სენსორული სისტემების ფიზიოლოგია. სენსორული რეცეპტორები. ნერვული ბოჭკოები, ნეირომუსკულური სინაფსი რეცეპტორების როლი რეფლექსური რკალების ფორმირებაში
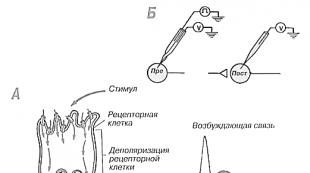
სენსორული რეცეპტორები არის სპეციალიზებული (ხშირად ნერვული) უჯრედები, რომლებიც პასუხისმგებელნი არიან ინფორმაციის გარდაქმნასა და გადაცემაზე. ნორმალური ნერვული უჯრედების მსგავსად, მათ აქვთ დენდრიტები და ერთი ან მეტი აქსონი. რეცეპტორები სპეციალიზირებულია გარემოს ენერგიის შესაბამისად.
რაზეც ისინი რეაგირებენ. Მაგალითად, ფოტორეცეპტორებიშეიცავს პიგმენტებს, რომლებიც ქიმიურად იცვლება სინათლის ზემოქმედებისას და სტიმულირებისას წარმოიქმნება ელექტრული პოტენციალი. IN მექანორცეპტორებიელექტროქიმიური ცვლილებები ხდება უჯრედის მემბრანის დეფორმაციის გამო. ენერგიის გარდაქმნა ჩვეულებრივ ხდება უჯრედის სხეულში და ყველა რეცეპტორისთვის დამახასიათებელია, რომ გარემოს ენერგია გარდაიქმნება თანდათანობით ელექტრულ პოტენციალად ე.წ. გენერატორის პოტენციალი,რომელიც ჩვეულებრივ პროპორციულია რეცეპტორების სტიმულაციის ინტენსივობისა. როდესაც გენერატორის პოტენციალი მიაღწევს გარკვეულ ზღვარს, ის იწვევს მოქმედების პოტენციალს, რომელიც გადის გასწვრივ
რეცეპტორული უჯრედის აქსონი. ეს არის სენსორული პროცესის გადაცემის ნაწილი და ინფორმაცია, როგორც წესი, დაშიფრულია ისე, რომ რაც უფრო ძლიერია სტიმული, მით უფრო მაღალია მოქმედების პოტენციალის სიხშირე. სტიმულაციის არარსებობის შემთხვევაში, გენერატორის პოტენციალი თანდათან მცირდება დასვენების დონემდე. როდესაც ის ზღურბლს ქვემოთ ეცემა, მოქმედების პოტენციალი წყვეტს გენერირებას. როდესაც სტიმულაცია განახლდება, შეიძლება იყოს მცირე შეფერხება (ლატენტური პერიოდი) მაშინ, როდესაც გენერატორის პოტენციალი დასვენებიდან ზღურბლამდე იზრდება. წყვეტილი სტიმულირებისას ის რიტმულად ადის და ეცემა, წარმოქმნის მოქმედების პოტენციალს. თუმცა, თუ წყვეტილი სტიმულაციის სიხშირე საკმარისად მაღალია, გენერატორის პოტენციალს შეიძლება არ ჰქონდეს დრო, რომ შემცირდეს სტიმულებს შორის და შემდეგ მოქმედების პოტენციალის წარმოქმნა უწყვეტი გახდება. ეს განმარტავს, თუ რატომ, წყვეტილი სტიმულაციის ძალიან მაღალი სიხშირით, ჩვენ არ შეგვიძლია განვასხვავოთ იგი უწყვეტი სტიმულაციისგან. ციმციმის შერწყმის ეს ფენომენი თანდაყოლილია ყველა გრძნობისთვის, რაც ყველაზე აშკარაა მხედველობის შემთხვევაში. ის ფაქტი, რომ სწრაფად მბჟუტავი შუქი წარმოქმნის იგივე ვიზუალურ შეგრძნებას, როგორც მუდმივი სინათლე, შესაძლებელს ხდის ტელევიზიას და კინოს.
მოქმედების პოტენციალი, რომელიც გადმოსცემს სენსორულ ინფორმაციას, არაფრით განსხვავდება სხვა ნერვული იმპულსებისგან. მათი სიდიდე განისაზღვრება აქსონის ზომით, ხოლო სიხშირე განისაზღვრება სტიმულაციის სიძლიერით. რეცეპტორის თითოეული ტიპი პირდაპირ ან ირიბად აგზავნის იმპულსებს ტვინის კონკრეტულ ნაწილზე. განცდილი შეგრძნებები დამოკიდებულია არა რეცეპტორის ტიპზე ან მის მიერ გაგზავნილ შეტყობინებებზე, არამედ ნაწილზე
ტვინი, რომელიც იღებს ამ შეტყობინებებს. შეგრძნების ლოკალიზაცია ასევე დამოკიდებულია ტვინზე. ასე, მაგალითად, ტკივილის დროს ნერვული ბოჭკოები ხელიდან აგზავნიან სიგნალებს ტვინის ერთ ნაწილზე, წინამხრიდან მეორეზე და ა.შ. ტვინის მიერ განცდილი „ტკივილი“ ლოკალიზებულია სხეულის იმ ნაწილში, საიდანაც შეტყობინება მოვიდა. ეს ფენომენი ილუსტრირებულია იმ ადამიანების ანგარიშებით, რომლებმაც გაიარეს კიდურების ამპუტაცია. რომლებიც უჩივიან ტკივილს, რომელიც თითქოს შორეული (ფანტომური) კიდურიდან მოდის. მოჭრილი ნერვული დაბოლოებების სტიმულირება აგზავნის იმპულსებს ტვინის იმ ნაწილებში, რომლებიც დაკავშირებული იყო ამპუტაციურ კიდურთან. ტვინი აღიქვამს შემომავალ სიგნალებს, როგორც დაკარგული კიდურებიდან მომდინარეობს და შედეგად მიღებული შეგრძნებები დამოკიდებულია იმაზე, თუ რომელი ნერვი გაღიზიანებულია. ამ ფანტომურმა კიდურმა ასევე შეიძლება გამოიწვიოს სითბოს, სიცივის ან შეხების შეგრძნება.
კუნთები და ჯირკვლები
ნერვული სისტემა აკონტროლებს ცხოველის ქცევას და გარკვეულწილად შინაგან გარემოს (იხ. თავი 15). ეს კონტროლი ხორციელდება კუნთებსა და ჯირკვლებზე მიცემული ბრძანებებით.
კუნთოვანი უჯრედები შეიცავს კომპლექსურ ცილის მოლეკულებს, რომლებსაც შეუძლიათ შეკუმშვა და მოდუნება. ნერვული დაბოლოებები უკავშირდება კუნთებს სინაფსების მეშვეობით, რომლებიც მსგავსია ნეირონების მიერ. ნეირომუსკულარულ შეერთებაზე მისვლისას ნერვული იმპულსები წარმოქმნიან ელექტრულ პოტენციალს, რაც იწვევს კუნთების შეკუმშვას. მისი მოდუნება ხდება სტიმულაციის არარსებობის შემთხვევაში. შეკუმშვისას კუნთი მცირდება, თუ ეს არ არის აღკვეთილი მისი ორივე ბოლოზე დაჭერით. მოდუნებისას კუნთს შეუძლია გახანგრძლივდეს, მაგრამ მხოლოდ იმ შემთხვევაში, თუ ის დაჭიმულია სხვა კუნთებით ან რაიმე გარე ძალით. კუნთები ჩვეულებრივ განლაგებულია ანტაგონისტურ, დაპირისპირებულ ჯგუფებად. ზოგიერთ უხერხემლოში, როგორიცაა ანელიდები, კუნთების შეკუმშვა შეიძლება შეფერხდეს ჰიდროსტატიკური გზით
ლოგიკური წნევა, რომელიც იზრდება სხეულის ღრუს კუნთოვანი ნაწილის შეკუმშვისას. ეს წნევა იწვევს კუნთების გახანგრძლივებას, როდესაც ისინი მოდუნდებიან. სხვა უხერხემლოებში, როგორიცაა ართროპოდები, კუნთები განლაგებულია ხისტი ეგზოჩონჩხის შიგნით, რომელიც ქმნის არა-
ბერკეტების აუცილებელი სისტემა კუნთების ანტაგონისტური ჯგუფებისთვის (სურ. 11.5). ხერხემლიანებში ასეთი სისტემაა შიდა ჩონჩხი და კუნთები განლაგებულია ისე, რომ მის ნაწილებს საპირისპირო მიმართულებით უქაჩონ (სურ. 11.6). კუნთების ერთი ჯგუფი მოდუნდება, როდესაც მეორე იკუმშება.
ზოგიერთი ჯირკვალი ნერვული კონტროლის ქვეშ იმყოფება. ხერხემლიანებში ეს მოიცავს, მაგალითად, სანერწყვე ჯირკვლებს, თირკმელზედა ჯირკვლის ტვინს, რომელიც გამოიმუშავებს ადრენალინს და უკანა ჰიპოფიზის ჯირკვალს, რომელიც გამოიმუშავებს რამდენიმე მნიშვნელოვან ჰორმონს. ამ ჯირკვლების სეკრეციას შეუძლია ირიბად გავლენა მოახდინოს ქცევაზე ცხოველის შინაგან მდგომარეობაზე ზემოქმედებით, როგორც ეს ნაჩვენები იქნება ამ თავის ბოლოს.
მათი სტრუქტურული ორგანიზაციისა და ფუნქციიდან გამომდინარე, სენსორული რეცეპტორები შეიძლება იყოს პირველადი ან მეორადი სენსორული. პირველადი სენსორული რეცეპტორები- ეს არის სენსორული ნეირონების პროცესების ნერვული დაბოლოებები. ისინი გვხვდება კანსა და ლორწოვან გარსებში, ჩონჩხის კუნთებში, მყესებში და პერიოსტეუმში, აგრეთვე შინაგანი გარემოს ბარიერულ სტრუქტურებში - სისხლისა და ლიმფური სისხლძარღვების კედლებში, ინტერსტიციულ სივრცეში, თავის ტვინისა და ზურგის ტვინის გარსებში და ცერებროსპინალური სითხის სისტემა. აღქმული სტიმულის ბუნებიდან გამომდინარე, პირველადი სენსორული რეცეპტორები იყოფა:
მექანორეცეპტორები (გაჭიმვის ან შეკუმშვის აღქმა, ქსოვილის ხაზოვანი ან რადიალური გადანაცვლება);
ქიმიორეცეპტორები (ქიმიური სტიმულის აღქმა);
თერმორეცეპტორები (ტემპერატურის აღქმა);
nociceptors (ტკივილის აღქმა).
მეორადი სენსორული რეცეპტორები- ეს არის რეცეპტორული უჯრედები, რომლებიც სპეციალიზირებულია გარკვეული სტიმულის აღქმაზე, ჩვეულებრივ ეპითელური ბუნების, რომლებიც გრძნობათა ორგანოების ნაწილია - ხედვა, სმენა, გემო, წონასწორობა. სტიმულის აღქმის შემდეგ, ეს რეცეპტორული უჯრედები ინფორმაციას გადასცემენ სენსორული ნეირონების აფერენტული გამტარების დაბოლოებებს. ამრიგად, ნერვული სისტემის აფერენტული ნეირონები იღებენ ინფორმაციას რეცეპტორულ უჯრედებში უკვე დამუშავებული სტიმულის შესახებ (რომელმაც განსაზღვრა ამ რეცეპტორების სახელწოდება).
ყველა ტიპის რეცეპტორები, აღქმული გაღიზიანების წყაროდან გამომდინარე, იყოფა ექსტერორეცეპტორები(გარე გარემოდან სტიმულების აღქმა) და ინტერრეცეპტორები(განკუთვნილია შინაგანი გარემოს გამღიზიანებლებისთვის). ინტერრეცეპტორებს შორის გამოიყოფა პროპრიორეცეპტორები, ე.ი. კუნთოვანი სისტემის საკუთარი რეცეპტორები, სისხლძარღვების კედლებში განლაგებული ანგიორეცეპტორები და ქსოვილის რეცეპტორები ლოკალიზებული ინტერსტიციულ სივრცეში და უჯრედულ მიკროგარემოში. პროპრიორეცეპტორებს შორის განსაკუთრებული ადგილი უკავია კუნთების ნაკვთებს, რომლებიც წარმოადგენენ ფორმირებებს, რომლებიც რეაგირებენ კუნთების გაჭიმვაზე და შეუძლიათ შეცვალონ მათი მგრძნობელობა ცენტრალური ნერვული სისტემიდან მათკენ მიმავალი იმპულსების გავლენის ქვეშ. ეს რეცეპტორები მონაწილეობენ კუნთების ტონის რეგულირებაში
ყველა ტიპის სენსორული რეცეპტორების საერთო ფუნქციური თვისებაა ერთი ტიპის ენერგიის მეორეში გადაქცევის უნარი: მექანიკური, თერმული, მსუბუქი, ბგერა და ა.შ.
ვინაიდან რეცეპტორები სპეციალიზირებულია გარკვეული ტიპის სტიმულის აღქმაზე, მათი მგრძნობელობა ასეთი სტიმულის მიმართ ყველაზე დიდია. სტიმულის მინიმალურ ძალას, რომელსაც შეუძლია რეცეპტორის აგზნება, ეწოდება სტიმულირების აბსოლუტური ბარიერი. ამასთან დაკავშირებით, სტიმულები, რომელთა მიმართ რეცეპტორს აქვს ყველაზე მაღალი მგრძნობელობა, ე.ი. მინიმალური ზღვრული მნიშვნელობა ეწოდება ადეკვატურს. ამავდროულად, ზოგიერთ რეცეპტორს ასევე შეუძლია უპასუხოს სტიმულს, რომელიც არ შეესაბამება მათ სპეციალობას; ასეთი სტიმულის ბარიერი, რომელსაც არაადეკვატური ეწოდება, აღმოჩნდება ძალიან მაღალი და სტიმულის მნიშვნელოვანი ძალაა საჭირო რეცეპტორის აღგზნებისთვის.
თუ პირველადი სენსორული რეცეპტორები წარმოიქმნება ერთი სენსორული ნეირონის პროცესის ტოტებით, ისინი ქმნიან სენსორული ნეირონის მიმღებ ველს. როგორც წესი, რეცეპტორები ქმნიან სხვადასხვა სიმკვრივის მტევანებს ქსოვილებში. იმ შემთხვევებში, როდესაც ეს დაგროვება წარმოშობს სპეციფიკურ რეფლექსს, მათ უწოდებენ მიმღებ რეფლექსურ ველებს. თუ კლასტერი შეიცავს სხვადასხვა ტიპის სტიმულის რეცეპტორებს, რომლებიც წარმოქმნიან სხვადასხვა რეფლექსებს, მათ რეფლექსოგენურ ზონებს უწოდებენ. ამის მაგალითია სისხლძარღვთა რეფლექსოგენური ზონები, სადაც განლაგებულია მექანო- და ქიმიორეცეპტორები, რომელთა გაღიზიანება იწვევს გულ-სისხლძარღვთა, რესპირატორული და სხეულის სხვა სისტემების სხვადასხვა რეფლექსურ რეაქციებს.
ანალიზატორები (სენსორული სისტემები) შექმნილია იმისთვის, რომ სხეულს მიაწოდოს ინფორმაცია ჰაბიტატსა და სხეულის შიდა გარემოში მომხდარი ცვლილებების შესახებ.
ანალიზატორი - პერიფერიული, გამტარი და ცენტრალური ნერვული წარმონაქმნების ნაკრები, რომლებიც აღიქვამენ და აანალიზებენ სიგნალებს სხეულის გარე და შიდა გარემოში ცვლილებების შესახებ.
გრძნობის ორგანო - პერიფერიული წარმონაქმნი, რომელიც აღიქვამს და ნაწილობრივ აანალიზებს არსებული გარემო ფაქტორების ცვლილებებს.
რეცეპტორები (სენსორული) - სპეციალიზებული უჯრედების ჯგუფები, რომლებიც ლოკალიზებულია სენსორულ ორგანოში ან სხეულის შინაგან გარემოში, რომელსაც შეუძლია აღიქვას, გარდაქმნას და გადასცეს ინფორმაცია მოქმედი გარემო ფაქტორების შესახებ ცენტრალურ ნერვულ სისტემაში.
სენსორული სისტემა - პერიფერიული (რეცეპტორი, სენსორული ორგანო), კომუნიკაციური (გამტარი განყოფილება) და ცენტრალური ნერვული წარმონაქმნების სისტემა, რომელიც უზრუნველყოფს სხეულის ინფორმაციას შიდა და გარე გარემოში მომხდარი მოვლენების შესახებ და აკონტროლებს აღქმის სტრუქტურების მოქმედებას.
სენსორული სისტემების აფერენტული ბმული (რეცეპტორები და გზები) ყოველთვის ჭარბობს ეფერენტულს. სხეულის ნებისმიერ შერეულ ნერვში სენსორული ბოჭკოების რაოდენობა აღემატება საავტომობილო ბოჭკოების რაოდენობას, მაგალითად, საშოს ნერვში აფერენტული ბოჭკოების ნაწილი 92%-ია.
ფუნქციური პასუხისმგებლობის მიმართულებიდან გამომდინარე, ანალიზატორები იყოფა გარე და შიდა.
1. გარე ანალიზატორები გარე გარემოში ცვლილებების აღქმა და ანალიზი. მათ შორისაა ვიზუალური, სმენითი, ყნოსვითი, გემოთი, ტაქტილური და ტემპერატურის ანალიზატორები, რომელთა აქტივობა სუბიექტურად აღიქმება შეგრძნებების სახით.
2. შიდა (ვისცერული) ანალიზატორები სხეულის შიდა გარემოში ცვლილებების აღქმა და ანალიზი. ფიზიოლოგიურ დიაპაზონში შინაგანი გარემოს ინდიკატორების რყევებს, როგორც წესი, ადამიანი არ აღიქვამს შეგრძნებების სახით (სისხლის pH, მასში CO 2 შემცველობა, არტერიული წნევა და ა.შ.).
ანალიზატორის განყოფილებები.პავლოვის მიერ წამოყენებული იდეის თანახმად, ყველა ანალიზატორს აქვს სამი ფუნქციური და ანატომიური განყოფილება.
ანალიზატორის პერიფერიული განყოფილება წარმოდგენილია რეცეპტორული უჯრედებით. რეცეპტორებს ახასიათებთ სპეციფიკა, მოდალობა ან აქტიური ადეკვატური სტიმულის გარკვეული ტიპის ენერგიის აღქმის უნარი (მიულერის კანონი სპეციფიკური ენერგიის შესახებ).
ანალიზატორის დირიჟორის განყოფილება მოიცავს ცენტრალური ნერვული სისტემის ღეროვანი და სუბკორტიკალური სტრუქტურების აფერენტულ (პერიფერიულ) და შუალედურ ნეირონებს (მათ შორის პროცესებს). გამტარი გზები უზრუნველყოფს სიგნალების არადეკრემენტულ გადაცემას თავის ტვინის ქერქში შუალედური ანალიზით და ინფორმაციის დატკეპნით სინაფსებში ნერვული იმპულსების გადაცემის დროს.
ანალიზატორის ცენტრალური ან კორტიკალური განყოფილება, პავლოვის თანახმად, იგი შედგება ორი ზონისგან: ცენტრალური ("ბირთვი", პირველადი), წარმოდგენილია სპეციფიკური ნეირონებით, რომლებიც ამუშავებენ შემომავალ იმპულსებს ანალიზატორის გამტარი ნაწილიდან და პერიფერიული, მეორადი, სადაც ნეირონების პოპულაციები იღებენ აფერენტულ შეყვანას. სხვადასხვა ანალიზატორების პერიფერიული განყოფილებებიდან.
ანალიზატორების კორტიკალურ პროგნოზებს ასევე უწოდებენ "სენსორული უბნები".
ანალიზატორებს ახასიათებთ მაღალი მგრძნობელობა მოდალურად სპეციფიკური სტიმულის მოქმედების მიმართ, რომელიც განისაზღვრება არა მხოლოდ რეცეპტორების თვისებებით, არამედ კორტიკალური ერთეულის ფუნქციონირებით.
დადგენილია ანალიზატორების ფუნქციონირების ზოგიერთი კანონი.
ვებერის კანონი
დისკრიმინაციის ბარიერი PR=ΔI/I=კონსტ
აღწერს ზღვრის დამოკიდებულებას სტიმულის ინტენსივობის ზრდის სუბიექტური შეგრძნების წარმოქმნის სტიმულის სიძლიერეზე.
ლოგარითმული ფორმით, ეს დამოკიდებულება გამოხატულია ფეხნერის კანონით
შეგრძნების ინტენსივობა Estr.=Кlg (ΔI/I)
საინტერესოა, რომ ვებერ-ფეხნერის კანონით აღწერილი ნიმუშები ცნობილი იყო ხალხისთვის ჯერ კიდევ ძველ ეპოქაში. ასტრონომები, კერძოდ ჰიპარქესი, როდესაც ქმნიდნენ ვარსკვლავთა სიდიდის სკალას მათი სიკაშკაშის მიხედვით, იყენებდნენ ვიზუალურ დაკვირვებებს, ე.ი. თქვენი ვიზუალური ანალიზატორის თვისებები. ვარსკვლავების თანამედროვე კლასიფიკაცია მათი სიკაშკაშის მიხედვით ადასტურებს, რომ, ფაქტობრივად, ჰიპარქემ პირველმა დაადგინა ვიზუალური ანალიზატორის მუშაობის დროს შეგრძნებების განმასხვავებლად ზღურბლის დამოკიდებულების ლოგარითმული ბუნება, ამის გაცნობიერების გარეშე.
მრავალი ანალიზატორის აფერენტული რგოლის აქტივობის გაანალიზებისას, დაახლოებით იგივე ნიმუში ვლინდება. უფრო მოსახერხებელია მისი აღწერა სტივენსის დენის ფუნქციით
n=1 მნიშვნელობებისთვის სტივენსის ფუნქცია ჩვეულებრივი პირდაპირპროპორციული ფუნქციაა, მაგრამ n-სთვის<1 (как это бывает в большинстве внешних анализаторов) кривая, описывающая эту функциональную закономерность на графике, круто уходит вверх при небольших приростах аргумента. Это означает, что происходит уплотнение информации (фактически, математическое логарифмирование) на уровне рецепторов для передачи ее в сжатом виде в ЦНС.
ნებისმიერი ანალიზატორის სენსორული ორგანოების და აფერენტული ნაწილების ძირითადი ფუნქციები განისაზღვრება სენსორული რეცეპტორების თვისებებით.
სენსორულ ფიზიოლოგიაში რეცეპტორები იყოფა გარე (ექსტერორეცეპტორებად) და შიდა (ინტერორეცეპტორებად).
რეცეპტორები კლასიფიცირდება მოდალობის მიხედვით
მექანორეცეპტორები (თმის უჯრედები - ფონორეცეპტორები, პაცინის კორპუსკულები, ბარორეცეპტორები, დაჭიმვის რეცეპტორები, კუნთების ღეროები და ა.შ.)
თერმორეცეპტორები ( ლოკალიზებულია შინაგან ორგანოებსა და კანში)
ქიმიორეცეპტორები (გლუკორეცეპტორები, ოსმორეცეპტორები, ყნოსვითი და გემო სენსორული სისტემის რეცეპტორები)
ფოტორეცეპტორები (ღეროები და კონუსები, ინფრაწითელი დეტექტორი გველებში)
ელექტრორეცეპტორები (ზოგიერთ თევზსა და ამფიბიაში)
ტკივილის რეცეპტორები ან ნოციცეპტორები (ხშირად მულტიმოდალური რეცეპტორები და ქსოვილის იშემიური რეცეპტორები).
რეცეპტორების ყველაზე მნიშვნელოვანი თვისებაა მათი მაღალი შერჩევითი მგრძნობელობა ადეკვატური სტიმულის მოქმედების მიმართ, რაც უზრუნველყოფილია ქსოვილებში მათი ლოკალიზაციის სტრუქტურული თავისებურებებით და სატელიტური უჯრედების არსებობით. თანმხლები რეცეპტორული უჯრედების არსებობის მიხედვით, ყველა რეცეპტორი იყოფა ორ ჯგუფად.
1.პირველადი რეცეპტორები, ან პირველადი სენსორული რეცეპტორები. სტიმულის გამოვლენა ხდება უშუალოდ სენსორული ნეირონის ბოლოს, ლოკალიზებულია პერიფერიაზე (ნერვული უჯრედის სხეული შეიძლება იყოს შორს სტიმულის მოქმედების ადგილიდან, ზოგიერთ სენსორულ განგლიონში). რეცეპტორის პოტენციალიდა გენერატორის პოტენციალიწარმოიქმნება ერთ ნერვულ უჯრედში. მაგალითი - მექანორეცეპტორები, პროპრიორეცეპტორები, ნოციცეპტორები, ქიმიორეცეპტორები.
2.მეორადი რეცეპტორები, მეორადი სენსორული რეცეპტორები. სენსორული ნეირონის ბოლოსა და სტიმულის აღქმის ადგილს შორის არის დამხმარე მიმღები უჯრედი. რეცეპტორის პოტენციალიწარმოიქმნება მიმღებ სატელიტურ უჯრედში, ის სინაპტიკურად ააქტიურებს აფერენტულ ნეირონს, რომელშიც ის წარმოიქმნება. გენერატორის პოტენციალი. ამის მაგალითია ფოტორეცეპტორები, შიდა ყურის ფონორეცეპტორები.
რეცეპტორული მოქმედების მექანიზმები .
სტიმულის მოქმედება არ იწვევს ენერგიის გადაცემას რეცეპტორზე; სტიმულის მიმღებ უჯრედში ენერგია უკვე ინახება K + –Na + ATPase-ს მუშაობით. მხოლოდ სტიმული ფოტონის სახით, გარემოს ვიბრაცია ან ნივთიერების კონცენტრაციის ცვლილება იწვევს სიგნალის გადაცემას.
მიმღები უჯრედები ხასიათდება მოსვენების პოტენციალის არსებობით. უჯრედის პლაზმალემა ჰყოფს საპირისპირო მუხტის მქონე უბნებს. უარყოფითი მუხტი შიგნით და დადებითი მუხტი გარეთ, ინტერსტიციუმში, უზრუნველყოფს რეცეპტორის მზადყოფნას სიგნალის შესაქმნელად. ყველაზე ხშირად, სიგნალი, რომელსაც რეცეპტორული უჯრედი წარმოქმნის, არის რეცეპტორის პოტენციალი, ეს არის თანდათანობითი, ელექტროტონურად გამრავლების პოტენციალი, დეპოლარიზაციის ან ჰიპერპოლარიზაციის სახით. პოტენციალი მიზნად ისახავს შეტყობინებების გადაცემის ქიმიური კავშირის პროცესის დაწყებას სინაფსში, რომელიც წარმოიქმნება რეცეპტორული უჯრედის მიერ აფერენტულ ნეირონზე. კატიონები მოქმედებენ როგორც მუხტის მატარებლები უჯრედებში. გამოიყენება მემბრანების საკაბელო თვისებები.
ამისთვის პირველადი გრძნობებირეცეპტორები, მოვლენების თანმიმდევრობა აღწერილია შემდეგი სქემით.
3. რეცეპტორის პოტენციალის გავრცელება ელექტროტონური მეთოდით ნეირონის პერიფერიული პროცესის ყველაზე აგზნებად ზონაში.
4.მოქმედების პოტენციალის გენერაცია. RP=GP
ამისთვის მეორადირეცეპტორი
1. სტიმულის სპეციფიკური ურთიერთქმედება რეცეპტორულ მემბრანასთან მოლეკულურ დონეზე
2. მგრძნობიარე იონური არხების ლოკალიზაციის ადგილას რეცეპტორული პოტენციალის გაჩენა
3. რეცეპტორული პოტენციალის ელექტროტონული გზის გავრცელება სინაფსამდე ნეირონის პერიფერიული პროცესით
4. გადამცემის გამოშვება სინაფსზე
5. EPSP-ის (აგზნებადი პოსტსინაფსური პოტენციალის) წარმოქმნა აფერენტული ნეირონის მემბრანაში. EPSP=GP
6. GP-ის ელექტროტონური გავრცელება აფერენტული ნეირონის ყველაზე აგზნებად ზონაში
7. PD-ს წარმოქმნა და მისი გადაცემა ცენტრალურ ნერვულ სისტემაში.
რეცეპტორების (და ნებისმიერი ანალიზატორის აფერენტული რგოლის) მთავარი ფუნქციაა ანალოგური სიგნალების გარდაქმნა გარე ან შიდა გარემოდან (შუქი, ხმა, სითბო, წნევა) სიხშირის (ციფრულ) კოდში, რომელიც შეიძლება გადაიცეს, თუ შესაძლებელია, შეკუმშული ფორმით, მაგრამ ინფორმაციის კომპონენტის დამახინჯების გარეშე. ამრიგად, გარდაქმნილი სიგნალი ჰგავს მოქმედების პოტენციალების ერთობლიობას (იმპულსებს), რომლებიც მოჰყვება გარკვეული სიხშირით. მაგრამ არა მხოლოდ სიხშირე მნიშვნელოვანია. კოდის სასარგებლო ინფორმაცია შეიძლება იყოს იმპულსების რაოდენობა, პულსთაშორისი ინტერვალი, იმპულსების არსებობა ან მათი არარსებობა. ამ შემთხვევაში, ხშირად ინტერპულსის ინტერვალები, თუნდაც ერთ აფერენტულ გაგზავნაში, ან იმპულსების სერიაში, შეიძლება იყოს თანაბარი ან განსხვავებული. აფერენტული რგოლის პასუხის დამოკიდებულების ლოგარითმული ბუნება სტიმულის მოქმედებაზე და სენსორული ნეირონის ინტეგრაციული როლი შესაძლებელს ხდის დროის კოდის მინიმიზაციას და ინფორმაციის გადაცემას, რომელზეც სენსორული სისტემა პასუხისმგებელია ცენტრალურ ნერვულ სისტემაზე. დამახინჯების გარეშე.
გაღიზიანების სიძლიერეც და მისი ხარისხიც კოდირებულია. სტიმულის მდებარეობა ასევე კოდირებულია. არსებობს სხვადასხვა ჰიპოთეზა ტვინში აფერენტული სისტემებიდან მომდინარე ინფორმაციის ანალიზის პროცესების ორგანიზების შესახებ.
ჰიპოთეზა ეტიკეტირებული ხაზი, ანუ სპეციფიკურობის თეორია, პოსტულირებულია ნეირონების სპეციალური ანატომიურად განსაზღვრული ჯაჭვების, საკომუნიკაციო არხების არსებობაზე, რომლებიც სპეციფიკურია თითოეული მიმდინარე სტიმულისთვის.
მხარდამჭერები ნიმუშის ჰიპოთეზები, ან ინტენსივობით, ითვლება, რომ სტიმულის ხარისხის შესახებ ინფორმაციის გადაცემა დაშიფრულია რეცეპტორების მრავალი პოპულაციის აგზნების სივრცითი-დროებითი ნიმუშით (ნიმუში) და საბოლოო ანალიზი ხდება მხოლოდ ცერებრალური ქერქის დონეზე.
1.2.1. სენსორული რეცეპტორების სტრუქტურული და ფუნქციური მახასიათებლები
სენსორული რეცეპტორების თვისებები.რეცეპტორების აგზნებადობა ძალიან მაღალია, ის აღემატება უახლესი ტექნიკური მოწყობილობების მგრძნობელობას, რომლებიც ჩაწერენ შესაბამის სიგნალებს. კერძოდ, ბადურის ფოტორეცეპტორის აღსაძვრელად საკმარისია სინათლის 1-2 კვანტა, ყნოსვის რეცეპტორისთვის კი სუნიანი ნივთიერების ერთი მოლეკულა. თუმცა, ვისცერორეცეპტორების აგზნებადობა უფრო დაბალია, ვიდრე ექსტერორეცეპტორების. ტკივილის რეცეპტორებს, რომლებიც ადაპტირებულია საზიანო სტიმულის მოქმედებაზე რეაგირებისთვის, აქვთ დაბალი აგზნებადობა.
რეცეპტორის ადაპტაცია -ეს არის მათი აგზნებადობის დაქვეითება სტიმულის გახანგრძლივებული ზემოქმედების დროს, რაც გამოიხატება RP-ის ამპლიტუდის დაქვეითებით და, შედეგად, იმპულსების სიხშირით აფერენტულ ნერვულ ბოჭკოში. სტიმულის მოქმედების საწყის ეტაპზე მათ დამხმარე სტრუქტურებს შეუძლიათ მნიშვნელოვანი როლი შეასრულონ რეცეპტორების ადაპტაციაში. მაგალითად, ვიბრაციის რეცეპტორების სწრაფი ადაპტაცია (Pacinian corpuscles) განპირობებულია იმით, რომ მათი კაფსულა საშუალებას აძლევს სტიმულის მხოლოდ სწრაფად ცვალებად პარამეტრებს გადავიდეს ნერვულ დაბოლოებამდე და "გაფილტროს" მისი სტატიკური კომპონენტები. უნდა აღინიშნოს, რომ ფოტორეცეპტორების ტერმინი „ბნელი ადაპტაცია“ ნიშნავს მათი აგზნებადობის ზრდას. რეცეპტორების ადაპტაციის ერთ-ერთი მექანიზმია მასში Ca 2+-ის დაგროვება აგზნებისას, რაც ააქტიურებს Ca 2+-დამოკიდებულ კალიუმის არხებს; ამ არხებით უჯრედიდან K+-ის გამოყოფა ხელს უშლის მისი მემბრანის დეპოლარიზაციას და, შესაბამისად, RP-ის წარმოქმნას. აღმოჩენილია ბიოქიმიური რეაქციები, რომლებიც ბლოკავს RP-ს წარმოქმნას. რეცეპტორების ადაპტაციის მნიშვნელობა ის არის, რომ ის იცავს სხეულს იმპულსების გადაჭარბებული ნაკადისგან, ზოგჯერ კი უსიამოვნო შეგრძნებებისგან.
სპონტანური აქტივობაზოგიერთი რეცეპტორები (ფონო-, ვესტიბულო-, თერმო-, ქიმიო- და პროპრიორეცეპტორები) მათზე გამაღიზიანებლის მოქმედების გარეშე, რაც დაკავშირებულია უჯრედის მემბრანის იონების გამტარიანობასთან, რაც პერიოდულად იწვევს PP-ს და CP-მდე დაქვეითებას. AP-ის წარმოქმნა ნერვულ ბოჭკოში. ასეთი რეცეპტორების აგზნებადობა უფრო მაღალია, ვიდრე რეცეპტორების ფონის აქტივობის გარეშე; სუსტ სტიმულსაც კი შეუძლია მნიშვნელოვნად გაზარდოს ნეირონის გასროლის სიჩქარე. რეცეპტორების ფონური აქტივობა ფიზიოლოგიური დასვენების პირობებში მონაწილეობს ცენტრალური ნერვული სისტემის ტონუსის შენარჩუნებაში და სხეულის სიფხიზლის მდგომარეობაში.
სენსორული რეცეპტორების ფუნქცია(ლათ. სენსუსი- განცდა, რეცეპტუმი-მიღება) არის სტიმულის აღქმა - ცვლილებები სხეულის გარე და შიდა გარემოში. ეს მიიღწევა სტიმულაციის ენერგიის RP-ში გადაქცევით, რაც უზრუნველყოფს ნერვული იმპულსების გაჩენას.
ევოლუციის პროცესში მყოფი რეცეპტორის თითოეული ტიპი ადაპტირებულია ერთი ან რამდენიმე ტიპის სტიმულის აღქმაზე. ასეთ სტიმულებს ე.წ ადეკვატური. მათ მიმართ ყველაზე დიდი მგრძნობელობა აქვთ რეცეპტორებს (მაგალითად, თვალის ბადურის რეცეპტორები აღფრთოვანებულია სინათლის ენერგიის 1-2 კვანტის მოქმედებით). Სხვებთან - არაადეკვატური სტიმული- რეცეპტორები არ არის მგრძნობიარე. შეუსაბამო სტიმულს შეუძლია სენსორული რეცეპტორების აღგზნებაც, მაგრამ ამ სტიმულების ენერგია მილიონობით და მილიარდჯერ მეტი უნდა იყოს, ვიდრე ადეკვატურის ენერგია. სენსორული რეცეპტორები არის პირველი რგოლი რეფლექსურ გზასა და სენსორული სისტემების პერიფერიულ ნაწილზე.
სენსორული რეცეპტორების კლასიფიკაციახორციელდება რამდენიმე კრიტერიუმის მიხედვით (სურ. 12).
ბრინჯი. 12. რეცეპტორების კლასიფიკაცია პირველად და მეორად. მეორად რეცეპტორებს აქვთ რეცეპტორული უჯრედი, რომელსაც უახლოვდება სენსორული ნეირონის აფერენტული დაბოლოებები (Agajanyan, 2007).
სტრუქტურული და ფუნქციური ორგანიზაციის მიხედვითგანასხვავებენ პირველადიდა მეორადირეცეპტორები.
პირველადი რეცეპტორებიწარმოადგენს აფერენტული ნეირონის დენდრიტის სენსორულ დაბოლოებებს. მათ შორისაა ყნოსვითი, ტაქტილური, ტემპერატურის, ტკივილის რეცეპტორები და პროპრიორეცეპტორები. ნეირონის სხეული მდებარეობს ზურგის განგლიაში ან კრანიალური ნერვების განგლიაში.
მეორადი რეცეპტორებიაქვთ სპეციალური უჯრედი სინაფსურად დაკავშირებული სენსორული ნეირონის დენდრიტის ბოლოსთან. მეორადი რეცეპტორები მოიცავს გემოს, ფოტოს (ვიზუალურს), ფონოს (სმენის) და ვესტიბულორეცეპტორებს.
ადაპტაციის სიჩქარითგანასხვავებენ სწრაფად ადაპტირებადი (ფაზური), ნელა ადაპტირებადი (მატონიზირებელი) და შერეული (ფაზურ-მატონიზირებელი) რეცეპტორები, ადაპტირება საშუალო სიჩქარით.სწრაფი ადაპტაციის რეცეპტორების მაგალითია კანის ვიბრაციის (Pacini corpuscles) და შეხების (Meissner corpuscles) რეცეპტორები. ნელ-ნელა ადაპტირებადი რეცეპტორები მოიცავს პროპრიორეცეპტორებს, ზოგიერთ ტკივილის რეცეპტორს და ფილტვების მექანიკურ რეცეპტორებს. ბადურის ფოტორეცეპტორები და კანის თერმორეცეპტორები ადაპტირდება საშუალო სიჩქარით.
აღქმული სტიმულის ტიპის მიხედვითგამოყოფს ოთხი ტიპირეცეპტორები, კერძოდ: ქიმიორეცეპტორები- გემოსა და ყნოსვის რეცეპტორები, სისხლძარღვთა და ქსოვილის რეცეპტორების ნაწილი (პასუხისმგებელია სისხლის, ლიმფის, უჯრედშორისი სითხის ქიმიური შემადგენლობის ცვლილებებზე) - იმყოფება ჰიპოთალამუსში (მაგალითად, კვების ცენტრში) და მედულას გრძივი (რესპირატორული) ცენტრი); მექანორცეპტორები- განლაგებულია კანსა და ლორწოვან გარსებში, ძვალ-კუნთოვან სისტემაში, სისხლძარღვებში, შინაგან ორგანოებში, სმენის, ვესტიბულურ და ტაქტილურ სენსორულ სისტემებში; თერმორეცეპტორები(იყოფენ სიცხესა და სიცივედ) - გვხვდება კანში, სისხლძარღვებში, შინაგან ორგანოებში, ცენტრალური ნერვული სისტემის სხვადასხვა ნაწილში (ჰიპოთალამუსი, შუა, მედულა და ზურგის ტვინი); ფოტორეცეპტორები- თვალის ბადურაზე განლაგებული, ისინი აღიქვამენ სინათლის (ელექტრომაგნიტურ) ენერგიას.
უნარიდან გამომდინარე აღიქვამენ ერთი ან რამდენიმე სახის სტიმულსგამოყოფს მონოსენსორული(აქვს მაქსიმალური მგრძნობელობა ერთი ტიპის სტიმულის მიმართ, მაგალითად, ბადურის რეცეპტორების მიმართ) და პოლისენსორული(რამდენიმე ადეკვატური სტიმულის აღქმა, მაგალითად მექანიკური და ტემპერატურის ან მექანიკური, ქიმიური და ტკივილის) რეცეპტორები. მაგალითად არის ფილტვების გამაღიზიანებელი რეცეპტორები, ტკივილის რეცეპტორები.
სხეულში მდებარეობის მიხედვითრეცეპტორები იყოფა გარე -და ინტერრეცეპტორები. TO ინტერრეცეპტორებიმოიცავს შინაგანი ორგანოების (ვისცერორეცეპტორების), სისხლძარღვების და ცენტრალური ნერვული სისტემის რეცეპტორებს. მრავალფეროვანი ინტერრეცეპტორები არის კუნთოვანი სისტემის რეცეპტორები (პროპრიორეცეპტორები) და ვესტიბულური რეცეპტორები. TO ექსტერორეცეპტორებიმათ შორისაა კანის რეცეპტორები, ხილული ლორწოვანი გარსები (მაგალითად, პირის ღრუს ლორწოვანი გარსი) და სენსორული ორგანოები: ვიზუალური, აუდიტორია, გემო, თერმორეცეპტორები, ყნოსვა.
იგრძნობა რეცეპტორებიდაყოფილია ვიზუალური, სმენის, გემოვნების, ყნოსვის თერმორეცეპტორები, ტაქტილური, ტკივილი(nociceptors) არის თავისუფალი ნერვული დაბოლოებები, რომლებიც გვხვდება კბილებში, კანში, კუნთებში, სისხლძარღვებსა და შინაგან ორგანოებში. მათ აღელვებს მექანიკური, თერმული და ქიმიური (ჰისტამინი, ბრადიკინინი, K+, H+ და სხვ.) სტიმულის მოქმედებით.
რეცეპტორის აგზნების მექანიზმი(სურ. 13).

ბრინჯი. 13. რეცეპტორული უჯრედიდან სიგნალის გაჩენისა და გადაცემის მექანიზმი (ჩესნოკოვა, 2007)
როდესაც ექვემდებარება ადეკვატურ სტიმულს პირველადი რეცეპტორიწარმოიქმნება რეცეპტორული პოტენციალი (RP), რაც წარმოადგენს უჯრედის მემბრანის დეპოლარიზაციას, ჩვეულებრივ, უჯრედში Na + იონების გადაადგილების გამო. RP არის ლოკალური პოტენციალი, ის არის ნერვული დაბოლოების გამაღიზიანებელი (ელექტრული ველის გამო) და უზრუნველყოფს AP-ის გაჩენას პულპალურ ბოჭკოებში - Ranvier-ის პირველ კვანძში, არაპულპურ ბოჭკოებში - უშუალო სიახლოვეს. რეცეპტორის.
In მეორადი რეცეპტორებისტიმულის ზემოქმედებისას, RP ასევე პირველად ჩნდება რეცეპტორულ უჯრედში Na + უჯრედში გადაადგილების გამო (გემოვნების კვირტები) ან K + (სმენა და ვესტიბულური რეცეპტორები).
RP-ის გავლენით, შუამავალი გამოიყოფა სინაფსურ ნაპრალში, რომელიც მოქმედებს პოსტსინაფსურ მემბრანაზე, უზრუნველყოფს GP-ის გენერატორის პოტენციალის ფორმირებას (ასევე ლოკალური).
ეს უკანასკნელი არის სტიმული (ელექტრული ველი), რომელიც უზრუნველყოფს AP-ის გაჩენას ნერვულ დაბოლოებაში, ასევე დაბოლოებებში პირველადი რეცეპტორებით.
AP სიხშირის დამოკიდებულება აფერენტულ ნერვულ ბოჭკოში RP მნიშვნელობაზე ნაჩვენებია ნახ. 14.

ბრინჯი. 14. ტიპიური ურთიერთობები RP-ის ამპლიტუდასა და AP-ების სიხშირეს შორის, რომლებიც წარმოიქმნება ეფერენტულ ნერვულ ბოჭკოში RP-ის ზღურბლზე ზემოთ (Guyton, 2008)
სენსორული რეცეპტორების კონცეფცია.პერიფერიული სენსორული სისტემების მთავარი კომპონენტია რეცეპტორი. ეს არის უაღრესად სპეციალიზებული სტრუქტურა (პირველადი სენსორული რეცეპტორებისთვის ეს არის აფერენტული ნეირონის მოდიფიცირებული დენდრიტი, მეორადი სენსორული რეცეპტორებისთვის ეს არის სენსორული რეცეპტორული უჯრედი), რომელსაც შეუძლია აღიქვას ადეკვატური სტიმულის მოქმედება გარე ან შიდა გარემოდან. და საბოლოო ჯამში მისი ენერგიის გარდაქმნა მოქმედების პოტენციალებად - ნერვული სისტემის სპეციფიკური აქტივობა. აქვე უნდა გავიხსენოთ, რომ ფიზიოლოგიაში „რეცეპტორის“ ცნება (ლათინური geserio, gesertum - მიღება, მიღება) ორი მნიშვნელობით გამოიყენება. პირველ რიგში, მიუთითეთ უჯრედის მემბრანის ან ციტოზოლის სპეციფიკური ცილები, რომლებიც განკუთვნილია ჰორმონების, შუამავლების და სხვა ბიოლოგიურად აქტიური ნივთიერებების გამოსავლენად. ასეთ რეცეპტორებს ჩვეულებრივ უწოდებენ მემბრანულ, უჯრედულ ან ჰორმონალურ (მაგალითად, ალფა-ადრენერგულ რეცეპტორებს). მეორეც, რეცეპტორების დანიშვნა სენსორული სისტემის კომპონენტებად. ამ რეცეპტორებს ხშირად უწოდებენ სენსორულ რეცეპტორებს, ან სენსორულ რეცეპტორულ უჯრედებს.
რეცეპტორების კლასიფიკაცია.იმისდა მიხედვით, აღიქმება თუ არა სტიმული შიდა თუ გარე გარემოდან, ყველა სენსორული რეცეპტორი იყოფა: ექსტერორეცეპტორებიდა ინტერრეცეპტორები. ექსტერორეცეპტორები აღიქვამენ სიგნალებს გარე გარემოდან. მათ შორისაა ბადურის ფოტორეცეპტორები, კორტის ორგანოს ფონორეცეპტორები, ნახევარწრიული არხების და ვესტიბულური ჩანთების ვესტიბულორეცეპტორები, კანისა და ლორწოვანი გარსების ტაქტილური, ტემპერატურისა და ტკივილის რეცეპტორები, ენის გემოვნების კვირტები, ცხვირის ყნოსვითი რეცეპტორები. ინტერრეცეპტორებს შორის არის ვისცერორეცეპტორები, რომლებიც შექმნილია შინაგანი გარემოში ცვლილებების გამოსავლენად და პროპრეცეპტორები (კუნთების და სახსრების რეცეპტორები, ანუ კუნთოვანი სისტემის რეცეპტორები). ვისცერორეცეპტორები არის შინაგანი ორგანოებისა და სისხლძარღვების სხვადასხვა ქიმიო-, მექანო-, თერმო-, ბარორეცეპტორები, აგრეთვე ნოციცეპტორები.
გარემოსთან კონტაქტის ხასიათიდან გამომდინარე, ექსტერორეცეპტორები იყოფა შორეულიინფორმაციის მიღება სტიმულაციის წყაროდან დაშორებით (ვიზუალური, სმენითი და ყნოსვითი) და კონტაქტი- აღგზნებულია სტიმულთან პირდაპირი კონტაქტით (გემოთი, ტაქტილური).
აღქმული სტიმულის მოდალობის სახეობიდან გამომდინარე, ე.ი. სტიმულის ბუნებიდან გამომდინარე, რომელზეც რეცეპტორები ოპტიმალურად არის მორგებული, სენსორული რეცეპტორები იყოფა 6 ძირითად ჯგუფად.: მექანორცეპტორები, თერმორეცეპტორები, ქიმიორეცეპტორები, ფონორეცეპტორები, ნოციცეპტორები და ელექტრორეცეპტორები (ეს უკანასკნელი გვხვდება მხოლოდ ზოგიერთ თევზსა და ამფიბიაში).
მექანიკური რეცეპტორები ადაპტირებულია გამაღიზიანებელი სტიმულის მექანიკური ენერგიის აღქმაზე. ისინი შედიან სომატური (ტაქტილური), კუნთოვანი, სმენის, ვესტიბულური და ვისცერული სენსორული სისტემების, აგრეთვე (თევზებში და ამფიბიებში) გვერდითი ხაზის სენსორული სისტემის ნაწილი. თერმორეცეპტორები აღიქვამენ ტემპერატურის სტიმულაციას, ე.ი. მოლეკულური მოძრაობის ინტენსივობა და ტემპერატურის სენსორული სისტემის ნაწილია. ისინი წარმოდგენილია კანის, შინაგანი ორგანოების და ჰიპოთალამუსის თერმომგრძნობიარე ნეირონების სითბოს და სიცივის რეცეპტორებით. ქიმიორეცეპტორები მგრძნობიარეა სხვადასხვა ქიმიკატების მოქმედების მიმართ და წარმოადგენს გემოს, ყნოსვის და ვისცერული სენსორული სისტემების ნაწილს. ფოტორეცეპტორები გრძნობენ სინათლის ენერგიას და ქმნიან ვიზუალური სენსორული სისტემის საფუძველს. ტკივილის (ნოციცეპტორული) რეცეპტორები აღიქვამენ ტკივილის სტიმულს, მათ შორის მექანოციცეპტორებს - გადაჭარბებული მექანიკური სტიმულის მოქმედებას, ქემონოციცეპტორებს - სპეციფიკური ტკივილის შუამავლების მოქმედებას; ისინი არიან ნოციცეპციური სენსორული სისტემის საწყისი კომპონენტი. რამდენიმე თევზის და ამფიბიების გვერდითი ხაზის გამოვლენილი ელექტრორეცეპტორები მგრძნობიარეა ელექტრომაგნიტური რხევების მოქმედების მიმართ.
ხაზგასმით უნდა აღინიშნოს, რომ ევოლუციის პროცესში შეირჩა ის რეცეპტორები და შესაბამისი სენსორული სისტემები, რომლებიც თითოეულ ორგანიზმს აწვდიდნენ საკმარის ინფორმაციას მისი ნორმალური არსებობისა და გარე გარემოში ადაპტაციისთვის. ამასთან დაკავშირებით შეგვიძლია მოვიყვანოთ ხატოვანი ფრაზა (A.D. Nozdrachev et al., 1991): „ელექტრორეცეპტორები, რომლებიც არსებობს თევზებში, ადამიანებში არ არის ნაპოვნი; არ არსებობს რეცეპტორები, რომლებიც აღიქვამენ პირდაპირ ინფრაწითელ გამოსხივებას, როგორც ჭინჭრის ციება; ადამიანის თვალი არ აღიქვამს სინათლის პოლარიზაციას, როგორც ზოგიერთი მწერის თვალები, მისი ყური ვერ გრძნობს ულტრაბგერითი ვიბრაციების, როგორც ღამურების და მრავალი ღამის ძუძუმწოვრების სმენის აპარატი“. მაგრამ, ზოგადად, ადამიანებისთვის ხელმისაწვდომი სენსორული სისტემები საშუალებას აძლევს მას, უფრო წარმატებულად გამოიკვლიოს დედამიწა, ვიდრე ცხოველთა სამყაროს სხვა წარმომადგენლები.
წარმოდგენილი ორი კლასიფიკაციის გარდა, მნიშვნელოვანია ყველა სენსორული რეცეპტორების დაყოფა მათი სტრუქტურისა და აფერენტულ სენსორულ ნეირონთან ურთიერთობის მიხედვით ორ დიდ კლასად - პირველადი სენსორული (პირველადი) და მეორადი სენსორული (მეორადი) რეცეპტორები. ეს განსაზღვრავს რეცეპტორის შერჩევით მგრძნობელობას ადეკვატური სტიმულის მიმართ (მეორად სენსორებში ეს ბევრად აღემატება პირველად სენსორებში), ასევე გარე სიგნალის ენერგიის ტრანსფორმაციის თანმიმდევრობას ნეირონის მოქმედების პოტენციალად.
პირველადი სენსორული რეცეპტორები მოიცავს იმ რეცეპტორებს, რომლებიც წარმოადგენენ აფერენტული ნეირონის დენდრიტის მოდიფიცირებულ, სპეციალიზებულ დასასრულს. ეს ნიშნავს, რომ აფერენტული ნეირონი უშუალოდ (ანუ, პირველ რიგში) ურთიერთქმედებს გარე სტიმულთან. პირველადი სენსორული რეცეპტორები მოიცავს მექანორეცეპტორების გარკვეულ ტიპებს (კანისა და შინაგანი ორგანოების თავისუფალი ნერვული დაბოლოებები), ცივი და სითბოს თერმორეცეპტორები, ნოციცეპტორები, კუნთების შტრიხები, მყესების რეცეპტორები, სახსრების რეცეპტორები, ყნოსვითი რეცეპტორები.
მეორადი რეცეპტორები არის არანერვული წარმოშობის უჯრედები, რომლებიც სპეციალურად არის ადაპტირებული გარე სიგნალის აღქმისთვის, რომლებიც, ადეკვატური სტიმულის მოქმედების საპასუხოდ აღგზნებული, გადასცემს სიგნალს (ჩვეულებრივ, სინაფსიდან გადამცემის გათავისუფლებით) აფერენტული ნეირონის დენდრიტი. შესაბამისად, ამ შემთხვევაში ნეირონი აღიქვამს სტიმულს ირიბად, ირიბად (მეორადად) სენსორული რეცეპტორული უჯრედის (მიმღები უჯრედის) აგზნების გამო. მეორადი სენსორული რეცეპტორები მოიცავს კანში არსებულ მექანორეცეპტორების ბევრ ტიპს (მაგალითად, პაცინის კორპუსკულებს, მერკელის დისკებს, მაისნერის უჯრედებს), ფოტორეცეპტორებს, ფონორეცეპტორებს, ვესტიბულორეცეპტორებს, გემოვნების კვირტებს და ელექტრორეცეპტორებს თევზებსა და ამფიბიებში.
სენსორული რეცეპტორების ადაპტაცია.სენსორულ რეცეპტორებს შეუძლიათ ადაპტაცია, რაც მდგომარეობს იმაში, რომ სენსორულ რეცეპტორზე სტიმულის მუდმივი ზემოქმედებით, მისი აგზნება სუსტდება, ე.ი. მცირდება რეცეპტორების პოტენციალის სიდიდე, ისევე როგორც აფერენტული ნეირონის მიერ მოქმედების პოტენციალის წარმოქმნის სიხშირე. მსგავსი ფენომენი შეინიშნება ჰორმონის რეცეპტორების ურთიერთქმედების დროს. ამ შემთხვევაში მას დესენსიბილიზაციას უწოდებენ და ასოცირდება ქვედა დინების სიგნალის გადაცემის დარღვევასთან. სენსორული რეცეპტორების ადაპტაცია კიდევ უფრო რთულია. ერთის მხრივ, ეს დამოკიდებულია პროცესებზე, რომლებიც ხდება სენსორული სტიმულის ურთიერთქმედების ეტაპზე სენსორული რეცეპტორის „აქტიურ ცენტრთან“ (არსებითად, ეს არის დესენსიბილიზაციის ფენომენი). მეორეს მხრივ, რეცეპტორების ადაპტაცია ასოცირდება სენსორულ რეცეპტორთან მისასვლელი იმპულსების ნაკადთან, ეფერენტული ბოჭკოების მეშვეობით ტვინის ზემოდან მოთავსებული ნეირონებიდან (მათ შორის რეტიკულური წარმონაქმნის ნეირონებიდან), ე.ი. აქტიური პროცესია. გარკვეულწილად, ადაპტაცია შეიძლება განისაზღვროს პერიფერიული სენსორული სისტემის დამხმარე სტრუქტურების თვისებებითა და მდგომარეობით. ზოგადად, ადაპტაცია ვლინდება სენსორული სისტემის აბსოლუტური მგრძნობელობის დაქვეითებით და მატებით. სხვადასხვა რეცეპტისთვის ადაპტაციის სიჩქარე განსხვავებულია: ყველაზე დიდი ტაქტილური რეცეპტორებისთვის და ყველაზე მცირე ვესტიბულური და პროპრიორეცეპტორებისთვის. ტაქტილური რეცეპტორების ადაპტაციის მაღალი სიჩქარის წყალობით, ჩვენ სწრაფად ვწყვეტთ სათვალეების, საათების ან ტანსაცმლის შეგრძნებას, ხოლო კუნთების რეცეპტორების ადაპტაციის დაბალი სიჩქარის წყალობით, ჩვენ შეგვიძლია გავაკეთოთ ძალიან კოორდინირებული და ზუსტი მოძრაობები.
გარე სტიმულის ენერგიის რეცეპტორულ პოტენციალად გადაქცევის ძირითადი ეტაპები (სენსორული რეცეპტორების აგზნების მექანიზმები). სენსორული რეცეპტორების მორფოფუნქციური მახასიათებლების მთელი მრავალფეროვნებით, ამ პროცესის ზოგადი სქემა შეიძლება წარმოდგენილი იყოს ზოგიერთი განზოგადებული დიაგრამის სახით. IN პირველადი რეცეპტორებიპირობითად შეიძლება განვასხვავოთ სენსორული სიგნალის გადაცემის ხუთი ძირითადი ეტაპი: 1) აღქმული სტიმულის ურთიერთქმედება სენსორული რეცეპტორის „აქტიურ“ ნაწილთან; 2) მემბრანის იონური გამტარიანობის ცვლილება; 3) სენსორული რეცეპტორის მემბრანული პოტენციალის დონის დაქვეითება, ე.ი. რეცეპტორული პოტენციალის წარმოქმნა, რომლის დონე დამოკიდებულია აღქმული სტიმულის სიდიდეზე; 4) მოქმედების პოტენციალის წარმოქმნა ან სპონტანური მოქმედების პოტენციალის წარმოქმნის სიხშირის ზრდა აფერენტული ნეირონის სომაში (აქსონის ბორცვი); 5) აქსონის გასწვრივ მოქმედების პოტენციალის გავრცელება მოცემული სენსორული სისტემის მეორე აფერენტულ ნეირონამდე. In მეორადი გრძნობებისენსორულ უჯრედებში პირველი სამი სტადია ერთნაირი ნიმუშია; შემდეგ ემატება კიდევ ორი შუალედური ეტაპი - 4ა) რეცეპტორული უჯრედის სინაფსში მედიატორის კვანტების (მაგალითად, აცეტილქოლინის) გამოყოფა რეცეპტორული პოტენციალის გავლენით; 5ა) აფერენტული ნეირონის დენდრიტის რეაქცია გადამცემის გათავისუფლებაზე აგზნების პოსტსინაფსური პოტენციალის ან გენერატორის პოტენციალის წარმოქმნით. დანარჩენი ორი ეტაპი (4 და 5) მიმდინარეობს ისევე, როგორც პირველადი სენსორული რეცეპტორების დროს. ამ წესის ერთადერთი გამონაკლისი არის ვიზუალური სენსორული სისტემის მოვლენების ჯაჭვი, რომლის დროსაც, სინათლის მოქმედების საპასუხოდ, ფოტორეცეპტორული უჯრედი ზრდის მემბრანულ პოტენციალს, რის შედეგადაც მასში მცირდება ინჰიბიტორული გადამცემის წარმოება. , რაც საბოლოოდ იწვევს ბიპოლარული ნეირონის აგზნებას, რაც თავის მხრივ აღაგზნებს განგლიურ უჯრედს.