Plazma membrananing funktsiyalari. Plazma membranasi Plazmaning tuzilishi va funktsiyasi
Plazma membranasi alohida o'rin tutadi, chunki u hujayrani tashqaridan cheklaydi va hujayradan tashqari muhit bilan bevosita bog'liqdir. U taxminan 10 nm qalinlikda va eng qalin hujayra membranasi hisoblanadi. Asosiy komponentlar oqsillar (60% dan ortiq), lipidlar (taxminan 40%) va uglevodlar (taxminan 1%). Boshqa barcha hujayra membranalari singari, u EPS kanallarida sintezlanadi.
Plazmalemma funktsiyalari.
Transport.
Plazma membranasi yarim o'tkazuvchan, ya'ni. tanlab turli molekulalar turli tezlikda u orqali o'tadi. Membrana orqali moddalarni tashishning ikki yo'li mavjud: passiv va faol transport.
Passiv transport. Passiv transport yoki diffuziya energiya kiritishni talab qilmaydi. Zaryadlanmagan molekulalar kontsentratsiya gradienti bo'ylab tarqaladi, zaryadlangan molekulalarning tashilishi vodorod protonlarining kontsentratsiya gradientiga va elektrokimyoviy proton gradientiga birlashtirilgan transmembran potentsiallar farqiga bog'liq. Qoida tariqasida, membrananing ichki sitoplazmatik yuzasi manfiy zaryadga ega bo'lib, musbat zaryadlangan ionlarning hujayra ichiga kirib borishini osonlashtiradi. Diffuziyaning ikki turi mavjud: oddiy va engil.
Oddiy diffuziya kichik neytral molekulalar (H 2 O, CO 2, O 2), shuningdek, hidrofobik past molekulyar og'irlikdagi organik moddalar uchun xosdir. Ushbu molekulalar membrananing teshiklari yoki kanallari orqali membrana oqsillari bilan hech qanday o'zaro ta'sir qilmasdan, kontsentratsiya gradienti saqlanib qolgan holda o'tishi mumkin.
Yengillashtirilgan diffuziya gidrofil molekulalarga xos bo'lib, ular membrana bo'ylab konsentratsiya gradienti bo'ylab ham tashiladi, lekin printsip bo'yicha maxsus membrana tashuvchisi oqsillari yordamida. formalar.
Osonlashtirilgan diffuziya juda selektivdir, chunki tashuvchi oqsil tashilgan moddani to'ldiruvchi bog'lanish markaziga ega va uzatish oqsildagi konformatsion o'zgarishlar bilan birga keladi. Yengillashtirilgan diffuziyaning mumkin bo'lgan mexanizmlaridan biri quyidagicha: transport oqsili (translokaz) moddani bog'laydi, so'ngra membrananing qarama-qarshi tomoniga yaqinlashadi, bu moddani chiqaradi, dastlabki konformatsiyani oladi va yana transport funktsiyasini bajarishga tayyor bo'ladi. Proteinning o'zi qanday harakatlanishi haqida juda kam narsa ma'lum. Boshqa mumkin bo'lgan uzatish mexanizmi bir nechta tashuvchi oqsillarning ishtirokini o'z ichiga oladi. Bunday holda, dastlab bog'langan birikmaning o'zi bir oqsildan ikkinchisiga o'tadi, membrananing qarama-qarshi tomonida bo'lgunga qadar u yoki bu oqsil bilan ketma-ket bog'lanadi.
Faol transport. Bunday transport uzatish konsentratsiya gradientiga qarshi amalga oshirilganda sodir bo'ladi. Bu hujayra tomonidan energiya sarflashni talab qiladi. Faol transport hujayra ichidagi moddalarning to'planishi uchun xizmat qiladi. Energiya manbai ko'pincha ATP hisoblanadi. Faol tashish uchun energiya manbaiga qo'shimcha ravishda membrana oqsillarining ishtiroki zarur. Hayvon hujayralaridagi faol transport tizimlaridan biri Na va K + ionlarini hujayra membranasi orqali tashish uchun javobgardir. Ushbu tizim deyiladi Na + - K * - nasos. K + ionlarining konsentratsiyasi Na * ionlaridan yuqori bo'lgan hujayra ichidagi muhit tarkibini saqlash uchun javobgardir.
Ikkala ionning konsentratsiya gradienti K + ni hujayraning ichki qismiga, Na + ni esa tashqi tomonga o'tkazish orqali saqlanadi. Ikkala transport ham konsentratsiya gradientiga qarshi sodir bo'ladi. Ionlarning bunday taqsimlanishi hujayralardagi suv miqdorini, nerv hujayralari va mushak hujayralarining qo'zg'aluvchanligini va normal hujayralarning boshqa xususiyatlarini belgilaydi. Na + -K + -nasos oqsildir - ATPazni tashish. Bu fermentning molekulasi oligomer bo'lib, membranadan o'tadi. Nasos ishining to'liq siklida hujayradan hujayralararo moddaga 3 Na + ionlari va teskari yo'nalishda - 2 K + ionlari o'tadi, bunda ATP molekulasining energiyasi ishlatiladi. Kaltsiy ionlarini uzatish uchun transport tizimlari (Ca 2+ -ATPase), proton nasoslari (H + -ATPase) va boshqalar mavjud.
Boshqa moddaning kontsentratsiya gradientining energiyasi hisobiga amalga oshiriladigan moddaning membrana orqali faol o'tkazilishi deyiladi. simptom... Bunday holda, transport ATPazasi ikkala modda uchun bog'lanish joylariga ega. Antiport moddaning konsentratsiya gradientiga qarshi harakatidir. Bunday holda, boshqa modda konsentratsiya gradienti bo'ylab teskari yo'nalishda harakat qiladi. Simptom va antiport (kotransport) aminokislotalarning ichakdan so'rilishi va birlamchi siydikdan glyukozaning reabsorbsiyasi paytida, Na +, K + -ATPase tomonidan yaratilgan Na + ionlarining konsentratsiya gradienti energiyasidan foydalangan holda paydo bo'lishi mumkin.
Tashishning yana 2 turi - endositoz va ekzotsitoz.
Endositoz- katta zarrachalarni hujayra tomonidan ushlash. Endositozning bir necha usullari mavjud: pinotsitoz va fagotsitoz. Odatda ostida pinotsitoz suyuq kolloid zarrachalarning hujayra, ostida tutilishini tushunish fagotsitoz- tanachalarni ushlash (boshqa hujayralargacha zichroq va kattaroq zarralar). Pino va fagotsitozning mexanizmi boshqacha.
Umuman olganda, qattiq zarrachalar yoki suyuqlik tomchilarining hujayra ichiga tashqaridan kirishi geterofagiya deyiladi. Bu jarayon protozoalarda eng keng tarqalgan, ammo u odamlarda (shuningdek, boshqa sutemizuvchilarda) juda muhimdir. Geterofagiya organizmni himoya qilishda (segmentlangan neytrofillar - granulotsitlar; makrofagotsitlar), suyak to'qimasini (osteoklastlar) qayta qurishda, qalqonsimon bez follikulalari tomonidan tiroksin hosil bo'lishida, oqsil va boshqa proksilmal nefronlarda reabsorbtsiyada muhim rol o'ynaydi. va boshqa jarayonlar.
Pinotsitoz.
Tashqi molekulalarning hujayra ichiga kirishi uchun ular birinchi navbatda glikokaliks retseptorlari (membrananing sirt oqsillari bilan bog'langan molekulalar to'plami) bilan bog'lanishi kerak (rasm).
Bunday bog'lanish joyida plazmalemma ostida klatrin oqsil molekulalari topiladi. Plazmalemma tashqi tomondan biriktirilgan molekulalar va sitoplazma ostida joylashgan klatrin bilan birga kirib kela boshlaydi. Protrusion chuqurroq bo'ladi, uning qirralari yaqinlashadi va keyin yopiladi. Natijada ushlangan molekulalarni olib yuruvchi plazmalemmadan pufak ajralib chiqadi. Klatrin uning yuzasida elektron mikrografiyada notekis chegaraga o'xshaydi, shuning uchun bunday pufakchalar chegaralangan deb ataladi.
Klatrin vesikulalarning hujayra ichidagi membranalarga yopishishini oldini oladi. Shunday qilib, chegaralangan pufakchalar hujayra ichida sitoplazmaning tarkibidan foydalanish kerak bo'lgan qismlarga erkin ko'chirilishi mumkin. Bu, xususan, steroid gormonlar yadroga etkazib beriladi. Biroq, odatda, chegaralangan pufakchalar plazmalemmadan ajralganidan keyin tez orada chegaralarini to'kib tashlaydi. Klatrin plazmalemmaga o'tadi va yana endositoz reaktsiyalarida qatnashishi mumkin.
Sitoplazmada hujayra yuzasida ko'proq doimiy pufakchalar - endosomalar mavjud. Chegaralangan pufakchalar klatrinni chiqaradi va endosomalar bilan birlashadi, endosomalarning hajmi va yuzasini oshiradi. Keyin endosomalarning ortiqcha qismi yangi pufakcha shaklida bo'linadi, unda hujayra ichiga kirgan moddalar bo'lmaydi, ular endosomada qoladilar. Yangi vesikula hujayra yuzasiga yo'naltiriladi va membrana bilan birlashadi. Natijada, chegaralangan pufakchaning bo'linishi paytida yuzaga keladigan plazmalemmaning pasayishi tiklanadi, uning retseptorlari ham plazmalemmaga qaytadi.
Endosomalar sitoplazmaga botiriladi va lizosoma membranalari bilan birlashadi. Bunday ikkilamchi lizosoma ichidagi kiruvchi moddalar turli biokimyoviy o'zgarishlarga uchraydi. Jarayon tugagandan so'ng, lizosomaning membranasi bo'laklarga parchalanishi mumkin va parchalanish mahsulotlari va lizosoma tarkibi hujayra ichidagi metabolik reaktsiyalar uchun mavjud bo'ladi. Masalan, aminokislotalar tRNK bilan bog'lanadi va ribosomalarga yetkaziladi, glyukoza esa Golji kompleksiga yoki agranulyar EPS kanalchalariga kirishi mumkin.
Endosomalar klatrin chegarasiga ega bo'lmasa ham, ularning hammasi ham lizosomalar bilan birikmaydi. Ulardan ba'zilari bir hujayra yuzasidan boshqasiga yo'naltiriladi (agar hujayralar epiteliya qatlamini hosil qilsa). U erda endosomaning membranasi plazmolemma bilan birlashadi va tarkibi tashqariga chiqariladi. Natijada moddalar hujayra orqali bir muhitdan ikkinchi muhitga o'zgarishsiz o'tadi. Bu jarayon deyiladi transsitoz... Protein molekulalari, xususan, immunoglobulinlar ham transsitoz orqali tashilishi mumkin.
Fagotsitoz.
Agar katta zarrachaning yuzasida hujayra retseptorlari tomonidan tan olinadigan molekulyar guruhlar mavjud bo'lsa, u bog'lanadi. Hech qachon begona zarralarning o'zlari bunday guruhlarga ega emaslar. Biroq, tanaga kirib, ular doimo qonda va hujayradan tashqari muhitda topilgan immunoglobulinlar (opsoninlar) molekulalari bilan o'ralgan. Immunoglobulinlar har doim fagotsitar hujayralar tomonidan tan olinadi.
Begona zarrachani qoplagan opsoninlar fagotsitning retseptorlari bilan bog'langandan so'ng uning sirt kompleksi faollashadi. Aktin mikrofilamentlari miyozin bilan o'zaro ta'sir qila boshlaydi va hujayra sirtining konfiguratsiyasi o'zgaradi. Fagotsitlar sitoplazmasining o'simtalari zarracha atrofida cho'zilgan. Ular zarrachaning sirtini qoplaydi va uning ustiga birlashadi. O'simtalarning tashqi barglari birlashib, hujayra yuzasini yopadi.
O'simtalarning chuqur barglari so'rilgan zarracha atrofida membranani hosil qiladi - u hosil bo'ladi fagosoma. Fagosoma lizosomalar bilan birlashadi, natijada ularning kompleksi - geterolizosoma (heterosoma, yoki fagolizosoma). Unda zarrachaning ushlangan tarkibiy qismlarining lizisi sodir bo'ladi. Lizis mahsulotlarining ba'zilari geterosomadan chiqariladi va hujayra tomonidan ishlatiladi, ba'zilari esa lizosomal fermentlarning ta'siriga mos kelmasligi mumkin. Bu qoldiqlar qoldiq jismlarni hosil qiladi.
Potentsial barcha hujayralar fagotsitoz qobiliyatiga ega, ammo tanada faqat bir nechtasi bu yo'nalishda ixtisoslashgan. Bular neytrofil leykotsitlar va makrofaglardir.
Ekzotsitoz.
Bu hujayradan moddalarni olib tashlashdir. Birinchidan, yirik molekulyar birikmalar Golji kompleksida transport pufakchalari shaklida ajraladi. Ikkinchisi mikrotubulalar ishtirokida hujayra yuzasiga yo'naltiriladi. Pufakchaning membranasi plazmalemma ichiga singib ketgan va pufakchaning tarkibi hujayradan tashqarida joylashgan (rasm). Bu ekzotsitoz deyiladi konstitutsiyaviy. O'z metabolizmi mahsulotlarining aksariyati hujayradan shunday chiqariladi. Biroq, bir qator hujayralar maxsus birikmalar - tananing boshqa qismlarida ishlatiladigan sirlarni sintez qilish uchun mo'ljallangan. Siri bo'lgan transport pufakchasi plazma membranasi bilan birlashishi uchun tashqi signallar kerak. Shundan keyingina sintez sodir bo'ladi va sir ochiladi. Bu ekzotsitoz deyiladi tartibga solingan... Sekretsiyani bo'shatishga yordam beruvchi signal molekulalari deyiladi liberinlar (ozod qiluvchi omillar), va yo'q qilishning oldini olish - statinlar.
Retseptor funktsiyalari.
Ular asosan plazmalemma yuzasida joylashgan va ularning ligandlari bilan bog'lanishga qodir bo'lgan glikoproteinlar tomonidan ta'minlanadi. Ligand o'z retseptoriga qulfning kaliti kabi mos keladi. Ligandning retseptor bilan bog'lanishi polipeptid konformatsiyasining o'zgarishiga olib keladi. Transmembran oqsilining bunday o'zgarishi bilan hujayradan tashqari va hujayra ichidagi muhit o'rtasida aloqa o'rnatiladi.
Retseptorlarning turlari.
Protein ion kanallari bilan bog'liq retseptorlar. Ular ionlarning o'tishi uchun kanalni vaqtincha ochadigan yoki yopadigan signalizatsiya molekulasi bilan o'zaro ta'sir qiladi. (Masalan, atsetilxolin neyrotransmitterining retseptorlari ion kanalini tashkil etuvchi 5 subbirlikdan tashkil topgan oqsildir. Atsetilxolin bo'lmaganda kanal yopiladi va biriktirilgandan keyin u ochiladi va natriy ionlarining o'tishiga imkon beradi).
Katalitik retseptorlari. Ular hujayradan tashqari qismdan (retseptorning o'zi) va prolin kinaz fermenti (masalan, o'sish gormoni retseptorlari) vazifasini bajaradigan hujayra ichidagi sitoplazmatik qismdan iborat.
G-oqsillari bilan bog'langan retseptorlari. Bular ligand va G-oqsil (guanozin trifosfat bilan bog'langan tartibga soluvchi oqsil) bilan o'zaro ta'sir qiluvchi retseptordan tashkil topgan transmembran oqsillari bo'lib, ular signalni membrana bilan bog'langan fermentga (adenilat siklaza) yoki ion kanaliga uzatadi. Natijada siklik AMP yoki kaltsiy ionlari faollashadi. (Adenilatsiklaza tizimi shunday ishlaydi. Masalan, jigar hujayralarida insulin gormoni uchun retseptor mavjud. Retseptorning yuqori hujayrali qismi insulin bilan bog'lanadi. Bu hujayra ichidagi qism - adenilatsiklaza fermenti faollashishiga olib keladi. U turli xil hujayra ichidagi jarayonlar tezligini tartibga soluvchi, u yoki boshqa metabolik fermentlarning faollashishi yoki inhibisyoniga olib keladigan ATP dan tsiklik AMP sintez qiladi.
Jismoniy omillarni qabul qiluvchi retseptorlar. Masalan, fotoretseptor oqsili rodopsin. Yorug'likni yutganda, u o'zining konformatsiyasini o'zgartiradi va asab impulsini qo'zg'atadi.
U bilipid qatlamdan iborat bo'lib, uning lipidlari qat'iy yo'naltirilgan - lipidlarning hidrofobik qismi (dumi) qatlamning ichki tomoniga, gidrofil qismi (boshi) esa tashqi tomonga qaragan. Plazma membranasini qurishda lipidlardan tashqari uch turdagi membrana oqsillari ishtirok etadi: periferik, integral va yarim integral.
Hozirgi vaqtda membranalarni o'rganish yo'nalishlaridan biri bu membranalarni tashkil etuvchi har xil strukturaviy va tartibga soluvchi lipidlar va individual integral va yarim integral oqsillarning xususiyatlarini batafsil o'rganishdir.
Integral membrana oqsillari
Membrananing o'zini tashkil qilishda asosiy rolni globulyar tuzilishga ega bo'lgan va lipid fazasi bilan gidrofil - hidrofobik o'zaro ta'sirlar bilan bog'langan integral va yarim integral oqsillar o'ynaydi. Integral oqsillarning globulalari membrananing butun qalinligi bo'ylab kirib boradi va ularning hidrofobik qismi globulaning o'rtasida joylashgan bo'lib, lipid fazasining hidrofobik zonasiga botiriladi.
Yarim integral membrana oqsillari
Yarim integral oqsillarda gidrofob aminokislotalar globulaning qutblaridan birida to'plangan va shunga ko'ra, globulalar membranaga faqat yarmiga botib, bir (tashqi yoki ichki) membrana yuzasidan tashqariga chiqib turadi.
Membran oqsillarining vazifalari
Plazma membranasining integral va yarim integral oqsillariga ilgari ikkita funktsiya berilgan: umumiy strukturaviy va o'ziga xos. Shunga ko'ra, ular orasida strukturaviy va funktsional oqsillar ajratilgan. Biroq, membranalarning oqsil fraktsiyalarini ajratish usullarini takomillashtirish va alohida oqsillarni batafsilroq tahlil qilish endi hech qanday o'ziga xos funktsiyalarni bajarmaydigan barcha membranalar uchun universal strukturaviy oqsillarning yo'qligini ko'rsatadi. Aksincha, o'ziga xos funktsiyalarga ega bo'lgan membrana oqsillari juda xilma-xildir. Bular retseptor funktsiyalarini bajaradigan oqsillar, turli birikmalarning faol va passiv tashuvchisi bo'lgan oqsillar va nihoyat, ko'plab ferment tizimlarining bir qismi bo'lgan oqsillardir. Saytdan olingan material
Membran oqsillarining xossalari
Bu barcha integral va yarim integral membrana oqsillarining umumiy xususiyati nafaqat funktsional, balki kimyoviy jihatdan ham farqlanadi, ularning asosiy harakat qilish qobiliyati, suyuq lipid fazasida membrana tekisligida "suzish". Yuqorida ta'kidlanganidek, ba'zi hujayralarning plazma membranalarida bunday harakatlarning mavjudligi eksperimental tarzda isbotlangan. Ammo bu membrana oqsillarida aniqlangan yagona harakat turidan uzoqdir. Yon siljishdan tashqari, individual integral va yarim integral oqsillar membrana tekisligida gorizontal va hatto vertikal yo'nalishda aylanishi mumkin, shuningdek, molekulaning lipid fazaga botirish darajasini o'zgartirishi mumkin.
Opsin. Protein globullarining bu xilma-xil va murakkab harakatlarining barchasi, ayniqsa, fotoretseptor hujayralar membranalari uchun xos bo'lgan opsin oqsili misolida yaxshi ko'rsatilgan (3-rasm). Ma'lumki, opsin zulmatda er-xotin ciss aloqasini o'z ichiga olgan karotenoid retinal bilan bog'liq; retinal va opsin majmuasi rodopsin yoki vizual binafsha rang hosil qiladi. Rodopsin molekulasi membrananing gorizontal tekisligida lateral harakat va aylanishga qodir (3-rasm, A). Yorug'lik ta'sirida ko'zning to'r pardasi fotoizomerizatsiyaga uchraydi va trans shaklga aylanadi. Bunda retinaning konformatsiyasi o'zgaradi va u opsindan ajralib chiqadi, bu esa o'z navbatida aylanish tekisligini gorizontaldan vertikalga o'zgartiradi (3-rasm, B). Bunday o'zgarishlarning natijasi ionlar uchun membrana o'tkazuvchanligining o'zgarishi bo'lib, bu nerv impulsining paydo bo'lishiga olib keladi.
Qizig'i shundaki, yorug'lik energiyasidan kelib chiqqan opsin globulalari konformatsiyasidagi o'zgarishlar nafaqat ko'zning to'r pardasi hujayralarida sodir bo'lgan nerv impulsini yaratishga xizmat qilishi mumkin, balki maxsus binafsha rangli bakteriyalarda joylashgan eng oddiy fotosintetik tizimdir.
Biologik membranalar hujayraning strukturaviy tashkil etilishining asosini tashkil qiladi. Plazma membrana (plazmalemma) tirik hujayraning sitoplazmasini o'rab turgan membranadir. Membranalar lipidlar va oqsillardan tashkil topgan. Lipidlar (asosan fosfolipidlar) ikki qatlam hosil qiladi, bunda molekulalarning hidrofobik "dumlari" membrananing ichki qismiga, hidrofiliklari esa uning sirtiga yo'naltiriladi. Protein molekulalari membrananing tashqi va ichki yuzasida joylashgan bo'lishi mumkin, ular qisman lipid qatlamiga botishi yoki unga va bo'ylab kirib borishi mumkin. Suvga cho'mgan membrana oqsillarining aksariyati fermentlardir. Bu plazma membranasi strukturasining suyuqlik-suyak-mozaik modeli. Protein va lipid molekulalari harakatchan bo'lib, membrananing dinamizmini ta'minlaydi. Membranalar tarkibida glikolipidlar va membrananing tashqi yuzasida joylashgan glikoproteinlar (glikokaliks) holidagi uglevodlar ham mavjud. Har bir hujayraning membrana yuzasida joylashgan oqsillar va uglevodlar to'plami o'ziga xos bo'lib, hujayra turining o'ziga xos ko'rsatkichidir.
Membran funktsiyalari:
- Ajratish. U hujayraning ichki tarkibi va tashqi muhit o'rtasida to'siq hosil qilishdan iborat.
- Sitoplazma va tashqi muhit o'rtasida moddalar almashinuvini ta'minlash. Hujayra ichiga suv, ionlar, noorganik va organik molekulalar kiradi (transport funktsiyasi). Hujayrada hosil bo'lgan mahsulotlar (sekret funktsiyasi) tashqi muhitga chiqariladi.
- Transport. Membrana bo'ylab tashish turli yo'llar bilan davom etishi mumkin. Passiv tashish energiya sarfisiz, oddiy diffuziya, osmoz yoki tashuvchi oqsillar yordamida osonlashtirilgan diffuziya orqali amalga oshiriladi. Faol transport tashuvchi oqsillar yordamida amalga oshiriladi va u energiya talab qiladi (masalan, natriy-kaliy nasosi). Saytdan olingan material
Endositoz natijasida hujayra ichiga biopolimerlarning yirik molekulalari kiradi. U fagotsitoz va pinotsitozga bo'linadi. Fagotsitoz - katta zarrachalarning hujayra tomonidan tutilishi va so'rilishi. Bu hodisani birinchi marta I.I. Mechnikov. Birinchidan, moddalar plazma membranasiga, o'ziga xos retseptor oqsillariga yopishadi, so'ngra membrana cho'kib, depressiya hosil qiladi.
Ovqat hazm qilish vakuolasi hosil bo'ladi. U hujayra ichiga kirgan moddalarni hazm qiladi. Odamlar va hayvonlarda leykotsitlar fagotsitozga qodir. Oq qon hujayralari bakteriyalar va boshqa zarrachalarni o'zlashtiradi.
Pinotsitoz - suyuqlik tomchilarini unda erigan moddalar bilan ushlash va singdirish jarayoni. Moddalar membrana oqsillariga (retseptorlariga) yopishadi va eritma tomchisi membrana bilan o'ralgan bo'lib, vakuola hosil qiladi. ATP energiyasining sarflanishi bilan pinotsitoz va fagotsitoz sodir bo'ladi.
- Sekretsiya. Sekretsiya - hujayrada sintez qilingan moddalarning hujayra tomonidan tashqi muhitga chiqishi. Gormonlar, polisaxaridlar, oqsillar, yog 'tomchilari membrana bilan chegaralangan pufakchalarga o'raladi va plazmalemmaga boradi. Membranalar birlashadi va vesikulaning tarkibi hujayrani o'rab turgan muhitga chiqariladi.
- To'qimalarda hujayralarning ulanishi (katlanmış o'sishlar tufayli).
- Retseptor. Membranalar ko'p sonli retseptorlarni o'z ichiga oladi - maxsus oqsillar, ularning roli signallarni tashqi tomondan hujayraning ichki qismiga o'tkazishdir.
1. To‘siq- atrof-muhit bilan tartibga solinadigan, selektiv, passiv va faol metabolizmni ta'minlaydi.
Hujayra membranalari mavjud selektiv o'tkazuvchanlik: glyukoza, aminokislotalar, yog 'kislotalari, glitserin va ionlar ular orqali asta-sekin tarqaladi, membranalarning o'zi bu jarayonni faol ravishda tartibga soladi - ba'zi moddalarga ruxsat beriladi, boshqalari esa yo'q.
2. Transport- membrana orqali moddalar hujayra ichiga va tashqarisiga o'tkaziladi. Membranalar orqali tashish quyidagilarni ta'minlaydi: ozuqa moddalarini etkazib berish, so'nggi metabolik mahsulotlarni olib tashlash, turli moddalar sekretsiyasi, ion gradientlarini yaratish, hujayra fermentlarining ishlashi uchun zarur bo'lgan hujayradagi tegishli pH va ion konsentratsiyasini saqlash.
Moddalarning hujayra ichiga kirishi yoki ularni hujayradan tashqariga chiqarishning to'rtta asosiy mexanizmi mavjud:
a) passiv (diffuziya, osmoz) (energiya sarfini talab qilmaydi)
Diffuziya
Bir moddaning molekulalari yoki atomlarining molekulalar yoki boshqasining atomlari o'rtasida tarqalishi, bu ularning kontsentratsiyasining egallangan hajm bo'ylab o'z-o'zidan tenglashishiga olib keladi. Ba'zi hollarda moddalardan biri allaqachon tenglashtirilgan konsentratsiyaga ega va ular bir moddaning boshqasiga tarqalishi haqida gapiradi. Bunday holda, moddaning yuqori konsentratsiyali hududdan past konsentratsiyali hududga (kontsentratsiya gradienti vektori bo'ylab) o'tishi sodir bo'ladi. (2.4-rasm).
Guruch. 2.4. Diffuziya jarayoni diagrammasi
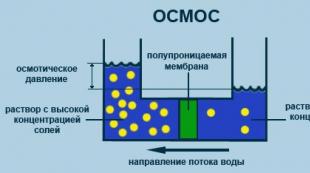
Osmos
Erituvchi molekulalarning yarim o'tkazuvchan membranasi orqali erigan moddaning konsentratsiyasi past bo'lgan hajmdan erigan moddaning yuqori konsentratsiyasiga qarab bir tomonlama diffuziya jarayoni. (2.5-rasm).

Guruch. 2.5. Osmos jarayoni diagrammasi
b) Faol transport (energiya sarfini talab qiladi)
Natriy-kaliyli nasos- natriy ionlarini (hujayradan) va kaliy ionlarini (hujayraga) faol konjugatsiyalangan transmembran tashish mexanizmi, bu konsentratsiya gradienti va transmembran potentsial farqini ta'minlaydi. Ikkinchisi hujayralar va organlarning ko'plab funktsiyalari uchun asos bo'lib xizmat qiladi: bez hujayralarining sekretsiyasi, mushaklarning qisqarishi, nerv impulslarini o'tkazish va boshqalar. (2.6-rasm).

Guruch. 2.6. Kaliy-natriy nasosining sxemasi
Birinchi bosqichda Na + / K + -ATPase fermenti membrananing ichki tomonidan uchta Na + ionini biriktiradi. Ushbu ionlar ATPazning faol markazining konformatsiyasini o'zgartiradi. Shundan so'ng, ferment bitta ATP molekulasini gidrolizlashga qodir. Gidrolizdan keyin chiqarilgan energiya tashuvchining konformatsiyasini o'zgartirishga sarflanadi, buning natijasida membrananing tashqi tomonida uchta ion Na + va PO 4 3- ioni (fosfat) paydo bo'ladi. Bu erda Na + ionlari bo'linadi va PO 4 3 - ikkita K + ionlari bilan almashtiriladi. Shundan so'ng ferment o'zining dastlabki konformatsiyasiga qaytadi va membrananing ichki tomonida K + ionlari paydo bo'ladi. Bu erda K + ionlari bo'linadi va tashuvchi yana ishlashga tayyor.
Natijada hujayradan tashqari muhitda Na + ionlarining yuqori konsentratsiyasi, hujayra ichida esa K + ning yuqori konsentratsiyasi hosil bo'ladi. Bu kontsentratsiya farqi nerv impulsini o'tkazish paytida hujayralarda qo'llaniladi.
c) endotsitoz (fagotsitoz, pinotsitoz)
Fagotsitoz(hujayra tomonidan ovqatlanish) - hujayra tomonidan qattiq jismlarni, masalan, eukaryotik hujayralar, bakteriyalar, viruslar, o'lik hujayralar qoldiqlari va boshqalarni singdirish jarayoni. So'rilgan ob'ekt atrofida katta hujayra ichidagi vakuola (fagosoma) hosil bo'ladi. Fagosomalarning o'lchami 250 nm va undan yuqori. Fagosomani birlamchi lizosoma bilan birlashtirib, ikkilamchi lizosoma hosil bo'ladi. Kislotali muhitda gidrolitik fermentlar ikkilamchi lizosomadagi makromolekulalarni parchalaydi. Keyinchalik parchalanish mahsulotlari (aminokislotalar, monosaxaridlar va boshqa foydali moddalar) lizosomal membrana orqali hujayraning sitoplazmasiga o'tkaziladi. Fagotsitoz juda keng tarqalgan. Yuqori darajada tashkil etilgan hayvonlar va odamlarda fagotsitoz jarayoni himoya rolini o'ynaydi. Leykotsitlar va makrofaglarning fagotsitar faolligi organizmni patogen mikroblar va unga kiruvchi boshqa kiruvchi zarralardan himoya qilishda katta ahamiyatga ega. Fagotsitozni birinchi marta rus olimi I.I.Mechnikov ta'riflagan (2.7-rasm)
Pinotsitoz(hujayra tomonidan ichish) - eruvchan moddalar, shu jumladan yirik molekulalar (oqsillar, polisaxaridlar va boshqalar) bo'lgan muhitdan suyuqlik fazasining hujayra tomonidan so'rilishi jarayoni. Pinotsitoz bilan hujayra ichidagi membranadan kichik pufakchalar - endosomalar ajralib chiqadi. Ular fagosomalardan kichikroq (ularning o'lchami 150 nm gacha) va odatda katta zarrachalarni o'z ichiga olmaydi. Endosoma hosil bo'lgandan keyin birlamchi lizosoma unga yaqinlashadi va bu ikki membrana pufakchalari birlashadi. Olingan organella ikkilamchi lizosoma deb ataladi. Pinotsitoz jarayoni doimo barcha eukaryotik hujayralar tomonidan amalga oshiriladi. (7-rasm)
Retseptor vositachiligidagi endositoz - faol o'ziga xos jarayon, bunda hujayra membranasi hujayra ichiga bo'rtib, qirrali chuqurlarni hosil qiladi. Chiziqli chuqurchaning hujayra ichidagi tomonida moslashuvchan oqsillar to'plami mavjud. Hujayra yuzasida o'ziga xos retseptorlar bilan bog'langan makromolekulalar hujayra ichiga pinotsitoz orqali kiradigan moddalarga qaraganda ancha yuqori tezlikda o'tadi.

Guruch. 2.7. Endositoz
d) ekzotsitoz (salbiy fagotsitoz va pinotsitoz)
Hujayra ichidagi pufakchalar (membrana pufakchalari) tashqi hujayra membranasi bilan birikadigan hujayrali jarayon. Ekzotsitoz jarayonida sekretor pufakchalarning tarkibi (ekzositar pufakchalar) tashqariga chiqariladi va ularning membranasi hujayra membranasi bilan birlashadi. Bu usul bilan hujayradan deyarli barcha makromolekulyar birikmalar (oqsillar, peptid gormonlar va boshqalar) ajralib chiqadi. (2.8-rasm)

Guruch. 2.8. Ekzotsitoz sxemasi
3. Biopotensiallarni hosil qilish va o‘tkazish- membrana yordamida hujayrada ionlarning doimiy kontsentratsiyasi saqlanadi: hujayra ichidagi K + ionining konsentratsiyasi tashqariga qaraganda ancha yuqori va Na + kontsentratsiyasi ancha past, bu juda muhim, chunki bu membranadagi potentsial farqning saqlanishini va nerv impulsining paydo bo'lishini ta'minlaydi.
4. Mexanik- hujayraning avtonomiyasini, uning hujayra ichidagi tuzilmalarini, shuningdek, boshqa hujayralar (to'qimalarda) bilan bog'lanishini ta'minlaydi.
5. Energiya- xloroplastlarda fotosintez va mitoxondriyalarda hujayrali nafas olish jarayonida ularning membranalarida oqsillar ham ishtirok etadigan energiya uzatish tizimlari ishlaydi;
6. Retseptor- membranadagi ba'zi oqsillar retseptorlar (hujayra ma'lum signallarni qabul qiladigan molekulalar).
7. Enzimatik- membrana oqsillari ko'pincha fermentlardir. Masalan, ichak epiteliy hujayralarining plazma membranalarida ovqat hazm qilish fermentlari mavjud.
8. Matritsa- membrana oqsillarining ma'lum bir o'zaro joylashishi va yo'nalishini, ularning optimal o'zaro ta'sirini ta'minlaydi;
9. Qafasni belgilash- membranada marker sifatida ishlaydigan antijenler mavjud - hujayrani aniqlash imkonini beruvchi "yorliqlar". Bular "antennalar" rolini o'ynaydigan glikoproteinlar (ya'ni, ularga tarvaqaylab ketgan oligosakkarid yon zanjirlari bo'lgan oqsillar). Markerlar yordamida hujayralar boshqa hujayralarni taniydi va ular bilan birgalikda harakat qiladi, masalan, organlar va to'qimalarning shakllanishi paytida. Shuningdek, u immunitet tizimiga begona antijenlarni tanib olish imkonini beradi.
Uyali birikmalar
Hujayra tarkibiga uglevodlar, yog'lar va oqsillar kiradi. Bu moddalarning barchasi hujayra sitoplazmasida turli o'lcham va shakldagi tomchilar va donalar shaklida to'planadi. Ular vaqti-vaqti bilan hujayrada sintezlanadi va metabolik jarayonda qo'llaniladi.
Sitoplazma
U tirik hujayraning (protoplast) plazma membranasi va yadrosiz qismidir. Sitoplazma tarkibiga quyidagilar kiradi: sitoplazmatik matritsa, sitoskeleton, organellalar va inklyuziyalar (ba'zan qo'shimchalar va vakuolalarning tarkibi sitoplazmaning tirik moddasi sifatida tasniflanmaydi). Plazma membranasi bilan tashqi muhitdan ajratilgan sitoplazma hujayralarning ichki yarim suyuq muhiti hisoblanadi. Yadro va turli organellalar eukaryotik hujayralar sitoplazmasida joylashgan. Shuningdek, u turli xil qo'shimchalarni o'z ichiga oladi - hujayra faolligi mahsulotlari, vakuolalar, shuningdek, hujayra skeletini tashkil etuvchi eng kichik tubulalar va filamentlar. Sitoplazmaning asosiy moddasi tarkibida oqsillar ustunlik qiladi.
Sitoplazmatik funktsiyalar
1) unda asosiy metabolik jarayonlar sodir bo'ladi.
2) yadro va barcha organoidlarni bir butunga birlashtiradi, ularning o'zaro ta'sirini ta'minlaydi.
3) harakatchanlik, asabiylashish, metabolizm va ko'payish.
Mobillik turli shakllarda bo'ladi:
Hujayra sitoplazmasining hujayra ichidagi harakati.
Amyoba shaklidagi harakat. Harakatning bu shakli sitoplazma tomonidan u yoki bu stimulga qarab yoki undan uzoqda psevdopodiya hosil bo'lishida ifodalanadi. Ushbu harakat shakli amyoba, qon leykotsitlari, shuningdek, ba'zi to'qimalar hujayralariga xosdir.
Miltillovchi harakat. U o'zini mayda protoplazmatik o'simtalar - siliya va flagella (kipriklar, ko'p hujayrali hayvonlarning epitelial hujayralari, sperma va boshqalar) urishi shaklida namoyon bo'ladi.
Shartnoma harakati. Bu sitoplazmada qisqarishi yoki cho'zilishi hujayraning qisqarishi va bo'shashishiga yordam beradigan maxsus organoid miyofibrillarning mavjudligi tufayli ta'minlanadi. Qisqartirish qobiliyati mushak hujayralarida eng rivojlangan.
Achchiqlanish hujayralarning metabolizm va energiyani o'zgartirish orqali tirnash xususiyati bilan javob berish qobiliyatida ifodalanadi.
Sitoskelet
Eukaryotik hujayraning o'ziga xos xususiyatlaridan biri uning sitoplazmasida mikronaychalar va oqsil tolalari to'plamlari ko'rinishidagi skelet shakllanishlarining mavjudligidir. Tashqi sitoplazmatik membrana va yadro qobig'i bilan chambarchas bog'langan sitoskeletning elementlari sitoplazmada murakkab to'qimalarni hosil qiladi.
Sitoskeleton mikronaychalar, mikrofilamentlar va mikrotrabekulyar tizimdan hosil bo'ladi. Sitoskelet hujayraning shaklini belgilaydi, hujayra harakatlarida, hujayraning bo'linishi va harakatlanishida, organellalarning hujayra ichidagi transportida ishtirok etadi.
Mikronaychalar barcha eukaryotik hujayralarda mavjud bo'lib, diametri 30 nm dan oshmaydigan va devor qalinligi 5 nm bo'lgan ichi bo'sh, tarmoqlanmagan silindrlardir. Ularning uzunligi bir necha mikrometrga etishi mumkin. Osonlik bilan parchalanadi va qayta yig'iladi. Mikronaychalar devori asosan spiral buklangan tubulin oqsili bo'linmalaridan iborat. (2.09-rasm)
Mikrotubulalar funktsiyalari:
1) yordamchi funktsiyani bajarish;
2) parchalanish shpindelini hosil qiladi; xromosomalarning hujayra qutblariga ajralishini ta'minlash; hujayra organellalarining harakati uchun javobgardir;
3) hujayra ichidagi tashish, sekretsiya, hujayra devorining shakllanishida ishtirok etish;
4) kiprikchalar, flagellalar, bazal tanachalar va sentriolalarning tuzilish qismidir.
Mikrofilamentlar mushak aktiniga yaqin bo'lgan aktin oqsilidan tashkil topgan diametri 6 nm bo'lgan filamentlar bilan ifodalanadi. Aktin hujayradagi umumiy oqsilning 10-15% ni tashkil qiladi. Ko'pgina hayvonlar hujayralarida plazma membranasi ostida aktin filamentlari va ular bilan bog'liq bo'lgan oqsillarning zich tarmog'i hosil bo'ladi.
Hujayrada aktindan tashqari miyozin filamentlari ham uchraydi. Biroq, ularning soni ancha kam. Aktin va miyozinning o'zaro ta'siri tufayli mushaklarning qisqarishi sodir bo'ladi. Mikrofilamentlar butun hujayra yoki uning ichidagi alohida tuzilmalarning harakati bilan bog'liq. Ba'zi hollarda harakat faqat aktin filamentlari bilan ta'minlanadi, boshqalarida - miyozin bilan birga aktin.
Mikrofilamentlarning vazifalari
1) mexanik kuch
2) hujayra shaklini o'zgartirish va harakat qilish imkonini beradi.

Guruch. 2.09. Sitoskelet
Organellalar (yoki organellalar)
ga bo'linadi membranasiz, bir membranali va ikki membranali.
TO membrana bo'lmagan organellalar Eukaryotik hujayralar o'zlarining yopiq membranasiga ega bo'lmagan organellalarni o'z ichiga oladi, xususan: ribosomalar va tubulin mikronaychalari asosida qurilgan organellalar - hujayra markazi (tsentriolalar) va harakat organellalari (flagella va siliya). Ko'pgina bir hujayrali organizmlarning hujayralarida va yuqori (erlik) o'simliklarning aksariyatida sentriolalar mavjud emas.
TO bir membranali organellalar bog'lash: endoplazmatik retikulum, Golji apparati, lizosomalar, peroksizomalar, sferosomalar, vakuolalar va boshqalar. Barcha bir membranali organellalar bitta hujayra tizimiga o'zaro bog'langan. O'simlik hujayralarida maxsus lizosomalar, hayvonlar hujayralarida maxsus vakuolalar mavjud: ovqat hazm qilish, chiqarish, qisqarish, fagotsitar, avtofagotsitar va boshqalar.
TO ikki membranali organellalar bog'lash mitoxondriyalar va plastidalar.
Membrana bo'lmagan organellalar
A) Ribosomalar- barcha organizmlarning hujayralarida joylashgan organellalar. Bu diametri taxminan 20 nm bo'lgan globulyar zarrachalar bilan ifodalangan kichik organellalardir. Ribosomalar teng bo'lmagan kattalikdagi ikkita kichik birlikdan iborat - katta va kichik. Ribosomalarga oqsillar va ribosoma RNK (rRNK) kiradi. Ribosomalarning ikkita asosiy turi mavjud: eukaryotik (80S) va prokaryotik (70S).
Hujayradagi lokalizatsiyaga qarab, sitoplazmada erkin ribosomalar joylashgan bo'lib, ular oqsillarni va biriktirilgan ribosomalarni sintez qiladi - EPR membranalarining tashqi yuzasi bilan katta bo'linmalar bilan bog'langan ribosomalar, Golji kompleksiga kiradigan oqsillarni sintez qiladi va keyin ajralib chiqadi. hujayra. Protein biosintezi jarayonida ribosomalar komplekslarga - poliribosomalarga (polisomalarga) birlashishi mumkin.
Eukaryotik ribosomalar yadroda hosil bo'ladi. Birinchidan, rRNK nukleolyar DNKda sintezlanadi, so'ngra sitoplazmadan keladigan ribosoma oqsillari bilan qoplanadi, kerakli o'lchamga ajraladi va ribosoma subbirliklarini hosil qiladi. Yadroda to'liq shakllangan ribosomalar mavjud emas. Subbirliklarning butun ribosomaga birlashishi sitoplazmada, odatda oqsil biosintezi jarayonida sodir bo'ladi.
Ribosomalar barcha organizmlarning hujayralarida mavjud. Har biri kichik va katta ikkita zarrachadan iborat. Ribosomalarga oqsillar va RNK kiradi.
Funksiyalar
oqsil sintezi.
Sintezlangan oqsillar avval endoplazmatik retikulumning kanallari va bo'shliqlarida to'planadi, so'ngra hujayraning organellalari va qismlariga o'tkaziladi. Uning membranalarida joylashgan EPS va ribosomalar oqsillarni biosintez qilish va tashish uchun yagona apparatdir. (2.10-2.11-rasm).

Guruch. 2.10. Ribosoma tuzilishi

Guruch. 2.11. Ribosoma tuzilishi
B) Hujayra markazi (tsentriolalar)
Tsentriol silindr (uzunligi 0,3 mkm va diametri 0,1 mkm) bo'lib, uning devori o'zaro bog'lanish orqali ma'lum vaqt oralig'ida o'zaro bog'langan uchta birlashtirilgan mikronaychalarning to'qqiz guruhidan (9 ta uchlik) hosil bo'ladi. Sentriolalar ko'pincha juft bo'lib, ular bir-biriga to'g'ri burchak ostida joylashgan. Agar tsentriola kiprikcha yoki flagellumning tagida yotsa, u bazal tana deyiladi.
Deyarli barcha hayvonlar hujayralarida hujayra markazining o'rta elementi bo'lgan bir juft sentriola mavjud.
Bo'linishdan oldin sentriolalar qarama-qarshi qutblarga ajralib chiqadi va ularning har birining yonida qiz tsentriola paydo bo'ladi. Hujayraning turli qutblarida joylashgan sentriolalardan bir-biriga qarab o'sib boruvchi mikronaychalar hosil bo'ladi.
Funksiyalar
1) genetik materialning qiz hujayralari o'rtasida bir xil taqsimlanishiga yordam beradigan mitotik shpindel hosil qiladi;
2) sitoskeletonni tashkil qilish markazidir. Shpindel filamentlarining bir qismi xromosomalarga biriktirilgan.
Sentriolalar sitoplazmaning o'z-o'zidan ko'payadigan organellalaridir. Ular mavjud bo'lganlarning takrorlanishi natijasida paydo bo'ladi. Bu sentriolalar ajralib chiqqanda sodir bo'ladi. Yetilmagan sentriolda 9 ta bitta mikronaycha bor; aftidan, har bir mikrotubula etuk sentriolaga xos bo'lgan uchliklarni yig'ish uchun matritsadir. (2.12-rasm).
Tsetriollar quyi o'simliklar (yosunlar) hujayralarida uchraydi.

Guruch. 2.12. Hujayra markazi sentriolalari
Yagona membranali organellalar
D) endoplazmatik retikulum (EPS)
Sitoplazmaning butun ichki zonasi ko'p sonli kichik kanallar va bo'shliqlar bilan to'ldirilgan bo'lib, ularning devorlari tuzilishi bo'yicha plazma membranasiga o'xshash membranalardir. Bu kanallar shoxlanadi, bir-biriga bog'lanadi va endoplazmatik to'r deb ataladigan tarmoq hosil qiladi. Endoplazmatik retikulum tuzilishi jihatidan heterojendir. Uning ikki turi ma'lum - donador va silliq.
Granüler tarmoqning kanallari va bo'shliqlari membranalarida ko'plab mayda yumaloq jismlar mavjud - ribosoma bu membranalarga qo'pol ko'rinish beradi. Silliq endoplazmatik retikulumning membranalari yuzasida ribosomalarni olib yurmaydi. EPS juda ko'p turli funktsiyalarni bajaradi.
Funksiyalar
Donador endoplazmatik retikulumning asosiy vazifasi ribosomalarda amalga oshiriladigan oqsil sintezida ishtirok etishdir. Silliq endoplazmatik retikulumning membranalarida lipidlar va uglevodlar sintezlanadi. Ushbu sintez mahsulotlarining barchasi kanallar va bo'shliqlarda to'planadi, so'ngra hujayraning turli organellalariga ko'chiriladi, u erda ular iste'mol qilinadi yoki hujayra qo'shimchalari sifatida sitoplazmada to'planadi. EPS hujayraning asosiy organellalarini bog'laydi (2.13-rasm).

Guruch. 2.13. Endoplazmatik retikulum (EPS) yoki retikulumning tuzilishi
E) Golji apparati
Ushbu organoidning tuzilishi shaklining xilma-xilligiga qaramay, o'simlik va hayvon organizmlari hujayralarida o'xshashdir. U juda ko'p muhim funktsiyalarga ega.
Yagona membrana organoidi. Bu cho'zilgan qirralari bo'lgan tekislangan "sisternalar" to'plami bo'lib, ular bilan kichik bir membranali pufakchalar tizimi (Golji pufakchalari) bog'langan. Golji pufakchalari, asosan, EPSga tutashgan tomonda va steklarning chetida to'plangan. Ular oqsillar va lipidlarni Golji apparatiga o'tkazadi, deb ishoniladi, ularning molekulalari sisternadan sisternaga o'tib, kimyoviy modifikatsiyadan o'tadi.
Bu moddalarning barchasi avval to'planib, kimyoviy jihatdan murakkablashadi, so'ngra katta va kichik pufakchalar shaklida sitoplazmaga kiradi va hayot davomida hujayraning o'zida ishlatiladi yoki undan chiqariladi va organizmda ishlatiladi. (2.14-2.15-rasm).

Guruch. 2.14. Golji apparatining tuzilishi
Funksiyalar:
Oqsillar, lipidlar, uglevodlarni o'zgartirish va to'plash;
Kiruvchi organik moddalarning membrana pufakchalariga (pufakchalariga) qadoqlash;
Lizosomalar hosil bo'lish joyi;
Sekretsiya funktsiyasi, shuning uchun Golji apparati sekretor hujayralarda yaxshi rivojlangan.

Guruch. 2.15. Golji kompleksi
E) Lizosomalar
Ular kichik dumaloq jismlardir. Lizosoma ichida oqsillarni, yog'larni, uglevodlarni, nuklein kislotalarni parchalaydigan fermentlar mavjud. Lizosomalar sitoplazmaga kirgan oziq-ovqat zarrachasiga yaqinlashadi, u bilan birlashadi va bitta hazm qilish vakuolasi hosil bo'ladi, uning ichida lizosomal fermentlar bilan o'ralgan oziq-ovqat zarrasi mavjud.
Lizosomal fermentlar qo'pol EPSda sintezlanadi, Golji apparatiga o'tkaziladi, u erda o'zgartiriladi va lizosomalarning membrana pufakchalariga o'raladi. Lizosoma 20 dan 60 tagacha turli xil gidrolitik fermentlarni o'z ichiga olishi mumkin. Fermentlar yordamida moddalarning parchalanishi deyiladi lizis.
Birlamchi va ikkilamchi lizosomalarni farqlang. Birlamchi lizosomalar Golji apparatidan kurtaklangan deb ataladi.
Birlamchi lizosomalarning endotsitik vakuolalar bilan birlashishi natijasida hosil bo'lgan ikkilamchi lizosomalar deyiladi. Bunday holda, ular fagotsitoz yoki pinotsitoz yo'li bilan hujayra ichiga kirgan moddalarni hazm qiladilar, shuning uchun ularni ovqat hazm qilish vakuolalari deb atash mumkin.
Lizosomalarning funktsiyalari:
1) endotsitoz paytida hujayra tomonidan ushlangan moddalar yoki zarrachalarni (bakteriyalar, boshqa hujayralar) hazm qilish;
2) autofagiya - hujayra uchun keraksiz tuzilmalarni yo'q qilish, masalan, eski organellalarni yangilari bilan almashtirish yoki hujayraning o'zida hosil bo'lgan oqsillar va boshqa moddalarni hazm qilish;
3) avtoliz - hujayraning o'limiga olib keladigan o'z-o'zini hazm qilish (ba'zida bu jarayon patologik emas, balki tananing rivojlanishi yoki ba'zi maxsus hujayralarning farqlanishi bilan birga keladi) (2.16-2.17-rasm).
Misol: Tadpoya qurbaqaga aylanganda, dum hujayralaridagi lizosomalar uni hazm qiladi: dum yo'qoladi va bu jarayonda hosil bo'lgan moddalar tananing boshqa hujayralari tomonidan so'riladi va ishlatiladi.

Guruch. 2.16. Lizosoma shakllanishi

Guruch. 2.17. Lizosomalarning ishlashi
G) Peroksizomalar
Organoidlar, tuzilishi jihatidan lizosomalarga o'xshash, 50 ga yaqin fermentlarni o'z ichiga olgan bir hil matritsaga ega bo'lgan diametri 1,5 mikrongacha bo'lgan pufakchalar.
Katalaz vodorod peroksid 2H 2 O 2 → 2H 2 O + O 2 parchalanishiga olib keladi va lipid peroksidlanishini oldini oladi.
Peroksizomalar ilgari mavjud bo'lganlardan tomurcuklanma orqali hosil bo'ladi, ya'ni. tarkibida DNK bo'lmasa ham, o'z-o'zidan ko'payadigan organellalarga tegishli. Ular fermentlarning kirib borishi hisobiga o'sadi, dag'al EPS va gialoplazmada peroksizoma fermentlari hosil bo'ladi. (2.18-rasm).

Guruch. 2.18. Peroksisoma (kristalli nukleoidning markazida)
H) Vakuolalar
Yagona membranali organellalar. Vakuolalar - bu organik va noorganik moddalarning suvli eritmalari bilan to'ldirilgan "idishlar". Vakuolalarni hosil qilishda EPS va Golji apparati ishtirok etadi.
Yosh o'simlik hujayralarida ko'plab mayda vakuolalar mavjud bo'lib, ular hujayralar o'sishi va farqlanishi bilan bir-biri bilan qo'shilib, bitta yirik markaziy vakuolani hosil qiladi.
Markaziy vakuola etuk hujayra hajmining 95% gacha egallashi mumkin, yadro va organellalar esa hujayra membranasiga qaytariladi. O'simlik vakuolasini cheklovchi membrana deyiladi tonoplast.
O'simlik vakuolasini to'ldiruvchi suyuqlik hujayra shirasi deb ataladi. Hujayra shirasining tarkibiga suvda eriydigan organik va noorganik tuzlar, monosaxaridlar, disaxaridlar, aminokislotalar, oxirgi yoki zaharli metabolik mahsulotlar (glikozidlar, alkaloidlar), ayrim pigmentlar (antosianinlar) kiradi.
Organik moddalardan shakar va oqsillar ko'proq saqlanadi. Shakarlar - ko'pincha eritmalar shaklida, oqsillar EPR pufakchalari va Golji apparati shaklida bo'ladi, shundan so'ng vakuolalar suvsizlanib, aleyron donalariga aylanadi.
Hayvonlar hujayralarida ikkilamchi lizosomalar guruhiga kiruvchi va gidrolitik fermentlarni o'z ichiga olgan kichik hazm qilish va avtofagik vakuolalar mavjud. Bir hujayrali hayvonlarda osmoregulyatsiya va chiqarish funktsiyasini bajaradigan qisqaruvchi vakuolalar ham mavjud.
Funksiyalar
O'simliklarda
1) suyuqlikning to'planishi va turgorni ushlab turish;
2) zahiradagi ozuqa moddalari va mineral tuzlarning to'planishi;
3) gullar va mevalarni bo'yash va shu bilan meva va urug'larni changlatuvchilar va tarqatuvchilarni jalb qilish.
Hayvonlarda:
4) ovqat hazm qilish vakuolalari - organik makromolekulalarni yo'q qiladi;
5) kontraktil vakuolalar hujayraning osmotik bosimini tartibga soladi va hujayradan keraksiz moddalarni olib tashlaydi.
6) fagotsitar vakuolalar antigenlarning immun hujayralari tomonidan fagotsitoz paytida hosil bo'ladi.
7) o'z to'qimalarining immun hujayralari tomonidan fagotsitoz paytida avtofagotsitar vakuolalar hosil bo'ladi.
Ikki membranali organellalar (mitoxondriya va plastidlar)
Bu organellalar yarim avtonomdir, chunki ularning o'z DNKsi va oqsil sintez qiluvchi apparati mavjud. Mitoxondriyalar deyarli barcha eukaryotik hujayralarda uchraydi. Plastidlar faqat o'simlik hujayralarida uchraydi.
I) Mitoxondriya
Bular hujayradagi metabolik jarayonlar uchun energiya ta'minoti organellalari. Gialoplazmada mitoxondriyalar odatda diffuz tarzda taqsimlanadi, lekin ixtisoslashgan hujayralarda ular energiyaga eng katta ehtiyoj bo'lgan joylarda to'plangan. Masalan, mushak hujayralarida ko'p miqdorda mitoxondriyalar kontraktil fibrillalar bo'ylab, sperma flagellumi bo'ylab, buyrak kanalchalari epiteliysida, sinapslar sohasida va boshqalarda to'plangan. Mitoxondriyalarning bunday joylashishi ATPning kamroq yo'qolishini ta'minlaydi. uning tarqalishi paytida.
Tashqi membrana mitoxondriyalarni sitoplazmadan ajratib turadi, o'z-o'zidan yopiladi va invaginatsiyalar hosil qilmaydi. Ichki membrana mitoxondriyaning ichki tarkibini - matritsani cheklaydi. Xarakterli xususiyat - bu ko'plab invaginatsiyalar - kristallarning shakllanishi, buning natijasida ichki membranalar maydoni ortadi. Kristalarning soni va rivojlanish darajasi to'qimalarning funktsional faolligiga bog'liq. Mitoxondriya o'ziga xos genetik materialga ega (2.19-rasm).
Mitoxondrial DNK yopiq dumaloq ikki zanjirli molekula bo'lib, inson hujayralarida u 16569 juft nukleotid o'lchamiga ega, bu yadroda joylashgan DNKdan taxminan 105 marta kichikdir. Mitoxondriyalar o'zlarining oqsil sintez qilish tizimiga ega, mitoxondriyal mRNK dan tarjima qilingan oqsillar soni cheklangan. Mitoxondriyal DNK barcha mitoxondriyal oqsillarni kodlay olmaydi. Mitoxondriyadagi oqsillarning aksariyati yadroning genetik nazorati ostida.

Guruch. 2.19. Mitoxondriyal tuzilishi
Mitoxondrial funktsiyalar
1) ATP hosil bo'lishi
2) oqsil sintezi
3) o'ziga xos sintezlarda ishtirok etish, masalan, steroid gormonlar sintezi (buyrak usti bezlari)
4) sarflangan mitoxondriyalar ham ajralib chiqish mahsulotlarini, zararli moddalarni to'plashi mumkin, ya'ni. boshqa hujayra organellalarining funktsiyalarini bajarishga qodir
K) Plastidlar
Plastidlar-Organellalar, faqat o'simliklarga xosdir.
Plastidlarning uch turi mavjud:
1) xloroplastlar(yashil plastidlar);
2) xromoplastlar(sariq, to'q sariq yoki qizil plastidlar)
3) leykoplastlar(rangsiz plastidlar).
Odatda, hujayrada faqat bitta turdagi plastidlar mavjud.
Xloroplastlar
Bu organoidlar barglar va boshqa yashil o'simlik organlari hujayralarida, shuningdek, turli xil suv o'tlarida uchraydi. Yuqori o'simliklar odatda bir hujayrada bir necha o'nlab xloroplastlarni o'z ichiga oladi. Xloroplastlarning yashil rangi ulardagi xlorofill pigmentining tarkibiga bog'liq.
Xloroplast o'simlik hujayralarining asosiy organoidi bo'lib, unda fotosintez sodir bo'ladi, ya'ni quyosh nuri energiyasidan foydalangan holda noorganik (CO 2 va H 2 O) dan organik moddalar (uglevodlar) hosil bo'ladi. Xloroplastlar tuzilishi jihatidan mitoxondriyalarga o'xshaydi.
Xloroplastlar murakkab tuzilishga ega. Ular gialoplazmadan ikkita membrana bilan ajratilgan - tashqi va ichki. Ichki tarkib deyiladi stroma... Ichki membrana yassi pufakchalar ko'rinishidagi murakkab, qat'iy tartibga solingan membranalar tizimini hosil qiladi. tilakoidlar.
Tilakoidlar qoziqlarda to'planadi - donalar tangalar ustunlariga o'xshaydi . Grana bir-biri bilan stromal tilakoidlar orqali bog'langan bo'lib, ular orqali plastid bo'ylab o'tadi (2.20-2.22-rasm). Xlorofill va xloroplastlar faqat yorug'lik ta'sirida hosil bo'ladi.

Guruch. 2.20. Xloroplastlar yorug'lik mikroskopi ostida

Guruch. 2.21. Elektron mikroskop ostida xloroplast tuzilishi

Guruch. 2.22. Xloroplastlarning sxematik tuzilishi
Funksiyalar
1) fotosintez(yorug'lik energiyasi ta'sirida noorganik moddalardan organik moddalar hosil bo'lishi). Bu jarayonda xlorofil asosiy rol o'ynaydi. U yorug'lik energiyasini o'zlashtiradi va uni fotosintetik reaktsiyalarni amalga oshirishga yo'naltiradi. Xloroplastlarda, xuddi mitoxondriyadagi kabi, ATP sintezlanadi.
2) aminokislotalar va yog 'kislotalari sintezida ishtirok etish;
3) vaqtinchalik kraxmal zahiralari ombori bo'lib xizmat qiladi.
Leykoplastlar- quyosh nuridan yashiringan organlar hujayralarida (ildiz, ildizpoya, ildiz, urug') joylashgan mayda rangsiz plastidlar. Ularning tuzilishi xloroplastlarning tuzilishiga o'xshaydi. (2.23-rasm).
Biroq, xloroplastlardan farqli o'laroq, leykoplastlar yomon rivojlangan ichki membrana tizimiga ega, chunki ular zahiradagi oziq moddalar - kraxmal, oqsillar va lipidlarning sintezi va to'planishida ishtirok etadilar. Yorug'likda leykoplastlar xloroplastlarga aylanishi mumkin.

Guruch. 2.23. Leykoplast tuzilishi
Xromoplastlar- to'q sariq, qizil va sariq rangli plastidlar, ular karotinoidlar guruhiga kiruvchi pigmentlardan kelib chiqadi. Xromoplastlar ko'plab o'simliklarning gulbarglari hujayralarida, pishgan mevalarda, kamdan-kam ildiz ekinlarida, shuningdek, kuzgi barglarda uchraydi. Xromoplastlarda ichki membrana tizimi, qoida tariqasida, mavjud emas (24-rasm).

Guruch. 2.24. Xromoplast tuzilishi
Xromoplastlarning ahamiyati hali to'liq ochib berilmagan. Ularning aksariyati qarigan plastidlardir. Ular, qoida tariqasida, xloroplastlardan rivojlanadi, plastidlarda xlorofill va ichki membrana tuzilishi buziladi va karotenoidlar to'planadi. Bu kuzda mevalar pishib, barglari sarg'ayganida sodir bo'ladi. Xromoplastlarning biologik ahamiyati shundaki, ular gullar va mevalarning yorqin rangini aniqlaydi, bu esa hasharotlarni o'zaro changlatish uchun va boshqa hayvonlarni mevalarni yoyish uchun jalb qiladi. Leykoplastlar xromoplastlarga ham aylanishi mumkin.
Plastidlarning vazifalari
Oddiy noorganik birikmalardan xlorofilldagi organik moddalarning sintezi: karbonat angidrid va suv kvantlari quyosh nurlari ishtirokida - fotosintez, Fotosintezning yorug'lik bosqichida ATP sintezi
Ribosomalarda oqsil sintezi (xloroplastning ichki membranalari o'rtasida DNK, RNK va ribosomalar mavjud, shuning uchun xloroplastlarda, shuningdek, mitoxondriyalarda protein sintezlanadi, bu organellalarning faoliyati uchun zarurdir).
Xromoplastlarning mavjudligi gullar, mevalar, kuzgi barglarning sariq, to'q sariq va qizil ranglari bilan izohlanadi.
Leykoplastlarda saqlovchi moddalar mavjud (poyada, ildizda, ildizda).
Xloroplastlar, xromoplastlar va leykoplastlar hujayralararo o'tish qobiliyatiga ega. Shunday qilib, kuzda mevalar pishganda yoki barglarning rangi o'zgarganda, xloroplastlar xromoplastlarga, leykoplastlar esa xloroplastlarga aylanishi mumkin, masalan, kartoshka ildizlari yashil rangga aylanganda.
Evolyutsion ma'noda plastidlarning birlamchi, boshlang'ich turi xloroplastlar bo'lib, ulardan qolgan ikki turdagi plastidalar paydo bo'lgan. Plastidlar mitoxondriya bilan juda ko'p o'xshashliklarga ega bo'lib, ularni sitoplazmaning boshqa tarkibiy qismlaridan ajratib turadi. Bu, birinchi navbatda, ikkita membrananing qobig'i va nisbiy genetik avtonomiya, o'z ribosomalari va DNKlari mavjudligi sababli. Organoidlarning bu o'ziga xos xususiyati plastidalar va mitoxondriyalarning prekursorlari evolyutsiya jarayonida eukaryotik hujayraga aylanib, asta-sekin xloroplastlar va mitoxondriyalarga aylangan bakteriyalar ekanligi haqidagi g'oyaga asos bo'ldi. (2.25-rasm).

Guruch. 2.25. Simbiogenez nazariyasiga ko'ra mitoxondriya va xloroplastlarning shakllanishi
Hujayra- tirik tizimning elementar birligi. Tirik hujayraning muayyan funktsiyani bajarish uchun mas'ul bo'lgan turli tuzilmalari butun organizmning organlari kabi organellalar deb ataladi. Hujayradagi o'ziga xos funktsiyalar organellalar, ma'lum bir shaklga ega bo'lgan hujayra ichidagi tuzilmalar, masalan, hujayra yadrosi, mitoxondriyalar va boshqalar o'rtasida taqsimlanadi.
Hujayra tuzilmalari:
Sitoplazma... Plazma membranasi va yadro o'rtasida joylashgan hujayraning majburiy qismi. sitozol Turli tuzlar va organik moddalarning yopishqoq suvli eritmasi bo'lib, oqsil filamentlari tizimi - sitoskeleton orqali o'tadi. Hujayraning asosiy kimyoviy va fiziologik jarayonlari sitoplazmada sodir bo'ladi. Tuzilishi: sitozol, sitoskeleton. Vazifalari: turli organoidlarni, hujayraning ichki muhitini o'z ichiga oladi
Plazma membranasi... Hayvonlarning, o'simliklarning har bir hujayrasi atrof-muhitdan yoki boshqa hujayralardan plazma membranasi bilan chegaralangan. Ushbu membrananing qalinligi juda kichik (taxminan 10 nm), uni faqat elektron mikroskop bilan ko'rish mumkin.

Lipidlar ular membranada qo'sh qavat hosil qiladi va oqsillar uning butun qalinligidan o'tadi, lipid qatlamida turli chuqurliklarga botiriladi yoki membrananing tashqi va ichki yuzasida joylashgan. Boshqa barcha organellalar membranalarining tuzilishi plazma membranasiga o'xshaydi. Tuzilishi: ikki qavatli lipidlar, oqsillar, uglevodlar. Vazifalari: cheklash, hujayra shaklini saqlash, shikastlanishdan himoya qilish, moddalarni qabul qilish va olib tashlashni tartibga solish.
Lizosomalar... Lizosomalar membrana organellalaridir. Ular oval shaklga ega va diametri 0,5 mikron. Ularda organik moddalarni yo'q qiladigan fermentlar to'plami mavjud. Lizosoma membranasi juda kuchli bo'lib, o'z fermentlarining hujayra sitoplazmasiga kirib borishiga to'sqinlik qiladi, lekin lizosoma har qanday tashqi ta'sirlardan zarar ko'rsa, u holda butun hujayra yoki uning bir qismi vayron bo'ladi.
Lizosomalar o'simliklar, hayvonlar va zamburug'larning barcha hujayralarida mavjud.
Lizosomalar turli xil organik zarralarni hazm qilish orqali hujayradagi kimyoviy va energiya jarayonlari uchun qo'shimcha "xom ashyo" beradi. Ochlik paytida lizosoma hujayralari hujayrani o'ldirmasdan ba'zi organellalarni hazm qiladi. Bu qisman hazm qilish hujayrani ma'lum vaqt davomida kerakli minimal ozuqa moddalari bilan ta'minlaydi. Ba'zida lizosomalar hayvonlarning rivojlanishida muhim rol o'ynaydigan butun hujayralar va hujayralar guruhlarini hazm qiladi. Bunga misol qilib kurtak qurbaqaga aylanganda dumi yo'qolishi mumkin. Tuzilishi: ovalsimon pufakchalar, tashqi membrana, ichidagi fermentlar. Funktsiyalari: organik moddalarni bo'lish, o'lik organellalarni yo'q qilish, sarflangan hujayralarni yo'q qilish.
Golji kompleksi... Endoplazmatik retikulumning bo'shliqlari va tubulalarining lümenlerine kiradigan biosintez mahsulotlari Golji apparatida to'planadi va tashiladi. Ushbu organoidning o'lchami 5-10 mkm.

Tuzilishi: membrana bilan o'ralgan bo'shliqlar (vesikulalar). Vazifalari: organik moddalarning to'planishi, qadoqlanishi, chiqarilishi, lizosomalarning shakllanishi
Endoplazmatik retikulum... Endoplazmatik retikulum - bu hujayra sitoplazmasidagi organik moddalarni sintez qilish va tashish tizimi bo'lib, u bog'langan bo'shliqlarning ochiq tuzilishidir.
Ko'p sonli ribosomalar endoplazmatik retikulum membranalariga biriktirilgan - diametri 20 nm bo'lgan sharga o'xshash hujayraning eng kichik organellalari. va RNK va oqsildan iborat. Protein sintezi ribosomalarda sodir bo'ladi. Keyin yangi sintezlangan oqsillar bo'shliqlar va tubulalar tizimiga kiradi va ular bo'ylab hujayra ichida harakatlanadi. Membranlardan bo'shliqlar, tubulalar, tubulalar, ribosoma membranalari yuzasida. Vazifalari: ribosomalar yordamida organik moddalarni sintez qilish, moddalarni tashish.
Ribosomalar... Ribosomalar endoplazmatik retikulumning membranalariga biriktirilgan yoki sitoplazmada erkin joylashadi, ular guruhlarga bo'linadi, ularda oqsillar sintezlanadi. Protein tarkibi, ribosoma RNK vazifalari: oqsil biosintezini (oqsil molekulasining yig'ilishini) ta'minlaydi.
Mitoxondriya... Mitoxondriyalar energiya organellalaridir. Mitoxondriyalarning shakli har xil bo'lib, ular qolgan qismi, tayoqchali, o'rtacha diametri 1 mikron bo'lgan filamentli bo'lishi mumkin. va uzunligi 7 mikron. Mitoxondriyalar soni hujayraning funktsional faolligiga bog'liq va hasharotlarning uchuvchi mushaklarida o'n minglabgacha yetishi mumkin. Mitoxondriyalar tashqi tomondan tashqi membrana bilan chegaralangan, ostida ichki membrana joylashgan bo'lib, u ko'plab o'simtalar - kristalarni hosil qiladi.

Mitoxondriya ichida RNK, DNK va ribosomalar mavjud. Uning membranalarida o'ziga xos fermentlar mavjud bo'lib, ular yordamida oziq-ovqat moddalarining energiyasi mitoxondriyadagi hujayra va umuman tananing hayoti uchun zarur bo'lgan ATP energiyasiga aylanadi.
Membran, matritsa, o'simtalar - kristallar. Funktsiyalari: ATP molekulasining sintezi, o'z oqsillari, nuklein kislotalar, uglevodlar, lipidlar sintezi, o'z ribosomalarini hosil qilish.
Plastidlar... Faqat o'simlik hujayrasida: lekoplastlar, xloroplastlar, xromoplastlar. Funktsiyalari: zahiradagi organik moddalarni to'plash, changlatuvchi hasharotlarni jalb qilish, ATP va uglevodlarni sintez qilish. Xloroplastlar diametri 4-6 mkm bo'lgan disk yoki shar shaklida bo'ladi. Ikki qavatli membrana bilan - tashqi va ichki. Xloroplast ichida bir-biri bilan va xloroplastning ichki membranasi bilan bog'langan ribosoma DNKsi va maxsus membrana tuzilmalari - donalar mavjud. Har bir xloroplast 50 ga yaqin donani o'z ichiga oladi, ular yorug'likni yaxshiroq ushlab turish uchun stadlangan. Xlorofil gran membranalarda joylashgan bo'lib, uning yordamida quyosh nuri energiyasi ATP kimyoviy energiyasiga aylanadi. ATP energiyasi xloroplastlarda organik birikmalar, birinchi navbatda uglevodlar sintezi uchun ishlatiladi.
Xromoplastlar... Xromoplastlarda joylashgan qizil va sariq pigmentlar o'simlikning turli qismlariga qizil va sariq rang beradi. sabzi, pomidor mevalari.
Leykoplastlar zaxira ozuqa moddasi - kraxmal to'planish joyidir. Ayniqsa, kartoshka ildizlari hujayralarida leykoplastlar ko'p. Yorug'likda leykoplastlar xloroplastlarga aylanishi mumkin (buning natijasida kartoshka hujayralari yashil rangga aylanadi). Kuzda xloroplastlar xromoplastlarga, yashil barglar va mevalar sariq va qizil rangga aylanadi.
Hujayra markazi... Bir-biriga perpendikulyar joylashgan ikkita tsilindr, sentriollardan iborat. Funktsiyalari: shpindellarni ajratish uchun ipni qo'llab-quvvatlash

Hujayra inkluzyonlari sitoplazmada paydo bo'ladi, so'ngra hujayra hayoti jarayonida yo'qoladi.

Granulalar ko'rinishidagi zich qo'shimchalar zahiradagi ozuqa moddalarini (kraxmal, oqsillar, shakar, yog'lar) yoki hujayraning chiqindi mahsulotlarini o'z ichiga oladi, ularni hali olib tashlab bo'lmaydi. O'simlik hujayralarining barcha plastidlari zahiradagi ozuqa moddalarini sintez qilish va to'plash qobiliyatiga ega. O'simlik hujayralarida zahira ozuqa moddalarining to'planishi vakuolalarda sodir bo'ladi.
Donlar, granulalar, tomchilar Funktsiyalari: organik moddalar va energiyani saqlaydigan doimiy bo'lmagan shakllanishlar
Yadro... Ikki membrananing yadro membranasi, yadro shirasi, yadro. Vazifalari: hujayradagi irsiy axborotni saqlash va uni ko'paytirish, RNK sintezi - axborot, transport, ribosoma. Yadro membranasida sporalar mavjud bo'lib, ular orqali yadro va sitoplazma o'rtasida faol moddalar almashinuvi sodir bo'ladi. Yadro nafaqat ma'lum hujayraning barcha belgilari va xususiyatlari, unda sodir bo'lishi kerak bo'lgan jarayonlar (masalan, oqsil sintezi), balki butun organizmning xususiyatlari haqida ham irsiy ma'lumotlarni saqlaydi. Ma'lumot xromosomalarning asosiy qismi bo'lgan DNK molekulalarida qayd etiladi. Yadroda yadrocha mavjud. Yadro, unda irsiy ma'lumotni o'z ichiga olgan xromosomalar mavjudligi sababli, hujayraning barcha hayotiy faoliyati va rivojlanishini nazorat qiluvchi markaz funktsiyalarini bajaradi.