქსიზომები, სფეროსომები, ვაკუოლები და სხვა.ყველა ერთმემბრანიანი ორგანელი ერთმანეთთან არის დაკავშირებული ერთ უჯრედულ სისტემაში. მცენარეთა უჯრედებს აქვთ სპეციალური ლიზოსომები, ცხოველურ უჯრედებს აქვთ სპეციალური ვაკუოლები: საჭმლის მომნელებელი, ექსკრეტორული, კონტრაქტული, ფაგოციტური, აუტოფაგოციტური და ა.შ.
TO ორმემბრანიანი ორგანელებიეხება მიტოქონდრია და პლასტიდები.
არამემბრანული ორგანელები
ა) რიბოსომები- ყველა ორგანიზმის უჯრედებში ნაპოვნი ორგანელები. ეს არის პატარა ორგანელები, რომლებიც წარმოდგენილია გლობული ნაწილაკებით, რომელთა დიამეტრი დაახლოებით 20 ნმ. რიბოსომები შედგება არათანაბარი ზომის ორი ქვედანაყოფისგან - დიდი და პატარა. რიბოსომები მოიცავს ცილებს და რიბოსომურ რნმ-ს (rRNA). არსებობს რიბოზომების ორი ძირითადი ტიპი: ევკარიოტული (80S) და პროკარიოტული (70S).
უჯრედში ლოკალიზაციის მიხედვით, ციტოპლაზმაში განლაგებულია თავისუფალი რიბოსომები, რომლებიც სინთეზირებენ ცილებს და მიმაგრებულ რიბოზომებს - რიბოზომები, რომლებიც დაკავშირებულია დიდი ქვედანაყოფებით EPR მემბრანების გარე ზედაპირთან, სინთეზირებს ცილებს, რომლებიც შედიან გოლგის კომპლექსში და შემდეგ გამოიყოფა. უჯრედი. ცილის ბიოსინთეზის დროს რიბოსომები შეიძლება გაერთიანდეს კომპლექსებად - პოლირიბოსომებად (პოლისომებად).
ევკარიოტული რიბოსომები წარმოიქმნება ბირთვში. პირველ რიგში, rRNA სინთეზირდება ბირთვულ დნმ-ზე, რომელიც შემდეგ იფარება ციტოპლაზმიდან გამომავალი რიბოსომური ცილებით, იშლება საჭირო ზომამდე და ქმნის რიბოსომის ქვედანაყოფებს. ბირთვში არ არის სრულად ჩამოყალიბებული რიბოსომები. ქვედანაყოფების გაერთიანება მთელ რიბოსომაში ხდება ციტოპლაზმაში, ჩვეულებრივ ცილის ბიოსინთეზის დროს.
რიბოსომები გვხვდება ყველა ორგანიზმის უჯრედში. თითოეული შედგება ორი ნაწილაკისგან, პატარა და დიდი. რიბოსომები შეიცავს ცილებს და რნმ-ს.
ფუნქციები
ცილის სინთეზი.
სინთეზირებული ცილები ჯერ ენდოპლაზმური ბადის არხებსა და ღრუებში გროვდება, შემდეგ კი უჯრედის ორგანელებსა და ნაწილებში გადაიგზავნება. EPS და მის მემბრანებზე განლაგებული რიბოსომები წარმოადგენს ერთ აპარატს ცილების ბიოსინთეზისა და ტრანსპორტირებისთვის. (სურ. 2.10-2.11).

ბრინჯი. 2.10. რიბოსომის სტრუქტურა

ბრინჯი. 2.11. რიბოსომის სტრუქტურა
ბ) უჯრედის ცენტრი (ცენტრიოლები)
ცენტრიოლი არის ცილინდრი (0,3 მკმ სიგრძით და 0,1 მკმ დიამეტრით), რომლის კედელი იქმნება სამი შერწყმული მიკროტუბულის ცხრა ჯგუფისგან (9 სამეული), რომლებიც დაკავშირებულია გარკვეული ინტერვალებით ჯვარედინი კავშირით. ცენტრიოლები ხშირად დაწყვილებულია, სადაც ისინი მართი კუთხით არიან ერთმანეთთან. თუ ცენტრიოლი დევს ცილიუმის ან ფლაგელის ძირში, მაშინ მას ბაზალურ სხეულს უწოდებენ.
თითქმის ყველა ცხოველურ უჯრედს აქვს ცენტრიოლების წყვილი, რომლებიც უჯრედის ცენტრის შუა ელემენტია.
გაყოფამდე ცენტრიოლები საპირისპირო პოლუსებზე გადადის და თითოეულ მათგანთან ასული ცენტრიოლა ჩნდება. უჯრედის სხვადასხვა პოლუსზე მდებარე ცენტრიოლებიდან წარმოიქმნება მიკროტუბულები, რომლებიც იზრდებიან ერთმანეთისკენ.
ფუნქციები
1) ქმნიან მიტოზურ ღეროს, რომელიც ხელს უწყობს გენეტიკური მასალის ერთგვაროვან განაწილებას ქალიშვილ უჯრედებს შორის,
2) წარმოადგენენ ციტოჩონჩხის ორგანიზაციის ცენტრს. spindle-ის ზოგიერთი ძაფები მიმაგრებულია ქრომოსომებზე.
ცენტრიოლები არის ციტოპლაზმის თვითგამრავლებადი ორგანელები. ისინი წარმოიქმნება არსებულის დუბლირების შედეგად. ეს ხდება მაშინ, როდესაც ცენტრიოლები განსხვავდებიან. გაუაზრებელი ცენტრიოლი შეიცავს 9 ცალკეულ მიკროტუბულს; როგორც ჩანს, თითოეული მიკრომილაკი არის მატრიცა მომწიფებული ცენტრიოლისთვის დამახასიათებელი სამეულების შეკრებისთვის (ნახ. 2.12).
ცეტრიოლები გვხვდება ქვედა მცენარეების (წყალმცენარეების) უჯრედებში.

ბრინჯი. 2.12. უჯრედის ცენტრი ცენტრიოლები
ერთმემბრანული ორგანელები
დ) ენდოპლაზმური რეტიკულუმი (EPS)
ციტოპლაზმის მთელი შიდა ზონა ივსება მრავალი მცირე არხებითა და ღრუებით, რომელთა კედლები პლაზმური მემბრანის აგებულებით მსგავსი გარსებია. ეს არხები განშტოდებიან, უერთდებიან ერთმანეთს და ქმნიან ქსელს, რომელსაც ენდოპლაზმურ რეტიკულუმს უწოდებენ. ენდოპლაზმური ბადე სტრუქტურაში ჰეტეროგენულია. ცნობილია მისი ორი ტიპი - მარცვლოვანიდა გლუვი.
მარცვლოვანი ქსელის არხებისა და ღრუების მემბრანებზე ბევრი პატარა მომრგვალებული სხეულია - რიბოსომარომლებიც გარსებს უხეშ იერს ანიჭებენ. გლუვი ენდოპლაზმური ბადის მემბრანები არ ატარებენ რიბოზომებს მათ ზედაპირზე. EPS ასრულებს მრავალ განსხვავებულ ფუნქციას.
ფუნქციები
მარცვლოვანი ენდოპლაზმური ბადის ძირითადი ფუნქციაა ცილის სინთეზში მონაწილეობა, რომელიც ხორციელდება რიბოსომებში. გლუვი ენდოპლაზმური ბადის გარსებზე სინთეზირდება ლიპიდები და ნახშირწყლები. ყველა ეს სინთეზის პროდუქტი გროვდება არხებში და ღრუებში, შემდეგ კი ტრანსპორტირდება უჯრედის სხვადასხვა ორგანელებში, სადაც ისინი მოიხმარენ ან გროვდებიან ციტოპლაზმაში, როგორც უჯრედული ჩანართები. EPS აკავშირებს უჯრედის მთავარ ორგანელებს (ნახ. 2.13).

ბრინჯი. 2.13. ენდოპლაზმური ბადის (EPS) ან რეტიკულუმის სტრუქტურა
ე) გოლჯის აპარატი
ამ ორგანოიდის სტრუქტურა მსგავსია მცენარეთა და ცხოველთა ორგანიზმების უჯრედებში, მიუხედავად მისი ფორმის მრავალფეროვნებისა. მას აქვს მრავალი მნიშვნელოვანი ფუნქცია.
ერთმემბრანული ორგანოიდი. იგი გაშლილი კიდეებით გაბრტყელებული „ცისტერნების“ დასტას წარმოადგენს, რომლებთანაც დაკავშირებულია პატარა ერთგარნიანი ბუშტების სისტემა (გოლგის ბუშტები). გოლჯის ბუშტები ძირითადად კონცენტრირებულია EPS-ის მიმდებარე მხარეს და სტეკების პერიფერიაზე. ითვლება, რომ ისინი გადააქვთ ცილებს და ლიპიდებს გოლჯის აპარატში, რომლის მოლეკულები ცისტერნიდან ცისტერნაში გადაადგილებისას განიცდიან ქიმიურ მოდიფიკაციას.
ყველა ეს ნივთიერება ჯერ გროვდება, ქიმიურად რთულდება, შემდეგ კი დიდი და პატარა ბუშტების სახით შედის ციტოპლაზმაში და ან გამოიყენება თავად უჯრედში სიცოცხლის განმავლობაში, ან გამოიყოფა მისგან და გამოიყენება ორგანიზმში. (სურ. 2.14-2.15).

ბრინჯი. 2.14. გოლჯის აპარატის სტრუქტურა
ფუნქციები:
ცილების, ლიპიდების, ნახშირწყლების მოდიფიკაცია და დაგროვება;
შემომავალი ორგანული ნივთიერებების მემბრანულ ვეზიკულებში (ვეზიკულებში) შეფუთვა;
ლიზოსომების წარმოქმნის ადგილი;
სეკრეტორული ფუნქცია, შესაბამისად, გოლჯის აპარატი კარგად არის განვითარებული სეკრეტორულ უჯრედებში.

ბრინჯი. 2.15. გოლგის კომპლექსი
ე) ლიზოსომები
ისინი პატარა მომრგვალებული სხეულებია. ლიზოსომის შიგნით არის ფერმენტები, რომლებიც ანადგურებენ ცილებს, ცხიმებს, ნახშირწყლებს, ნუკლეინის მჟავებს. ლიზოსომები უახლოვდება ციტოპლაზმაში შესულ საკვებ ნაწილაკს, ერწყმის მას და წარმოიქმნება საჭმლის მომნელებელი ვაკუოლი, რომლის შიგნით არის საკვების ნაწილაკი, რომელიც გარშემორტყმულია ლიზოსომური ფერმენტებით.
ლიზოსომური ფერმენტები სინთეზირდება უხეშ EPS-ზე, გადადის გოლჯის აპარატში, სადაც ისინი მოდიფიცირებულია და შეფუთულია ლიზოსომების მემბრანულ ვეზიკულებში. ლიზოსომა შეიძლება შეიცავდეს 20-დან 60-მდე სხვადასხვა ტიპის ჰიდროლიზურ ფერმენტს. ფერმენტების გამოყენებით ნივთიერებების დაშლას ე.წ ლიზისი.
განასხვავებენ პირველად და მეორად ლიზოსომებს. პირველადი ლიზოსომები ეწოდება გოლჯის აპარატიდან გაჩენილს.
მეორადი ლიზოსომები ეწოდება, რომლებიც წარმოიქმნება პირველადი ლიზოსომების ენდოციტურ ვაკუოლებთან შერწყმის შედეგად. ამ შემთხვევაში ისინი შლიან ნივთიერებებს, რომლებიც უჯრედში შევიდა ფაგოციტოზით ან პინოციტოზით, ამიტომ მათ შეიძლება ეწოდოს საჭმლის მომნელებელი ვაკუოლები.
ლიზოსომების ფუნქციები:
1) ენდოციტოზის დროს უჯრედის მიერ დაჭერილი ნივთიერებების ან ნაწილაკების (ბაქტერიები, სხვა უჯრედები) მონელება;
2) აუტოფაგია - უჯრედისთვის არასაჭირო სტრუქტურების განადგურება, მაგალითად, ძველი ორგანელების ახლით ჩანაცვლებისას, ან თავად უჯრედის შიგნით წარმოქმნილი ცილების და სხვა ნივთიერებების მონელებისას.
3) აუტოლიზი - უჯრედის თვითმონელება, რაც იწვევს მის სიკვდილს (ზოგჯერ ეს პროცესი არ არის პათოლოგიური, მაგრამ თან ახლავს სხეულის განვითარებას ან ზოგიერთი სპეციალიზებული უჯრედის დიფერენციაციას) (სურ. 2.16-2.17).
მაგალითი: როდესაც თათი ბაყაყად იქცევა, კუდის უჯრედებში არსებული ლიზოსომები მას შთანთქავს: კუდი ქრება და ამ პროცესის დროს წარმოქმნილი ნივთიერებები შეიწოვება და გამოიყენება სხეულის სხვა უჯრედების მიერ.

ბრინჯი. 2.16. ლიზოსომების წარმოქმნა

ბრინჯი. 2.17. ლიზოსომების ფუნქციონირება
ზ) პეროქსიზომები
ლიზოსომების აგებულებით მსგავსი ორგანოიდები, 1,5 მიკრონი დიამეტრის მქონე ვეზიკულები ერთგვაროვანი მატრიცით, რომელიც შეიცავს დაახლოებით 50 ფერმენტს.
კატალაზა იწვევს წყალბადის ზეჟანგის 2H 2 O 2 → 2H 2 O + O 2 დაშლას და ხელს უშლის ლიპიდების პეროქსიდაციას
პეროქსიზომები წარმოიქმნება ადრე არსებულიდან კვირტით, ე.ი. მიეკუთვნება თვითგამრავლების ორგანელებს, მიუხედავად იმისა, რომ ისინი არ შეიცავს დნმ-ს. ისინი იზრდებიან მათში ფერმენტების შეყვანის გამო, პეროქსიზომის ფერმენტები წარმოიქმნება უხეშ EPS-ზე და ჰიალოპლაზმაში. (ნახ. 2.18).

ბრინჯი. 2.18. პეროქსისომა (კრისტალური ნუკლეოიდის ცენტრში)
ჰ) ვაკუოლები
ერთმემბრანული ორგანელები. ვაკუოლები არის ორგანული და არაორგანული ნივთიერებების წყალხსნარებით სავსე „კონტეინერები“. EPS და გოლჯის აპარატი ჩართულია ვაკუოლების ფორმირებაში.
ახალგაზრდა მცენარის უჯრედები შეიცავს ბევრ პატარა ვაკუოლს, რომლებიც შემდეგ უჯრედების ზრდისა და დიფერენცირებისას ერწყმის ერთმანეთს და ქმნიან ერთ დიდ ცენტრალურ ვაკუოლს.
ცენტრალურ ვაკუოლს შეუძლია დაიკავოს მომწიფებული უჯრედის მოცულობის 95%, ხოლო ბირთვი და ორგანელები უკან იხევს უჯრედის მემბრანას. მემბრანა, რომელიც ზღუდავს მცენარის ვაკუოლს ე.წ ტონოპლასტი.
სითხეს, რომელიც ავსებს მცენარის ვაკუოლს, ეწოდება უჯრედის წვენი. უჯრედის წვენის შემადგენლობაში შედის წყალში ხსნადი ორგანული და არაორგანული მარილები, მონოსაქარიდები, დისაქარიდები, ამინომჟავები, საბოლოო ან ტოქსიკური მეტაბოლური პროდუქტები (გლიკოზიდები, ალკალოიდები), ზოგიერთი პიგმენტი (ანტოციანინები).
ორგანული ნივთიერებებიდან უფრო ხშირად ინახება შაქარი და ცილები. შაქარი - უფრო ხშირად ხსნარის სახით, ცილები გამოდის EPR ბუშტებისა და გოლჯის აპარატის სახით, რის შემდეგაც ვაკუოლები დეჰიდრატირებულია, გადაიქცევა ალევრონის მარცვლებად.
ცხოველურ უჯრედებში არის მცირე საჭმლის მომნელებელი და აუტოფაგიური ვაკუოლები, რომლებიც მიეკუთვნებიან მეორადი ლიზოსომების ჯგუფს და შეიცავს ჰიდროლიზურ ფერმენტებს. უჯრედულ ცხოველებში ასევე არის კონტრაქტული ვაკუოლები, რომლებიც ასრულებენ ოსმორეგულაციისა და გამოყოფის ფუნქციას.
ფუნქციები
მცენარეებში
1) სითხის დაგროვება და ტურგორის შენარჩუნება,
2) სარეზერვო ნუტრიენტებისა და მინერალური მარილების დაგროვება,
3) ყვავილებისა და ხილის შეღებვა და ამით დამბინძურებლების და ხილისა და თესლის გამავრცელებლების მოზიდვა.
ცხოველებში:
4) საჭმლის მომნელებელი ვაკუოლები - ანადგურებს ორგანულ მაკრომოლეკულებს;
5) კონტრაქტული ვაკუოლები არეგულირებს უჯრედის ოსმოსურ წნევას და აშორებს უჯრედიდან არასაჭირო ნივთიერებებს
6) ფაგოციტური ვაკუოლები წარმოიქმნება ფაგოციტოზის დროს ანტიგენების იმუნური უჯრედების მიერ
7) აუტოფაგოციტური ვაკუოლები წარმოიქმნება ფაგოციტოზის დროს საკუთარი ქსოვილების იმუნური უჯრედების მიერ.
ორმემბრანული ორგანელები (მიტოქონდრია და პლასტიდები)
ეს ორგანელები ნახევრად ავტონომიურია, რადგან მათ აქვთ საკუთარი დნმ და საკუთარი ცილის სინთეზის აპარატი. მიტოქონდრია გვხვდება თითქმის ყველა ევკარიოტულ უჯრედში. პლასტიდები გვხვდება მხოლოდ მცენარეულ უჯრედებში.
ი) მიტოქონდრია
ეს არის უჯრედში მეტაბოლური პროცესების ენერგიის მიწოდების ორგანელები. ჰიალოპლაზმაში, მიტოქონდრია ჩვეულებრივ ნაწილდება დიფუზურად, მაგრამ სპეციალიზებულ უჯრედებში ისინი კონცენტრირდება იმ ადგილებში, სადაც არის ენერგიის უდიდესი საჭიროება. მაგალითად, კუნთოვან უჯრედებში მიტოქონდრიების დიდი რაოდენობა კონცენტრირებულია კონტრაქტული ფიბრილების გასწვრივ, სპერმის ფლაგელუმის გასწვრივ, თირკმლის მილაკების ეპითელიუმში, სინაფსების მიდამოში და ა.შ. მიტოქონდრიების ეს მოწყობა უზრუნველყოფს ATP-ის ნაკლებ დაკარგვას მისი დიფუზიის დროს.
გარეთა გარსი გამოყოფს მიტოქონდრიას ციტოპლაზმისგან, იკეტება თავის თავზე და არ წარმოქმნის ინვაგინაციებს. შიდა მემბრანა ზღუდავს მიტოქონდრიის შიდა შინაარსს - მატრიცას. დამახასიათებელი თვისებაა მრავალი ინვაგინაციის წარმოქმნა - კრისტა, რის გამოც იზრდება შიდა გარსების ფართობი. კრისტალების რაოდენობა და განვითარების ხარისხი დამოკიდებულია ქსოვილის ფუნქციურ აქტივობაზე. მიტოქონდრიებს აქვთ საკუთარი გენეტიკური მასალა (ნახ. 2.19).
მიტოქონდრიული დნმ არის დახურული წრიული ორჯაჭვიანი მოლეკულა, ადამიანის უჯრედებში მას აქვს 16569 ნუკლეოტიდური წყვილის ზომა, რაც დაახლოებით 105-ჯერ უფრო მცირეა ვიდრე ბირთვში ლოკალიზებული დნმ. მიტოქონდრიას აქვს საკუთარი ცილის სინთეზირების სისტემა, ხოლო მიტოქონდრიული mRNA-დან გადათარგმნილი ცილების რაოდენობა შეზღუდულია. მიტოქონდრიულ დნმ-ს არ შეუძლია ყველა მიტოქონდრიული ცილის კოდირება. ცილების უმეტესობა მიტოქონდრიაში იმყოფება ბირთვის გენეტიკური კონტროლის ქვეშ.

ბრინჯი. 2.19. მიტოქონდრიული სტრუქტურა
მიტოქონდრიული ფუნქციები
1) ატფ-ის ფორმირება
2) ცილის სინთეზი
3) მონაწილეობა კონკრეტულ სინთეზებში, მაგალითად, სტეროიდული ჰორმონების (თირკმელზედა ჯირკვლების) სინთეზში.
4) დახარჯულ მიტოქონდრიაში ასევე შეიძლება დაგროვდეს გამოყოფის პროდუქტები, მავნე ნივთიერებები, ე.ი. შეუძლია მიიღოს სხვა უჯრედის ორგანელების ფუნქციები
კ) პლასტიდები
პლასტიდები–ორგანელები, დამახასიათებელი მხოლოდ მცენარეებისთვის.
პლასტიდების სამი ტიპი არსებობს:
1) ქლოროპლასტები(მწვანე პლასტიდები);
2) ქრომოპლასტები(ყვითელი, ნარინჯისფერი ან წითელი პლასტიდები)
3) ლეიკოპლასტები(უფერო პლასტიდები).
ჩვეულებრივ, უჯრედში მხოლოდ ერთი ტიპის პლასტიდები გვხვდება.
ქლოროპლასტები
ეს ორგანელები გვხვდება ფოთლების უჯრედებში და მცენარეთა სხვა მწვანე ორგანოებში, აგრეთვე წყალმცენარეების მრავალფეროვნებაში. უმაღლესი მცენარეები ჩვეულებრივ შეიცავს რამდენიმე ათეულ ქლოროპლასტს ერთ უჯრედში. ქლოროპლასტების მწვანე ფერი დამოკიდებულია მათში ქლოროფილის პიგმენტის შემცველობაზე.
ქლოროპლასტი არის მცენარეთა უჯრედების მთავარი ორგანოიდი, რომელშიც ხდება ფოტოსინთეზი, ანუ ორგანული ნივთიერებების (ნახშირწყლების) წარმოქმნა არაორგანულიდან (CO 2 და H 2 O) მზის ენერგიის გამოყენებით. ქლოროპლასტები სტრუქტურულად მიტოქონდრიის მსგავსია.
ქლოროპლასტს აქვს რთული სტრუქტურა. ისინი ჰიალოპლაზმიდან შემოიფარგლება ორი გარსით - გარე და შიდა. შინაგანი შინაარსი ე.წ სტრომა... შიდა მემბრანა ქმნის მემბრანების რთულ, მკაცრად მოწესრიგებულ სისტემას ბრტყელი ბუშტების სახით, ე.წ. თილაკოიდები.
თილაკოიდები გროვდება გროვაში - მარცვლებიმონეტების სვეტების მსგავსი . გრანა ერთმანეთთან დაკავშირებულია სტრომული თილაკოიდებით, რომლებიც გადიან მათ პლასტიდის გასწვრივ (სურ. 2.20-2.22).ქლოროფილი და ქლოროპლასტები წარმოიქმნება მხოლოდ სინათლის ზემოქმედებით.

ბრინჯი. 2.20. ქლოროპლასტები მსუბუქი მიკროსკოპის ქვეშ

ბრინჯი. 2.21. ქლოროპლასტის სტრუქტურა ელექტრონული მიკროსკოპის ქვეშ

ბრინჯი. 2.22. ქლოროპლასტების სქემატური აგებულება
ფუნქციები
1) ფოტოსინთეზი(არაორგანული ნივთიერებებისგან ორგანული ნივთიერებების წარმოქმნა სინათლის ენერგიის გამო). ქლოროფილი თამაშობს ცენტრალურ როლს ამ პროცესში. ის შთანთქავს სინათლის ენერგიას და მიმართავს მას ფოტოსინთეზური რეაქციების განსახორციელებლად. ქლოროპლასტებში, ისევე როგორც მიტოქონდრიებში, ATP სინთეზირებულია.
2) მონაწილეობა მიიღოს ამინომჟავების და ცხიმოვანი მჟავების სინთეზში,
3) ემსახურება როგორც სახამებლის დროებითი რეზერვების საცავს.
ლეიკოპლასტები- მცირე ზომის უფერო პლასტიდები, რომლებიც გვხვდება მზის შუქისგან დაფარული ორგანოების უჯრედებში (ფესვები, რიზომები, ტუბერები, თესლი). მათი აგებულება ქლოროპლასტების აგებულების მსგავსია. (ნახ. 2.23).
თუმცა, ქლოროპლასტებისაგან განსხვავებით, ლეიკოპლასტებს აქვთ ცუდად განვითარებული შიდა მემბრანული სისტემა, რადგან ისინი მონაწილეობენ სარეზერვო ნუტრიენტების - სახამებლის, ცილების და ლიპიდების სინთეზსა და დაგროვებაში. შუქზე, ლეიკოპლასტები შეიძლება გადაიქცეს ქლოროპლასტებად.

ბრინჯი. 2.23. ლეიკოპლასტის სტრუქტურა
ქრომოპლასტები- ნარინჯისფერი, წითელი და ყვითელი ფერის პლასტიდები, რაც გამოწვეულია კაროტინოიდების ჯგუფის პიგმენტებით. ქრომოპლასტები გვხვდება მრავალი მცენარის ფურცლების უჯრედებში, მწიფე ნაყოფის, იშვიათად ძირეული კულტურების, ასევე შემოდგომის ფოთლებში. ქრომოპლასტების შიდა მემბრანული სისტემა, როგორც წესი, არ არსებობს (სურ. 24).

ბრინჯი. 2.24. ქრომოპლასტის სტრუქტურა
ქრომოპლასტების მნიშვნელობა ჯერ კიდევ ბოლომდე არ არის გასაგები. მათი უმეტესობა დაბერებული პლასტიდებია. ისინი, როგორც წესი, ვითარდება ქლოროპლასტებისგან, ხოლო ქლოროფილი და შიდა მემბრანის სტრუქტურა პლასტიდებში ნადგურდება და კაროტინოიდები გროვდება. ეს ხდება მაშინ, როდესაც ნაყოფი მწიფდება და ფოთლები ყვითლდება შემოდგომაზე. ქრომოპლასტების ბიოლოგიური მნიშვნელობა იმაში მდგომარეობს, რომ ისინი იწვევენ ყვავილებისა და ხილის ნათელ ფერს, რაც იზიდავს მწერებს ჯვარედინი დამტვერვისთვის და სხვა ცხოველებს ხილის გასავრცელებლად. ლეიკოპლასტები ასევე შეიძლება იქცეს ქრომოპლასტებად.
პლასტიდების ფუნქციები
ორგანული ნივთიერებების სინთეზი ქლოროფილში მარტივი არაორგანული ნაერთებისგან: ნახშირორჟანგი და წყალი მზის კვანტების თანდასწრებით - ფოტოსინთეზი, ATP სინთეზი ფოტოსინთეზის მსუბუქ ფაზაში
ცილების სინთეზი რიბოსომებზე (ქლოროპლასტის შიდა გარსებს შორის არის დნმ, რნმ და რიბოსომები, შესაბამისად, ქლოროპლასტებში, ისევე როგორც მიტოქონდრიებში, სინთეზირებულია ცილა, რაც აუცილებელია ამ ორგანელების აქტივობისთვის).
ქრომოპლასტების არსებობა აიხსნება ყვავილების, ხილის, შემოდგომის ფოთლების კოროლას ყვითელი, ნარინჯისფერი და წითელი ფერით.
ლეიკოპლასტები შეიცავს შესანახ ნივთიერებებს (ღეროებში, ფესვებში, ტუბერებში).
ქლოროპლასტები, ქრომოპლასტები და ლეიკოპლასტები უჯრედებს შორის გადასვლის უნარი აქვთ. ასე რომ, როდესაც შემოდგომაზე ხილი მწიფდება ან იცვლის ფოთლის ფერს, ქლოროპლასტები გადაიქცევა ქრომოპლასტებად, ხოლო ლეიკოპლასტები შეიძლება იქცეს ქლოროპლასტებად, მაგალითად, როდესაც კარტოფილის ტუბერები მწვანე ხდება.
ევოლუციური გაგებით, პლასტიდების პირველადი, საწყისი ტიპი არის ქლოროპლასტები, საიდანაც წარმოიშვა დანარჩენი ორი ტიპის პლასტიდები. პლასტიდებს ბევრი მსგავსება აქვთ მიტოქონდრიებთან, რაც განასხვავებს მათ ციტოპლაზმის სხვა კომპონენტებისგან. ეს არის, უპირველეს ყოვლისა, ორი მემბრანის გარსი და შედარებითი გენეტიკური ავტონომია, საკუთარი რიბოზომების და დნმ-ის არსებობის გამო. ორგანელების ამ თავისებურებამ საფუძველი ჩაუყარა აზრს, რომ პლასტიდების და მიტოქონდრიების წინამორბედები იყვნენ ბაქტერიები, რომლებიც ევოლუციის პროცესში აღმოჩნდა ევკარიოტულ უჯრედში აგებული და თანდათან ქლოროპლასტები და მიტოქონდრიები გადაიქცა. (ნახ. 2.25).

ბრინჯი. 2.25. მიტოქონდრიებისა და ქლოროპლასტების წარმოქმნა სიმბიოგენეზის თეორიის მიხედვით
უჯრედი- ცოცხალი სისტემის ელემენტარული ერთეული. ცოცხალი უჯრედის სხვადასხვა სტრუქტურებს, რომლებიც პასუხისმგებელნი არიან კონკრეტული ფუნქციის შესრულებაზე, ეწოდება ორგანელებს, ისევე როგორც მთელი ორგანიზმის ორგანოებს. უჯრედში სპეციფიკური ფუნქციები ნაწილდება ორგანელებს შორის, უჯრედშიდა სტრუქტურებს შორის, რომლებსაც აქვთ გარკვეული ფორმა, როგორიცაა უჯრედის ბირთვი, მიტოქონდრია და ა.შ.
ფიჭური სტრუქტურები:
ციტოპლაზმა... უჯრედის სავალდებულო ნაწილი, რომელიც ჩასმულია პლაზმურ მემბრანასა და ბირთვს შორის. ციტოზოლიარის სხვადასხვა მარილების და ორგანული ნივთიერებების ბლანტი წყალხსნარი, რომელიც გაჟღენთილია ცილის ძაფების სისტემით - ციტოჩონჩხები. უჯრედის ქიმიური და ფიზიოლოგიური პროცესების უმეტესობა ციტოპლაზმაში მიმდინარეობს. სტრუქტურა: ციტოზოლი, ციტოჩონჩხი. ფუნქციები: მოიცავს სხვადასხვა ორგანელებს, უჯრედის შიდა გარემოს
პლაზმური მემბრანა... ცხოველთა, მცენარეთა ყოველი უჯრედი შემოიფარგლება გარემოდან ან სხვა უჯრედებიდან პლაზმური მემბრანით. ამ მემბრანის სისქე იმდენად მცირეა (დაახლოებით 10 ნმ), რომ მისი დანახვა შესაძლებელია მხოლოდ ელექტრონული მიკროსკოპით.

ლიპიდებიისინი ქმნიან მემბრანაში ორმაგ ფენას და პროტეინები გაჟღენთილია მის მთელ სისქეზე, ჩაეფლო ლიპიდურ შრეში სხვადასხვა სიღრმეში ან განლაგებულია მემბრანის გარე და შიდა ზედაპირებზე. ყველა სხვა ორგანელის მემბრანების სტრუქტურა პლაზმური მემბრანის მსგავსია. სტრუქტურა: ლიპიდების, ცილების, ნახშირწყლების ორმაგი ფენა. ფუნქციები: შეზღუდვა, უჯრედის ფორმის შენარჩუნება, დაზიანებისგან დაცვა, ნივთიერებების მიღებისა და მოცილების რეგულირება.
ლიზოსომები... ლიზოსომები მემბრანული ორგანელებია. მათ აქვთ ოვალური ფორმა და დიამეტრი 0,5 მიკრონი. ისინი შეიცავს ფერმენტების ერთობლიობას, რომლებიც ანადგურებენ ორგანულ ნივთიერებებს. ლიზოსომის მემბრანა ძალიან ძლიერია და ხელს უშლის საკუთარი ფერმენტების შეღწევას უჯრედის ციტოპლაზმაში, მაგრამ თუ ლიზოსომა დაზიანებულია რაიმე გარეგანი ზემოქმედებით, მაშინ მთელი უჯრედი ან მისი ნაწილი ნადგურდება.
ლიზოსომები გვხვდება მცენარეების, ცხოველების და სოკოების ყველა უჯრედში.
სხვადასხვა ორგანული ნაწილაკების მონელებისას, ლიზოსომები უზრუნველყოფენ დამატებით „ნედლეულს“ უჯრედში ქიმიური და ენერგეტიკული პროცესებისთვის. შიმშილის დროს ლიზოსომური უჯრედები შლიან ზოგიერთ ორგანელას უჯრედის მოკვლის გარეშე. ეს ნაწილობრივი მონელება უზრუნველყოფს უჯრედს საკვები ნივთიერებების აუცილებელ მინიმუმს გარკვეული დროის განმავლობაში. ზოგჯერ ლიზოსომები შთანთქავს მთელ უჯრედებს და უჯრედების ჯგუფებს, რაც მნიშვნელოვან როლს ასრულებს ცხოველების განვითარებაში. ამის მაგალითია კუდის დაკარგვა, როდესაც თათი ბაყაყად გარდაიქმნება. სტრუქტურა: ოვალური ფორმის ვეზიკულები, გარე მემბრანა, შიგნით ფერმენტები. ფუნქციები: ორგანული ნივთიერებების გაყოფა, მკვდარი ორგანელების განადგურება, დახარჯული უჯრედების განადგურება.
გოლგის კომპლექსი... ბიოსინთეზის პროდუქტები, რომლებიც შედიან ენდოპლაზმური ბადის ღრუებისა და მილაკების სანათურში, კონცენტრირდება და ტრანსპორტირდება გოლჯის აპარატში. ეს ორგანოიდი არის 5-10 მკმ ზომის.

სტრუქტურა: გარსით გარშემორტყმული ღრუები (ვეზიკულები). ფუნქციები: დაგროვება, შეფუთვა, ორგანული ნივთიერებების გამოყოფა, ლიზოსომების წარმოქმნა.
Ენდოპლაზმურ ბადეში... ენდოპლაზმური ბადე არის ორგანული ნივთიერებების სინთეზისა და ტრანსპორტირების სისტემა უჯრედის ციტოპლაზმაში, რომელიც წარმოადგენს დაკავშირებული ღრუების ღია სტრუქტურას.
რიბოზომების დიდი რაოდენობა მიმაგრებულია ენდოპლაზმური ბადის გარსებზე - უჯრედის ყველაზე პატარა ორგანელებზე, რომლებიც 20 ნმ დიამეტრის სფეროს ჰგავს. და შედგება რნმ-ისა და ცილისგან. ცილების სინთეზი ხდება რიბოზომებზე. შემდეგ ახლად სინთეზირებული ცილები ხვდება ღრუების და მილაკების სისტემაში, რომლის გასწვრივ ისინი უჯრედის შიგნით მოძრაობენ. ღრუები, ტუბულები, მილაკები მემბრანებიდან, რიბოსომის მემბრანების ზედაპირზე. ფუნქციები: ორგანული ნივთიერებების სინთეზი რიბოზომების გამოყენებით, ნივთიერებების ტრანსპორტირება.
რიბოზომები... რიბოსომები მიმაგრებულია ენდოპლაზმური ბადის გარსებზე ან თავისუფლად მდებარეობს ციტოპლაზმაში, განლაგებულია ჯგუფებად, მათზე სინთეზირებულია ცილები. ცილის შემადგენლობა, რიბოსომული რნმ ფუნქციები: უზრუნველყოფს ცილის ბიოსინთეზს (ცილის მოლეკულის შეკრება).
მიტოქონდრია... მიტოქონდრია არის ენერგეტიკული ორგანელები. მიტოქონდრიების ფორმა განსხვავებულია, ისინი შეიძლება იყოს დანარჩენი, ღეროების ფორმის, ძაფისებრი, საშუალო დიამეტრით 1 მიკრონი. და სიგრძე 7 მიკრონი. მიტოქონდრიების რაოდენობა დამოკიდებულია უჯრედის ფუნქციურ აქტივობაზე და შეიძლება მიაღწიოს ათეულ ათასობით მწერების მფრინავ კუნთებში. მიტოქონდრია გარედან შემოსაზღვრულია გარე გარსით, ქვემოდან არის შიდა გარსი, რომელიც ქმნის უამრავ გამონაზარდს - cristae.

მიტოქონდრიის შიგნით არის რნმ, დნმ და რიბოსომები. მის მემბრანებში ჩაშენებულია სპეციფიკური ფერმენტები, რომელთა დახმარებით საკვები ნივთიერებების ენერგია გარდაიქმნება ატფ-ის ენერგიად, რომელიც აუცილებელია უჯრედისა და მთლიანად ორგანიზმის სიცოცხლისთვის, მიტოქონდრიაში.
მემბრანა, მატრიცა, გამონაზარდები - cristae. ფუნქციები: ატფ-ის მოლეკულის სინთეზი, საკუთარი ცილების, ნუკლეინის მჟავების, ნახშირწყლების, ლიპიდების სინთეზი, საკუთარი რიბოზომების წარმოქმნა.
პლასტიდები... მხოლოდ მცენარეულ უჯრედში: ლეკოპლასტები, ქლოროპლასტები, ქრომოპლასტები. ფუნქციები: სარეზერვო ორგანული ნივთიერებების დაგროვება, დამბინძურებელი მწერების მოზიდვა, ატფ-ის და ნახშირწყლების სინთეზი. ქლოროპლასტები 4-6 მკმ დიამეტრის მქონე დისკის ან სფეროს ფორმისაა. ორმაგი გარსით - გარე და შიდა. ქლოროპლასტის შიგნით არის რიბოსომის დნმ და სპეციალური მემბრანული სტრუქტურები - მარცვლები, რომლებიც დაკავშირებულია ერთმანეთთან და ქლოროპლასტის შიდა გარსთან. თითოეული ქლოროპლასტი შეიცავს დაახლოებით 50 მარცვლეულს, რომლებიც მოძრაობენ სინათლის უკეთესად დაჭერისთვის. ქლოროფილი განლაგებულია გრან მემბრანებში, მისი წყალობით მზის სინათლის ენერგია გარდაიქმნება ატფ-ის ქიმიურ ენერგიად. ATP ენერგია გამოიყენება ქლოროპლასტებში ორგანული ნაერთების, ძირითადად ნახშირწყლების სინთეზისთვის.
ქრომოპლასტები... ქრომოპლასტებში აღმოჩენილი წითელი და ყვითელი პიგმენტები მცენარის სხვადასხვა ნაწილს წითელ და ყვითელ ფერს აძლევს. სტაფილო, პომიდვრის ხილი.
ლეიკოპლასტები არის სარეზერვო საკვები ნივთიერების - სახამებლის დაგროვების ადგილი. განსაკუთრებით ბევრი ლეიკოპლასტებია კარტოფილის ტუბერების უჯრედებში. შუქზე ლეიკოპლასტები შეიძლება გადაიქცეს ქლოროპლასტებად (რის შედეგადაც კარტოფილის უჯრედები მწვანე ხდება). შემოდგომაზე ქლოროპლასტები გადაიქცევა ქრომოპლასტებად, ხოლო მწვანე ფოთლები და ნაყოფი ყვითელ და წითლად იქცევა.
უჯრედის ცენტრი... შედგება ორი ცილინდრისაგან, ცენტრიოლისაგან, რომლებიც განლაგებულია ერთმანეთის პერპენდიკულარულად. ფუნქციები: ძაფის საყრდენი ღეროების გამყოფისთვის

უჯრედული ჩანართები ჩნდება ციტოპლაზმაში, შემდეგ ქრება უჯრედის სიცოცხლის პროცესში.

გრანულების სახით მკვრივი ჩანართები შეიცავს სარეზერვო საკვებ ნივთიერებებს (სახამებელი, ცილები, შაქარი, ცხიმები) ან უჯრედის ნარჩენ პროდუქტებს, რომელთა ამოღებაც ჯერ შეუძლებელია. მცენარეთა უჯრედების ყველა პლასტიდს აქვს სარეზერვო საკვები ნივთიერებების სინთეზისა და დაგროვების უნარი. მცენარეთა უჯრედებში სარეზერვო საკვები ნივთიერებების დაგროვება ხდება ვაკუოლებში.
მარცვლები, გრანულები, წვეთებიფუნქციები: არამუდმივი წარმონაქმნები, რომლებიც ინახავს ორგანულ ნივთიერებებს და ენერგიას
ბირთვი... ორი მემბრანის ბირთვული მემბრანა, ბირთვული წვენი, ნუკლეოლუსი. ფუნქციები: უჯრედში მემკვიდრეობითი ინფორმაციის შენახვა და მისი გამრავლება, რნმ-ის სინთეზი - საინფორმაციო, სატრანსპორტო, რიბოსომური. ბირთვულ მემბრანაში არის სპორები, რომელთა მეშვეობითაც ხდება ნივთიერებების აქტიური გაცვლა ბირთვსა და ციტოპლაზმას შორის. ბირთვი ინახავს მემკვიდრეობით ინფორმაციას არა მხოლოდ მოცემული უჯრედის ყველა ნიშნისა და თვისების შესახებ, პროცესების შესახებ, რომლებიც უნდა მოხდეს მასზე (მაგალითად, ცილის სინთეზი), არამედ მთლიანად ორგანიზმის მახასიათებლების შესახებ. ინფორმაცია ჩაწერილია დნმ-ის მოლეკულებში, რომლებიც ქრომოსომების ძირითადი ნაწილია. ბირთვში არის ბირთვი. ბირთვი, მასში მემკვიდრეობითი ინფორმაციის შემცველი ქრომოსომების არსებობის გამო, ასრულებს ცენტრის ფუნქციებს, რომელიც აკონტროლებს უჯრედის ყველა სასიცოცხლო აქტივობას და განვითარებას.