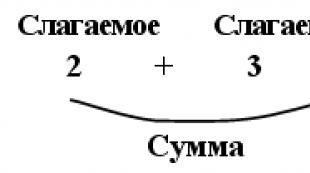
Տեսեք, թե ինչ են «Փոքր ՌՆԹ»-ները այլ բառարաններում։ Գենի արտահայտությունը և փոքր ՌՆԹ-ները ուռուցքաբանության մեջ Փոքր ՌՆԹ-ների գործառույթները
ՌՆԹ միջամտության երևույթի անվան հիմքում ընկած փոխաբերությունը վերաբերում է petunia-ի հետ փորձին, երբ գործարանում արհեստականորեն ներմուծված վարդագույն և մանուշակագույն պիգմենտների սինթետազային գեները չեն մեծացրել գույնի ինտենսիվությունը, այլ ընդհակառակը, կրճատել են այն: Նմանապես, «նորմալ» միջամտության դեպքում երկու ալիքների սուպերպոզիցիան կարող է հանգեցնել փոխադարձ «չեղարկման»:
Կենդանի բջջում միջուկի և ցիտոպլազմայի միջև տեղեկատվության հոսքը երբեք չի չորանում, սակայն դրա բոլոր «պտույտները» հասկանալը և դրանում կոդավորված տեղեկատվության վերծանումը իսկապես տիտանական խնդիր է: Անցյալ դարի կենսաբանության ամենակարևոր առաջընթացներից մեկը կարելի է համարել տեղեկատվական (կամ մատրիցային) ՌՆԹ-ի (mRNA կամ mRNA) մոլեկուլների հայտնաբերումը, որոնք ծառայում են որպես միջուկից (քրոմոսոմներից) տեղեկատվական «հաղորդագրություններ» փոխանցող միջնորդներ։ ցիտոպլազմ. ՌՆԹ-ի որոշիչ դերը սպիտակուցի սինթեզում կանխատեսվել էր դեռևս 1939 թվականին Տորբյորն Կասպերսոնի, Ժան Բրաշետի և Ջեք Շուլցի կողմից, իսկ 1971 թվականին Ջորջ Մարբեյսը սկսեց հեմոգլոբինի սինթեզը ձվաբջիջներում՝ ներարկելով այս ճագարի առաքիչ ՌՆԹ-ն կոդավորող առաջին մեկուսացված սպիտակուցը:
1956-57 թվականներին Խորհրդային Միությունում A.N.Belozersky-ը և A.S.Spirin-ը ինքնուրույն ապացուցեցին mRNA-ի գոյությունը, ինչպես նաև պարզեցին, որ բջջի ՌՆԹ-ի հիմնական մասը ոչ թե մատրիցա է, այլ ռիբոսոմային ՌՆԹ (rRNA): Ռիբոսոմային ՌՆԹ-ն՝ բջջային ՌՆԹ-ի երկրորդ «հիմնական» տեսակը, բոլոր օրգանիզմներում կազմում է ռիբոսոմների «կմախքը» և ֆունկցիոնալ կենտրոնը. դա rRNA (և ոչ սպիտակուցներ) է, որը կարգավորում է սպիտակուցների սինթեզի հիմնական փուլերը: Միաժամանակ նկարագրվել և ուսումնասիրվել է ՌՆԹ-ի երրորդ «հիմնական» տեսակը՝ տրանսպորտային ՌՆԹ-ն (tRNA), որը երկու այլ՝ mRNA-ի և rRNA-ի հետ միասին կազմում է մեկ սպիտակուցային սինթեզող համալիր։ Ըստ «ՌՆԹ աշխարհի» բավականին տարածված վարկածի, հենց այս նուկլեինաթթուն է ընկած Երկրի վրա կյանքի հենց սկզբնաղբյուրում:
Շնորհիվ այն բանի, որ ՌՆԹ-ն շատ ավելի հիդրոֆիլ է ԴՆԹ-ի համեմատ (դեզօքսիրիբոզան ռիբոզով փոխարինելու պատճառով), այն ավելի անկայուն է և կարող է համեմատաբար ազատ տեղաշարժվել բջջում և, հետևաբար, տրամադրել գենետիկ տեղեկատվության (mRNA) կարճատև կրկնօրինակներ: այնտեղ, որտեղ սկսվում է սպիտակուցի սինթեզը: Այնուամենայնիվ, հարկ է նշել սրա հետ կապված «անհարմարությունը»՝ ՌՆԹ-ն շատ անկայուն է։ Այն շատ ավելի վատ է, քան ԴՆԹ-ն, այն պահպանվում է (նույնիսկ բջջի ներսում) և քայքայվում պայմանների ամենաչնչին փոփոխության դեպքում (ջերմաստիճան, pH): Ի լրումն «սեփական» անկայունության, մեծ ներդրում ունեն ռիբոնուկլեազները (կամ RNases)՝ ՌՆԹ անջատող ֆերմենտների դասը, որոնք շատ կայուն են և «ամենուրեք տարածված»: ամբողջ փորձը. Դրա պատճառով ՌՆԹ-ի հետ աշխատելը շատ ավելի դժվար է, քան սպիտակուցների կամ ԴՆԹ-ի հետ. վերջիններս, ընդհանուր առմամբ, կարող են պահպանվել հարյուր հազարավոր տարիներ՝ քիչ կամ առանց վնասների:
Աշխատանքի մեջ ֆանտաստիկ ճշգրտություն, տրիդիսիլլատ, ստերիլ ձեռնոցներ, լաբորատոր ապակյա միանգամյա օգտագործման սպասք. այս ամենը անհրաժեշտ է ՌՆԹ-ի քայքայումը կանխելու համար, սակայն նման չափանիշներին համապատասխանելը միշտ չէ, որ հնարավոր է եղել: Ուստի երկար ժամանակ պարզապես անտեսվում էին ՌՆԹ-ի կարճ «բեկորները», որոնք անխուսափելիորեն աղտոտում էին լուծույթները։ Այնուամենայնիվ, ժամանակի ընթացքում պարզ դարձավ, որ, չնայած աշխատանքային տարածքի ստերիլությունը պահպանելու բոլոր ջանքերին, «բեկորները», բնականաբար, շարունակվեցին գտնել, և հետո պարզվեց, որ ցիտոպլազմայում միշտ կան հազարավոր կարճ երկշղթա ՌՆԹ: որոնք կատարում են բավականին կոնկրետ գործառույթներ և բացարձակապես անհրաժեշտ են բջիջների և օրգանիզմի նորմալ զարգացման համար։
ՌՆԹ-ի միջամտության սկզբունքը
Այսօր փոքր կարգավորող ՌՆԹ-ների ուսումնասիրությունը մոլեկուլային կենսաբանության ամենաարագ զարգացող ոլորտներից մեկն է: Պարզվել է, որ բոլոր կարճ ՌՆԹ-ները կատարում են իրենց գործառույթները ՌՆԹ-ի միջամտություն կոչվող երևույթի հիման վրա (այս երևույթի էությունը գենի արտահայտման ճնշումն է տրանսկրիպցիայի կամ թարգմանության փուլում՝ փոքր ՌՆԹ մոլեկուլների ակտիվ մասնակցությամբ): ՌՆԹ-ի միջամտության մեխանիզմը շատ սխեմատիկորեն ներկայացված է Նկար 1-ում:
Բրինձ. 1. ՌՆԹ-ի միջամտության հիմունքները
Երկաշղթա ՌՆԹ (dsRNA) մոլեկուլները սովորական բջիջներում հազվադեպ են, բայց դրանք շատ վիրուսների կյանքի ցիկլի կարևոր քայլ են: Հատուկ Dicer սպիտակուցը, հայտնաբերելով dsRNA բջջում, այն «կտրում» է փոքր բեկորների: Նման հատվածի հակազգայական շարանը, որն արդեն կարելի է անվանել կարճ միջամտող ՌՆԹ (siRNA, siRNA-ից՝ փոքր միջամտության ՌՆԹ), կապված է RISC կոչվող սպիտակուցների համալիրով (ՌՆԹ-ի պատճառած խլացման համալիր), որի կենտրոնական տարրն է. Արգոնավտների ընտանիքի էնդոնուկլեազ: ՍԻՌՆԹ-ին միանալը ակտիվացնում է RISC-ն և բջիջում սկսում է որոնել ԴՆԹ-ի և ՌՆԹ-ի մոլեկուլները, որոնք լրացնում են «կաղապար» siRNA-ին: Նման մոլեկուլների ճակատագիրը RISC համալիրի կողմից ոչնչացվելու կամ ապաակտիվանալու է:
Ամփոփելու համար, օտարերկրյա (ներառյալ միտումնավոր ներմուծված) երկշղթա ՌՆԹ-ի կարճ «գրությունները» ծառայում են որպես «կաղապար»՝ կոմպլեմենտար mRNA-ների լայնածավալ որոնման և ոչնչացման համար (և դա համարժեք է համապատասխան գենի արտահայտությունը ճնշելուն): եւ ոչ միայն մեկ խցում, այլեւ հարեւան։ Շատ օրգանիզմների՝ նախակենդանիների, փափկամարմինների, ճիճուների, միջատների, բույսերի համար այս երեւույթը վարակներից իմունային պաշտպանության հիմնական միջոցներից մեկն է:
2006 թվականին Էնդրյու Ֆայրը և Քրեյգ Մելլոն Նոբելյան մրցանակ են ստացել ֆիզիոլոգիայի կամ բժշկության բնագավառում՝ ՌՆԹ-ի միջամտության ֆենոմենի հայտնաբերման համար՝ dsRNA-ի մասնակցությամբ գեների լռեցման մեխանիզմը։ Թեև ՌՆԹ-ի միջամտության ֆենոմենն ինքնին նկարագրվել էր դրանից շատ առաջ (դեռևս 1980-ականների սկզբին), դա Fire-ի և Mello-ի աշխատանքն էր: ընդհանուր ուրվագիծորոշեց փոքր ՌՆԹ-ների կարգավորիչ մեխանիզմը և նախանշեց մոլեկուլային հետազոտության մինչ այժմ անհայտ տարածքը: Ահա նրանց աշխատանքի հիմնական արդյունքները.
- ՌՆԹ-ի ինտերֆերենսի դեպքում դա մՌՆԹ-ն է (և ոչ մի այլ), որը ճեղքվում է.
- Երկաշղթա ՌՆԹ-ն շատ ավելի արդյունավետ է գործում (առաջացնում է ճեղքվածք), քան միաշղթան: Այս երկու դիտարկումները կանխատեսում էին մասնագիտացված համակարգի առկայությունը, որը միջնորդում է dsRNA-ի գործողությունը.
- dsRNA-ն, որը կոմպլեմենտար է հասուն mRNA կայքին, հանգեցնում է վերջինիս ճեղքմանը: Սա ցույց տվեց գործընթացի ցիտոպլազմային տեղայնացումը և հատուկ էնդոնուկլեազի առկայությունը.
- dsRNA-ի փոքր քանակությունը (մեկ բջջում մի քանի մոլեկուլ) բավարար է թիրախ գենն ամբողջությամբ «անջատելու» համար, ինչը ցույց է տալիս կատալիզացման և/կամ ուժեղացման կասկադային մեխանիզմի առկայությունը:
Այս արդյունքները հիմք դրեցին ժամանակակից մոլեկուլային կենսաբանության մի ամբողջ ոլորտի՝ ՌՆԹ-ի միջամտության, և որոշեցին աշխարհի բազմաթիվ հետազոտական խմբերի աշխատանքի վեկտորը ավելի քան մեկ տասնյակ տարի: Մինչ այժմ հայտնաբերվել են փոքր ՌՆԹ-ների երեք մեծ խմբեր, որոնք խաղում են մոլեկուլային դաշտում «ՌՆԹ-ի միջամտության հրամանի» համար: Եկեք ավելի մանրամասն ճանաչենք նրանց։
Խաղացող # 1 - կարճ միջամտող ՌՆԹ-ներ
ՌՆԹ-ի միջամտության առանձնահատկությունը որոշվում է կարճ միջամտող ՌՆԹ-ներով (siRNAs)՝ հստակ հստակ կառուցվածքով փոքր երկշղթա ՌՆԹ-ի մոլեկուլներով (տես նկ. 2):
siRNA-ները էվոլյուցիոն առումով ամենավաղն են և առավել տարածված են բույսերի, միաբջիջ օրգանիզմների և անողնաշարավորների մեջ: Ողնաշարավորների մոտ siRNA-ները գործնականում սովորաբար չեն հայտնաբերվում, քանի որ դրանք փոխարինվել են կարճ ՌՆԹ-ների հետագա «մոդելներով» (տես ստորև):
siRNA-ն՝ ցիտոպլազմում փնտրելու և mRNA մոլեկուլները ոչնչացնելու «կաղապարներ», ունեն 20-25 նուկլեոտիդներ և ունեն հատուկ առանձնահատկություն՝ 2 չզույգված նուկլեոտիդներ 3 «ծայրերում և ֆոսֆորիլացված 5» ծայրերում: Հակազգայական siRNA-ն ի վիճակի է (իհարկե, ոչ ինքնին, այլ RISC-համալիրի օգնությամբ) ճանաչել mRNA-ն և հատուկ առաջացնել դրա քայքայումը. թիրախային mRNA-ի կտրվածքը միշտ տեղի է ունենում հենց այն տեղում, որը լրացնում է 10-րդը և Հակազգայական siRNA շղթայի 11-րդ նուկլեոտիդներ:

Բրինձ. 2. mRNA-ի և siRNA-ի «միջամտության» մեխանիզմը
«Խոչընդոտող» կարճ ՌՆԹ-ի մոլեկուլները կարող են և՛ դրսից ներթափանցել բջիջ, և՛ «կտրվել» արդեն տեղում՝ ավելի երկար երկշղթա ՌՆԹ-ներից: Հիմնական սպիտակուցը, որն անհրաժեշտ է dsRNA-ի կտրման համար, Dicer էնդոնուկլեազն է: Գենի «անջատումը» միջամտության մեխանիզմով իրականացվում է siRNA-ի կողմից RISC սպիտակուցային համալիրի հետ միասին, որը բաղկացած է երեք սպիտակուցներից՝ էնդոնուկլեազ Ago2 և երկու օժանդակ սպիտակուցներից՝ PACT և TRBP: Հետագայում պարզվեց, որ Dicer և RISC համալիրները որպես «պրայմեր» կարող են օգտագործել ոչ միայն dsRNA, այլ նաև միաշղթա ՌՆԹ, որը ձևավորում է երկշղթա մազակալ, ինչպես նաև պատրաստի siRNA (վերջինս շրջանցում է փուլը. «կտրում» և անմիջապես կապում է RISC-ին):
Անողնաշարավորների բջիջներում siRNA-ների գործառույթները բավականին բազմազան են։ Առաջինը և գլխավորը իմունային պաշտպանությունն է: «Ավանդական» իմունային համակարգը (լիմֆոցիտներ + լեյկոցիտներ + մակրոֆագեր) առկա է միայն բարդ բազմաբջիջ օրգանիզմներում։ Միաբջիջ օրգանիզմների, անողնաշարավորների և բույսերի մոտ (որոնք կամ չունեն նման համակարգ, կամ գտնվում են սաղմնային վիճակում), իմունային պաշտպանությունը հիմնված է ՌՆԹ միջամտության վրա։ ՌՆԹ-ի միջամտության վրա հիմնված անձեռնմխելիությունը բարդ օրգանների կարիք չունի իմունային բջիջների պրեկուրսորներին (փայծաղ, տիմուս) «մարզելու» համար. Միևնույն ժամանակ, տեսականորեն հնարավոր կարճ ՌՆԹ հաջորդականությունների բազմազանությունը (421 տարբերակ) համեմատելի է բարձրակարգ կենդանիների հնարավոր սպիտակուցային հակամարմինների քանակի հետ։ Բացի այդ, siRNA-ները սինթեզվում են «թշնամական» ՌՆԹ-ի հիման վրա, որը վարակել է բջիջը, ինչը նշանակում է, որ, ի տարբերություն հակամարմինների, դրանք անմիջապես «սրվում» են վարակի կոնկրետ տեսակի համար։ Ու թեև ՌՆԹ-ի միջամտության վրա հիմնված պաշտպանությունը բջջից դուրս չի գործում (համենայն դեպս նման տվյալներ դեռ չկան), այն ապահովում է ներբջջային իմունիտետ ավելի քան գոհացուցիչ։
Նախ և առաջ, siRNA-ն հակավիրուսային իմունիտետ է ստեղծում՝ ոչնչացնելով վարակիչ օրգանիզմների mRNA կամ գենոմային ՌՆԹ-ն (օրինակ՝ այսպես են հայտնաբերվել siRNA-ները բույսերում): Վիրուսային ՌՆԹ-ի ներմուծումը առաջացնում է հատուկ siRNA-ների հզոր ուժեղացում՝ հիմնված պրիմերի մոլեկուլի՝ հենց վիրուսային ՌՆԹ-ի վրա: Բացի այդ, siRNA-ները ճնշում են տարբեր շարժական գենետիկ տարրերի (MGE) արտահայտությունը, ինչը նշանակում է, որ նրանք պաշտպանում են էնդոգեն «վարակներից»: RISC համալիրի գեների մուտացիաները հաճախ հանգեցնում են գենոմի անկայունության բարձրացման՝ MGE-ի բարձր ակտիվության պատճառով; siRNA-ն կարող է սահմանափակել սեփական գեների արտահայտությունը՝ ի պատասխան դրանց գերարտահայտման: Գենների աշխատանքի կարգավորումը կարող է տեղի ունենալ ոչ միայն թարգմանության մակարդակում, այլ նաև տրանսկրիպցիայի ժամանակ՝ հիստոն H3-ում գեների մեթիլացման միջոցով:
Ժամանակակից փորձարարական կենսաբանության մեջ ՌՆԹ-ի միջամտության և կարճ ՌՆԹ-ների կարևորությունը դժվար թե կարելի է գերագնահատել: Մշակվել է առանձին գեների in vitro (բջջային կուլտուրաներում) և in vivo (սաղմերի մեջ) «անջատելու» (կամ նոկդաունի) տեխնոլոգիա, որն արդեն դարձել է ցանկացած գենի ուսումնասիրության դե ֆակտո ստանդարտ: Երբեմն, նույնիսկ առանձին գեների դերը ինչ-որ գործընթացում հաստատելու համար, հերթով իրականացվում է բոլոր գեների սիստեմատիկ «փակումը»։
Դեղագործները նույնպես հետաքրքրվել են siRNA-ի օգտագործման հնարավորությամբ, քանի որ առանձին գեների աշխատանքի ուղղորդված կարգավորման ունակությունը խոստանում է չլսված հեռանկարներ մի շարք հիվանդությունների բուժման համար: Փոքր չափը և գործողության բարձր առանձնահատկությունը խոստանում են siRNA-ի վրա հիմնված դեղամիջոցների բարձր արդյունավետություն և ցածր թունավորություն; սակայն, դեռևս հնարավոր չի եղել լուծել մարմնի հիվանդ բջիջներին siRNA առաքման խնդիրը՝ այս մոլեկուլների փխրունության և փխրունության պատճառով: Ու թեև այժմ տասնյակ թիմեր փորձում են գտնել այս «կախարդական փամփուշտները» ուղիղ թիրախի վրա (հիվանդ օրգանների ներսում), նրանք դեռ տեսանելի հաջողության չեն հասել։ Բացի դրանից, կան նաև այլ բարդություններ. Օրինակ, հակավիրուսային թերապիայի դեպքում siRNA-ի գործողության բարձր ընտրողականությունը կարող է լինել «արատ ծառայություն». քանի որ վիրուսները արագորեն մուտացվում են, փոփոխված շտամը շատ արագ կկորցնի զգայունությունը թերապիայի սկզբում ընտրված siRNA-ի նկատմամբ. Հայտնի է, որ siRNA-ում միայն մեկ նուկլեոտիդի փոխարինումը հանգեցնում է միջամտության ազդեցության զգալի նվազմանը:
Այս պահին հարկ է ևս մեկ անգամ հիշել. siRNA-ները հայտնաբերվել են միայն բույսերի, անողնաշարավորների և միաբջիջ օրգանիզմների մոտ. չնայած ՌՆԹ-ի միջամտության համար սպիտակուցների հոմոլոգները (Dicer, RISC համալիր) նույնպես առկա են բարձրակարգ կենդանիների մոտ, siRNA-ները չեն հայտնաբերվել սովորական մեթոդներով: Ինչպիսի՜ զարմանք էր, երբ արհեստականորեն ներմուծված siRNA-ի սինթետիկ անալոգները կաթնասունների բջիջների կուլտուրաներում առաջացրին դոզանից կախված ուժեղ ազդեցություն: Սա նշանակում էր, որ ողնաշարավորների բջիջներում ՌՆԹ-ի միջամտությունը չի փոխարինվել ավելի բարդ իմունային համակարգերով, այլ զարգացել է օրգանիզմների հետ մեկտեղ՝ դառնալով ավելի «առաջադեմ» մի բան։ Հետևաբար, կաթնասունների մոտ անհրաժեշտ էր փնտրել ոչ թե siRNA-ի ճշգրիտ անալոգներ, այլ դրանց էվոլյուցիոն ժառանգորդներ։
Խաղացող # 2 - միկրոՌՆԹ
Իրոք, ավելի զարգացած օրգանիզմներում ՌՆԹ-ի միջամտության էվոլյուցիոն բավականին հնագույն մեխանիզմի հիման վրա ի հայտ են եկել գեների աշխատանքը վերահսկելու երկու մասնագիտացված համակարգեր, որոնցից յուրաքանչյուրն օգտագործում է փոքր ՌՆԹ-ների իր խումբը՝ միկրոՌՆԹ (microRNA) և piRNA (piRNA, Piwi): - փոխազդող ՌՆԹ): Երկու համակարգերն էլ հայտնվեցին սպունգների և կոելենտերատների մեջ և զարգացան նրանց հետ միասին՝ տեղաշարժելով siRNA-ն և «մերկ» ՌՆԹ-ի միջամտության մեխանիզմը: Նրանց դերը անձեռնմխելիության ապահովման գործում նվազում է, քանի որ այս ֆունկցիան ստանձնել են բջջային իմունիտետի ավելի առաջադեմ մեխանիզմները, մասնավորապես՝ ինտերֆերոնային համակարգը: Այնուամենայնիվ, այս համակարգն այնքան զգայուն է, որ աշխատում է հենց siRNA-ի վրա. կաթնասունների բջիջներում փոքր երկշղթա ՌՆԹ-ների հայտնվելը «տագնապ» է առաջացնում (ակտիվացնում է ինտերֆերոնի սեկրեցումը և առաջացնում է ինտերֆերոնից կախված գեների արտահայտումը, որն արգելափակում է բոլոր թարգմանությունները: գործընթացները ամբողջությամբ): Այս առումով, բարձրակարգ կենդանիների մոտ ՌՆԹ-ի միջամտության մեխանիզմը միջնորդվում է հիմնականում միկրոՌՆԹ-ի և պԻՌՆԹ-ի միջոցով՝ կոնկրետ կառուցվածքով միաշղթա մոլեկուլներ, որոնք չեն հայտնաբերվում ինտերֆերոնային համակարգի կողմից:
Քանի որ գենոմը դառնում է ավելի բարդ, միկրոՌՆԹ-ները և պիրՆԹ-ները ավելի ու ավելի են ներգրավվում տրանսկրիպցիայի և թարգմանության կարգավորման մեջ: Ժամանակի ընթացքում դրանք վերածվեցին գենոմի կարգավորման լրացուցիչ, ճշգրիտ և նուրբ համակարգի: Ի տարբերություն siRNA-ի, միկրոՌՆԹ-ն և պիրՌՆԹ-ն (հայտնաբերվել է 2001 թ., տես նկ. 3, A-B) չեն արտադրվում օտարերկրյա երկշղթա ՌՆԹ մոլեկուլներից, այլ սկզբնապես կոդավորված են ընդունող օրգանիզմի գենոմում:
ՄիՌՆԹ-ի պրեկուրսորը տառադարձվում է գենոմային ԴՆԹ-ի երկու շղթաներից ՌՆԹ պոլիմերազ II-ով, ինչի արդյունքում առաջանում է միջանկյալ ձև՝ pri-miRNA, որը կրում է սովորական mRNA-ի՝ m7G-cap և polyA-tail-ի հատկանիշները: Այս պրեկուրսորը ստեղծում է հանգույց երկու միաշղթա «պոչերով» և մի քանի չզույգված նուկլեոտիդներով կենտրոնում (նկ. 3Ա): Նման օղակը ենթարկվում է երկփուլ մշակման (նկ. Բ). նախ՝ Դրոշա էնդոնուկլեազը կտրում է միաշղթա ՌՆԹ-ի «պոչերը» մազակալից, որից հետո կտրված մազակալը (նախամիկրոՌՆԹ) արտահանվում է ցիտոպլազմա, որտեղ այն ճանաչվում է Dicer-ի կողմից, որը ևս երկու հատում է կատարում (երկթելանի հատվածը կտրված է գունային կոդով Նկար 3Ա-ում): Այս ձևով հասուն միկրոՌՆԹ-ն, ինչպես siRNA-ն, ներառված է RISC համալիրում:
Շատ miRNA-ների գործողության մեխանիզմը նման է siRNA-ին. կարճ (21-25 նուկլեոտիդներ) միաշղթա ՌՆԹ-ն RISC սպիտակուցային համալիրում բարձր յուրահատկությամբ կապվում է թիրախային mRNA-ի 3'-չթարգմանված շրջանի կոմպլեմենտար շրջանին: . Կապի արդյունքում առաջանում է mRNA-ի տրոհումը Ago սպիտակուցի կողմից: Այնուամենայնիվ, միկրոՌՆԹ-ի ակտիվությունը (սիՌՆԹ-ի համեմատ) արդեն ավելի տարբերակված է. եթե կոմպլեմենտարությունը բացարձակ չէ, թիրախային mRNA-ն կարող է չքայքայվել, այլ միայն շրջելիորեն արգելափակվել (թարգմանություն չի լինի): Նույն RISC համալիրը կարող է օգտագործել նաև արհեստականորեն ներմուծված siRNAs: Սա բացատրում է, թե ինչու են siRNA-ները, որոնք պատրաստված են նախակենդանիների անալոգիայով, ակտիվ են նաև կաթնասունների մոտ:
Այսպիսով, մենք կարող ենք լրացնել ավելի բարձր (երկկողմանի սիմետրիկ) օրգանիզմներում ՌՆԹ-ի ինտերֆերենցիայի գործողության մեխանիզմի նկարազարդումը` մեկ պատկերում համատեղելով միկրոՌՆԹ-ների և կենսատեխնոլոգիականորեն ներդրված siRNA-ների գործողության սխեման (նկ. 3B):
Բրինձ. 3A. Երկաշղթա միկրոՌՆԹ պրեկուրսորի մոլեկուլի կառուցվածքը
Հիմնական առանձնահատկությունները. պահպանողական հաջորդականությունների առկայությունը, որոնք կազմում են վարսահարդարիչ; 3'-վերջում երկու «լրացուցիչ» նուկլեոտիդներով լրացուցիչ պատճենի (microRNA *) առկայությունը. հատուկ հաջորդականություն (2–8 bp), որը կազմում է էնդոնուկլեազների ճանաչման վայր։ ՄիկրոՌՆԹ-ն ինքնին ընդգծված է կարմիրով. ահա թե ինչ է կտրում Dicer-ը:

Բրինձ. 3B. միկրոՌՆԹ-ի մշակման և դրա գործունեության իրականացման ընդհանուր մեխանիզմը

Բրինձ. 3B. Արհեստական միկրոՌՆԹ-ների և siRNA-ների գործողության ընդհանրացված սխեման
Արհեստական siRNA-ները ներմուծվում են բջիջ՝ օգտագործելով մասնագիտացված պլազմիդներ (թիրախավորող siRNA վեկտոր):
ՄիկրոՌՆԹ-ի գործառույթները
ՄիկրոՌՆԹ-ների ֆիզիոլոգիական գործառույթները չափազանց բազմազան են. իրականում նրանք հանդես են գալիս որպես օնտոգենեզի հիմնական ոչ սպիտակուցային կարգավորիչներ: miRNA-ները չեն չեղարկվում, այլ լրացնում են գեների կարգավորման «դասական» սխեման (ինդուկտորներ, ճնշողներ, քրոմատինի խտացում և այլն): Բացի այդ, ինքնին միՌՆԹ-ների սինթեզը կարգավորվում է բարդ ձևով (միՌՆԹ-ների որոշ լողավազաններ կարող են միացնել ինտերֆերոնները, ինտերլեյկինները, ուռուցքային նեկրոզային գործոն α (TNF-α) և շատ այլ ցիտոկիններ): Արդյունքում, առաջանում է հազարավոր գեներից բաղկացած «նվագախմբի» թյունինգի զարմանալիորեն բարդ և ճկուն բազմաստիճան ցանց, բայց սա նույնպես չի ավարտվում:
միկրոՌՆԹ-ներն ավելի «ունիվերսալ» են, քան siRNA-ները. պարտադիր չէ, որ «պահեստային» գեները 100%-ով փոխլրացնող լինեն. կարգավորումն իրականացվում է նույնիսկ մասնակի փոխազդեցության դեպքում: Այսօր մոլեկուլային կենսաբանության ամենաթեժ թեմաներից մեկը միկրոՌՆԹ-ների որոնումն է, որոնք հանդես են գալիս որպես հայտնի ֆիզիոլոգիական գործընթացների այլընտրանքային կարգավորիչներ: Օրինակ, miRNAs-ները, որոնք ներգրավված են բույսերի, Drosophila-ի և nematodes-ի բջջային ցիկլի և ապոպտոզի կարգավորման մեջ, արդեն նկարագրված են. մարդկանց մոտ միկրոՌՆԹ-ները կարգավորում են իմունային համակարգը և արյունաստեղծ ցողունային բջիջների զարգացումը: Բիոչիպերի վրա հիմնված տեխնոլոգիաների կիրառումը (միկրո-զանգվածի ցուցադրություն) ցույց է տվել, որ փոքր ՌՆԹ-ների ամբողջ լողավազանները միացվում և անջատվում են բջիջների կյանքի տարբեր փուլերում: Կենսաբանական պրոցեսների համար հայտնաբերվել են տասնյակ հատուկ միկրոՌՆԹ-ներ, որոնց արտահայտման մակարդակը որոշակի պայմաններում փոխվում է հազարավոր անգամ՝ ընդգծելով այդ գործընթացների բացառիկ վերահսկելիությունը։
Մինչև վերջերս ենթադրվում էր, որ միկրոՌՆԹ-ները միայն ամբողջությամբ կամ մասամբ են ճնշում գեների աշխատանքը: Այնուամենայնիվ, վերջերս պարզվեց, որ միկրոՌՆԹ-ների ազդեցությունը կարող է արմատապես տարբեր լինել՝ կախված բջջի վիճակից։ Ակտիվորեն բաժանվող բջիջում միկրոՌՆԹ-ն, կապված լինելով mRNA-ի 3'-տարածաշրջանի կոմպլեմենտար հաջորդականության հետ, արգելակում է սպիտակուցի սինթեզը (թարգմանություն): Այնուամենայնիվ, հանգստի կամ սթրեսային վիճակում (օրինակ, երբ աճում է աղքատ միջավայրում), նույն իրադարձությունը հանգեցնում է ճիշտ հակառակ ազդեցության՝ թիրախային սպիտակուցի սինթեզի ավելացմանը:
ՄիկրոՌՆԹ-ի էվոլյուցիան
Բարձրագույն օրգանիզմներում միկրոՌՆԹ-ի սորտերի թիվը դեռ լիովին հաստատված չէ. որոշ տվյալների համաձայն՝ այն գերազանցում է սպիտակուցը կոդավորող գեների թվի 1%-ը (մարդկանց մոտ, օրինակ, ասում են մոտ 700 միկրոՌՆԹ, և այդ թիվը մշտապես առկա է. աճող): ՄիկրոՌՆԹ-ները կարգավորում են բոլոր գեների մոտ 30%-ի գործունեությունը (դրանցից շատերի թիրախները դեռ հայտնի չեն), և կան և՛ ամենուր տարածված, և՛ հյուսվածքներին հատուկ մոլեկուլներ, օրինակ՝ միկրոՌՆԹ-ների նման կարևոր լողավազան կարգավորում է արյան ցողունային բջիջների հասունացումը։ .
Տարբեր օրգանիզմների տարբեր հյուսվածքներում արտահայտման լայն պրոֆիլը և միկրոՌՆԹ-ների կենսաբանական տարածվածությունը վկայում են էվոլյուցիոն հնագույն ծագման մասին: Առաջին անգամ miRNA-ները հայտնաբերվեցին նեմատոդների մեջ, և երկար ժամանակ անց ենթադրվում էր, որ այդ մոլեկուլները հայտնվում են միայն սպունգներում և կոելենտերատներում. սակայն հետագայում դրանք հայտնաբերվել են նաև միաբջիջ ջրիմուռներում: Հետաքրքիր է, որ քանի որ օրգանիզմները դառնում են ավելի բարդ, միկրոՌՆԹ-ի ավազանի քանակն ու տարասեռությունը նույնպես մեծանում է: Սա անուղղակիորեն ցույց է տալիս, որ այդ օրգանիզմների բարդությունն ապահովվում է, մասնավորապես, միկրոՌՆԹ-ի գործառմամբ: ՄիկրոՌՆԹ-ների հնարավոր էվոլյուցիան ներկայացված է Նկար 4-ում:

Բրինձ. 4. Տարբեր օրգանիզմների միկրոՌՆԹ-ների բազմազանությունը
Որքան բարձր է օրգանիզմի կազմակերպվածությունը, այնքան ավելի շատ միկրոՌՆԹ է հայտնաբերվում նրա մեջ (թիվը փակագծերում)։ Միայնակ միկրոՌՆԹ-ներով տեսակները ընդգծված են կարմիրով: Համաձայն .
Հստակ էվոլյուցիոն հարաբերություններ կարելի է կազմել siRNA-ի և microRNA-ի միջև՝ հիմնվելով հետևյալ փաստերի վրա.
- Երկու տեսակների գործողությունը փոխարինելի է և միջնորդվում է հոմոլոգ սպիտակուցներով.
- Կաթնասունների բջիջներում ներմուծված siRNA-ները հատուկ «անջատում են» ցանկալի գեները (չնայած ինտերֆերոնի պաշտպանության որոշակի ակտիվացմանը);
- միկրոՌՆԹ-ները հայտնաբերվում են ավելի ու ավելի շատ հին օրգանիզմներում:
Այս և այլ տվյալներ հուշում են երկու համակարգերի ծագումն ընդհանուր «նախահայրից»: Հետաքրքիր է նաև նշել, որ «ՌՆԹ» անձեռնմխելիությունը, որպես սպիտակուցային հակամարմինների անկախ նախադրյալ, հաստատում է կյանքի առաջին ձևերի ծագման տեսությունը ՌՆԹ-ի, այլ ոչ թե սպիտակուցների հիման վրա (հիշենք, որ սա ակադեմիկոսի սիրելի տեսությունն է։ AS Spirin):
Թեև մոլեկուլային կենսաբանության ասպարեզում կային ընդամենը երկու «խաղացողներ»՝ siRNA և microRNA, ՌՆԹ-ի միջամտության հիմնական «նպատակը» թվում էր լիովին պարզ: Իրոք. տարբեր օրգանիզմների հոմոլոգ կարճ ՌՆԹ-ների և սպիտակուցների մի շարք կատարում են նմանատիպ գործողություններ. քանի որ օրգանիզմները դառնում են ավելի բարդ, ֆունկցիոնալությունը դառնում է ավելի բարդ:
Այնուամենայնիվ, էվոլյուցիայի գործընթացում բնությունը ստեղծել է մեկ այլ, էվոլյուցիոն առումով ամենավերջին և բարձր մասնագիտացված համակարգ, որը հիմնված է ՌՆԹ-ի միջամտության նույն հաջող սկզբունքի վրա: Խոսքը պԻՌՆԹ-ի մասին է (piRNA, Piwi-interaction RNA-ից):
Որքան ավելի բարդ է գենոմը կազմակերպված, այնքան ավելի զարգացած և հարմարեցված է օրգանիզմը (կամ հակառակը ;-): Այնուամենայնիվ, գենոմի աճող բարդությունն ունի նաև բացասական կողմ՝ բարդ գենետիկ համակարգը դառնում է անկայուն: Սա հանգեցնում է գենոմի ամբողջականության պահպանման համար պատասխանատու մեխանիզմների անհրաժեշտությանը, հակառակ դեպքում ԴՆԹ-ի ինքնաբուխ «խառնումը» պարզապես կանջատի այն: Շարժական գենետիկական տարրերը (MGE) - գենոմի անկայունության հիմնական գործոններից մեկը - կարճ անկայուն շրջաններ են, որոնք կարող են ինքնուրույն տառադարձվել և գաղթել գենոմով մեկ: Նման շարժական տարրերի ակտիվացումը հանգեցնում է քրոմոսոմներում ԴՆԹ-ի բազմաթիվ ընդմիջումների՝ հղի մահացու հետեւանքներով։
SHE-ների թիվը գենոմի չափի հետ ոչ գծային է ավելանում, և դրանց ակտիվությունը պետք է պարունակվի: Դրա համար կենդանիները, սկսած կոելենտերատներից, օգտագործում են ՌՆԹ միջամտության նույն երեւույթը։ Այս ֆունկցիան կատարում են նաև կարճ ՌՆԹ-ները, բայց ոչ նրանք, որոնք արդեն քննարկվել են, այլ դրանց երրորդ տիպը՝ պիրՆԹ-ները։
piRNA-ի «դիմանկար».
piRNA-ները 24-30 նուկլեոտիդների երկարությամբ կարճ մոլեկուլներ են, որոնք կոդավորված են քրոմոսոմի ցենտրոմերային և տելոմերային հատվածներում։ Դրանցից շատերի հաջորդականությունները լրացնում են հայտնի շարժական գենետիկական տարրերին, սակայն կան շատ այլ պԻՌՆԹ-ներ, որոնք համընկնում են գործող գեների շրջանների կամ գենոմի բեկորների հետ, որոնց գործառույթներն անհայտ են:
piRNA-ները (ինչպես նաև միկրոՌՆԹ-ները) կոդավորված են գենոմային ԴՆԹ-ի երկու շղթաներում. դրանք խիստ փոփոխական են և բազմազան (մինչև 500000 (!) տեսակ մեկ օրգանիզմում): Ի տարբերություն siRNA-ի և microRNA-ի, դրանք ձևավորվում են մեկ շղթայով, որն ունի բնորոշ հատկանիշ՝ ուրացիլ (U) 5' ծայրում և մեթիլացված 3' ծայրում: Կան նաև այլ տարբերություններ.
- Ի տարբերություն siRNA-ի և microRNA-ի, նրանք չեն պահանջում Dicer-ի մշակում;
- ՊԻՌՆԹ գեները ակտիվ են միայն սեռական բջիջներում (սաղմնածինության ընթացքում) և շրջակա էնդոթելիային բջիջներում.
- ՊիՌՆԹ համակարգի սպիտակուցային կազմը տարբեր է. դրանք Piwi դասի էնդոնուկլեազներ են (Piwi և Aub) և առանձին Argonaute բազմազանություն՝ Ago3:
ՊիՌՆԹ-ների մշակումն ու ակտիվությունը դեռևս վատ է հասկացված, բայց արդեն պարզ է, որ գործողության մեխանիզմը բոլորովին տարբերվում է մյուս կարճ ՌՆԹ-ներից. այսօր առաջարկվել է նրանց աշխատանքի պինգ-պոնգ մոդելը (նկ. 5 Ա, Բ):
ՊիՌՆԹ-ի կենսագենեզի պինգ-պոնգի մեխանիզմը

Բրինձ. 5A. piRNA-ի մշակման ցիտոպլազմային մաս
ՊիՌՆԹ-ի բիոգենեզը և ակտիվությունը միջնորդվում է Piwi էնդոնուկլեազների ընտանիքի կողմից (Ago3, Aub, Piwi): ՊիՌՆԹ-ի ակտիվությունն ապահովվում է երկու միաշղթա պիրՆԹ-ի մոլեկուլներով՝ զգայական և հակազգայական, որոնցից յուրաքանչյուրը կապված է հատուկ Piwi էնդոնուկլեազի հետ: ՊիՌՆԹ-ն ճանաչում է տրանսպոզոնային mRNA-ի (կապույտ շղթա) կոմպլեմենտար շրջանը և կտրում այն: Սա ոչ միայն ապաակտիվացնում է տրանսպոզոնը, այլև ստեղծում է նոր piRNA (կապված է Ago3-ի հետ 3'-վերջի մեթիլացման միջոցով Hen1 մեթիլազով): Այս պԻՌՆԹ-ն, իր հերթին, ճանաչում է mRNA-ն piRNA-ի պրեկուրսորների կլաստերի (կարմիր շղթա) տառադարձումներով - այս կերպ ցիկլը փակվում է և նորից արտադրվում է ցանկալի պիրՆԹ-ն:

Բրինձ. 5B: piRNA միջուկում
Բացի էնդոնուկլեազ Aub-ից, Piwi էնդոնուկլեազը կարող է նաև կապել հակազգայական piRNA-ն: Միանալուց հետո կոմպլեքսը տեղափոխվում է միջուկ, որտեղ այն առաջացնում է կոմպլեմենտար տառադարձումների քայքայումը և քրոմատինի վերադասավորումը, որն առաջացնում է տրանսպոզոնների ակտիվության ճնշում։
PiRNA գործառույթները
ՊԻՌՆԹ-ի հիմնական ֆունկցիան է ճնշել MGE-ի ակտիվությունը տրանսկրիպցիայի և թարգմանության մակարդակում: Ենթադրվում է, որ պԻՌՆԹ-ները ակտիվ են միայն սաղմի առաջացման ժամանակ, երբ գենոմի անկանխատեսելի խառնումը հատկապես վտանգավոր է և կարող է հանգեցնել սաղմի մահվան: Սա տրամաբանական է. երբ իմունային համակարգը դեռ չի սկսել աշխատել, սաղմի բջիջները պարզ, բայց արդյունավետ պաշտպանության կարիք ունեն։ Սաղմը հուսալիորեն պաշտպանված է արտաքին պաթոգեններից պլասենցայի (կամ ձվի կեղևի) միջոցով: Բայց բացի դրանից, անհրաժեշտ է նաև պաշտպանություն էնդոգեն (ներքին) վիրուսներից, առաջին հերթին MGE-ից:
PiRNA-ի այս դերը հաստատվել է փորձով. «նոկաուտը» կամ Ago3, Piwi կամ Aub գեների մուտացիաները հանգեցնում են զարգացման լուրջ խանգարումների (և նման օրգանիզմի գենոմում մուտացիաների քանակի կտրուկ աճին), ինչպես նաև առաջացնում են: անպտղություն սեռական բջիջների զարգացման խանգարման պատճառով:
ՊիՌՆԹ-ի բաշխումը և էվոլյուցիան
Առաջին piRNA-ները հայտնաբերվել են արդեն անեմոնների և սպունգերի մեջ: Բույսերը, ըստ երևույթին, գնացին այլ ուղի. դրանցում Piwi սպիտակուցներ չեն հայտնաբերվել, իսկ էնդոնուկլեազը Ago4-ը և siRNA-ն տրանսպոզոնների համար դունչի դեր են խաղում:
Բարձրագույն կենդանիների, ներառյալ մարդկանց մոտ, piRNA համակարգը շատ լավ զարգացած է, բայց այն կարող է հայտնաբերվել միայն սաղմնային բջիջներում և ամնիոտիկ էնդոթելիում: Թե ինչու է piRNA-ների բաշխումը մարմնում այդքան սահմանափակ, մնում է պարզել: Կարելի է ենթադրել, որ, ինչպես ցանկացած հզոր զենք, piRNA-ները օգտակար են միայն շատ կոնկրետ պայմաններում (պտղի զարգացման ընթացքում), իսկ մեծահասակների օրգանիզմում նրանց գործունեությունը ավելի շատ վնաս կտա, քան օգուտ: Այնուամենայնիվ, piRNA-ների թիվը մեծության կարգով ավելի մեծ է, քան հայտնի սպիտակուցների թիվը, և դժվար է կանխատեսել պԻՌՆԹ-ների ոչ սպեցիֆիկ ազդեցությունները հասուն բջիջներում:
| siRNA | միկրոՌՆԹ | piRNA | |
| Տարածում | Բույսեր, Դրոզոֆիլա, C. էլեգանս... Չի հայտնաբերվել ողնաշարավորների մոտ | Էուկարիոտներ | Սաղմնային կենդանիների բջիջներ (սկսած կոելենտերատներից): Ոչ նախակենդանիների և բույսերի մեջ |
| Երկարություն | 21-22 նուկլեոտիդներ | 19-25 նուկլեոտիդներ | 24-30 նուկլեոտիդներ |
| Կառուցվածք | Կրկնաշղթա, 19 փոխլրացնող նուկլեոտիդ և երկու չզույգված նուկլեոտիդ 3' ծայրում | Միաշղթա բարդ կառուցվածք | Միաշղթա բարդ կառուցվածք. U 5'-վերջում, 2'- Օ-մեթիլացված 3'-վերջ |
| Մշակում | Dicer-կախված | Dicer-կախված | Dicer-անկախ |
| Էնդոնուկլեազ | Ago2 | Ago1, Ago2 | Ago3, Piwi, Aub |
| Գործունեություն | Կոմպլեմենտար mRNA-ի դեգրադացիա, գենոմային ԴՆԹ-ի ացետիլացում | Թիրախային mRNA-ի թարգմանության քայքայումը կամ արգելակումը | MGE կոդավորող mRNA-ի դեգրադացիա, MGE տրանսկրիպցիայի կարգավորում |
| Կենսաբանական դեր | Հակավիրուսային իմունային պաշտպանություն, սեփական գեների գործունեության ճնշում | Գենի գործունեության կարգավորումը | Էմբրիոգենեզի ընթացքում MGE-ի գործունեության ճնշումը |
Եզրակացություն
Եզրափակելով, ես կցանկանայի տալ աղյուսակ, որը ցույց է տալիս ՌՆԹ-ի միջամտության մեջ ներգրավված սպիտակուցային ապարատի էվոլյուցիան (նկ. 6): Կարելի է տեսնել, որ նախակենդանիներն ունեն ամենազարգացած siRNA համակարգը (սպիտակուցային ընտանիքներ Ago, Dicer), և քանի որ օրգանիզմները դառնում են ավելի բարդ, շեշտը տեղափոխվում է ավելի մասնագիտացված համակարգերի վրա՝ միկրոՌՆԹ-ի (Դրոշա, Փաշա) և պիրՆԹ-ի սպիտակուցային իզոֆորմների քանակը: (Piwi, Hen1) ավելանում է: Միևնույն ժամանակ, siRNA-ի գործողությանը միջնորդող ֆերմենտների բազմազանությունը նվազում է։

Բրինձ. 6. ՌՆԹ-ի միջամտության մեջ ներգրավված սպիտակուցների բազմազանությունը
և
Թվերը ցույց են տալիս յուրաքանչյուր խմբի սպիտակուցների քանակը: ՍԻՌՆԹ-ին և միկրոՌՆԹ-ին բնորոշ տարրերը ընդգծված են կապույտով, իսկ պիրՆԹ-ի հետ կապված սպիտակուցները՝ կարմիրով: Համաձայն .
ՌՆԹ-ի միջամտության ֆենոմենն արդեն սկսել է կիրառվել ամենապարզ օրգանիզմների կողմից։ Այս մեխանիզմի հիման վրա բնությունը ստեղծել է իմունային համակարգի նախատիպը, և քանի որ օրգանիզմները դառնում են ավելի բարդ, ՌՆԹ-ի միջամտությունը դառնում է գենոմի ակտիվության անփոխարինելի կարգավորիչը: Երկու տարբեր մեխանիզմներ գումարած երեք տեսակի կարճ ՌՆԹ-ներ (տես ամփոփ աղյուսակը) - արդյունքում մենք տեսնում ենք տարբեր նյութափոխանակության և գենետիկ ուղիների հազարավոր նուրբ կարգավորիչներ: Այս ապշեցուցիչ նկարը ցույց է տալիս մոլեկուլյարների բազմակողմանիությունն ու էվոլյուցիոն հարմարվողականությունը կենսաբանական համակարգեր... Կարճ ՌՆԹ-ները կրկին ապացուցում են, որ բջջի ներսում «փոքր բաներ» չկան. կան միայն փոքր մոլեկուլներ, որոնց դերի ողջ նշանակությունը մենք նոր ենք սկսում հասկանալ:
Ճիշտ է, նման ֆանտաստիկ բարդությունը ավելի շուտ խոսում է այն մասին, որ էվոլյուցիան «կույր» է և գործում է առանց նախապես հաստատված «գլխավոր պլանի»։
գրականություն
- Gurdon J.B., Lane C.D., Woodland H.R., Marbaix G. (1971): Գորտի ձվերի և ձվաբջիջների օգտագործումը սուրհանդակային ՌՆԹ-ի և կենդանի բջիջներում դրա թարգմանության ուսումնասիրության համար: Բնություն 233,177-182;
- Spirin A.S. (2001): Սպիտակուցների կենսասինթեզը, ՌՆԹ-ի աշխարհը և կյանքի ծագումը. ՌԱՆ Տեղեկագիր 71, 320-328;
- Տարրեր. «Վերած կենդանիների ամբողջական միտոքոնդրիալ գենոմն այժմ կարելի է հանել մազերից»;
- Fire A., Xu S., Montgomery M.K., Kostas S.A., Driver S.E., Mello C.C. (1998): Ուժեղ և հատուկ գենետիկական միջամտություն երկշղթա ՌՆԹ-ով Caenorhabditis elegans... Nature 391, 806-311;
- Կենսամոլեկուլ. «ՄիկրոՌՆԹ-ները առաջին անգամ հայտնաբերվել են միաբջիջ օրգանիզմում»;
- Covey S., Al-Kaff N., Lángara A., Turner D. (1997): Բույսերը վարակի դեմ պայքարում են գեների լռեցման միջոցով: Nature 385, 781-782;
- Կենսամոլեկուլ. «Մոլեկուլային կրկնակի գործարկում. մարդկային գեները գործում են գրիպի վիրուսի համար»;
- Ren B. (2010). Տառադարձում. ուժեղացուցիչները ստեղծում են ոչ կոդավորող ՌՆԹ: Բնություն 465,173-174;
- Taganov K.D., Boldin M.P., Chang K.J., Baltimore D. (2006): NF-κB-ից կախված միկրոՌՆԹ miR-146 ինդուկցիա, արգելակիչ, որն ուղղված է բնածին իմունային պատասխանների ազդանշանային սպիտակուցներին: Պրոց. Նատլ. ակադ. Գիտ. ԱՄՆ. 103,12481-12486;
- O'Connell R. M., Rao D. S., Chaudhuri A. A., Boldin M. P., Taganov K.D., Nicoll J., Paquette R. L., Baltimore D. (2008): ՄիկրոՌՆԹ-155-ի կայուն արտահայտությունը արյունաստեղծ ցողունային բջիջներում առաջացնում է միելոպրոլիֆերատիվ խանգարում: J. Exp. Բժշկ. 205, 585-594;
- Կենսամոլեկուլ. «microRNA - որքան հեռու է անտառ, այնքան ավելի շատ վառելափայտ»;
- Տարրեր՝ «Հին կենդանիների օրգանիզմի բարդացումը կապված էր նոր կարգավորիչ մոլեկուլների առաջացման հետ»;
- Գրիմսոն Ա., Սրիվաստավա Մ., Ֆահեյ Բ., Վուդկրոֆթ Բ.Ջ., Չիանգ Հ.Ռ., Քինգ Ն., Դեգնան Բ.Մ., Ռոխսար Դ.Ս., Բարթել Դ.Պ. (2008): Կենդանիների մեջ միկրոՌՆԹ-ների և Piwi-ի հետ փոխազդող ՌՆԹ-ների վաղ ծագումն ու էվոլյուցիան: Բնություն 455, 1193-1197 թթ.
- Aravin A., Hannon G, Brennecke J. (2007): Piwi-piRNA ուղին ապահովում է հարմարվողական պաշտպանություն տրանսպոզոնային սպառազինությունների մրցավազքում: Գիտություն 318,761-764;
- Կենսամոլեկուլ.
Գիտնականները կարծում են, որ փոքր ՌՆԹ-ների ոչ ճիշտ արտահայտումը մի շարք հիվանդությունների պատճառներից մեկն է, որոնք շատ լրջորեն ազդում են աշխարհի բազմաթիվ մարդկանց առողջության վրա։ Այդպիսի հիվանդություններից են սրտանոթային 23 և ուռուցքաբանական 24 հիվանդությունները։ Ինչ վերաբերում է վերջինիս, ապա դա զարմանալի չէ. քաղցկեղը վկայում է բջիջների զարգացման և դրանց ճակատագրի շեղումների մասին, իսկ փոքր ՌՆԹ-ները կարևոր դեր են խաղում համապատասխան գործընթացներում։ Ահա քաղցկեղի մարմնի վրա փոքր ՌՆԹ-ների հսկայական ազդեցության ամենաբացահայտ օրինակներից մեկը: Խոսքը չարորակ ուռուցքի մասին է, որը բնութագրվում է այն գեների սխալ արտահայտմամբ, որոնք գործում են ոչ թե հետծննդյան, այլ օրգանիզմի սկզբնական զարգացման ժամանակ։ Դա մանկական ուղեղի ուռուցքի տեսակ է, որը սովորաբար ի հայտ է գալիս մինչև երկու տարեկանը։ Ավաղ, սա քաղցկեղի շատ ագրեսիվ ձև է, և կանխատեսումը վատ է նույնիսկ ինտենսիվ բուժման դեպքում: Ուռուցքաբանական պրոցեսը զարգանում է ուղեղի բջիջներում գենետիկական նյութի ոչ պատշաճ վերաբաշխման արդյունքում։ Պրոմոտորը, որը սովորաբար առաջացնում է սպիտակուցներ կոդավորող գեներից մեկի ուժեղ արտահայտում, ենթարկվում է ռեկոմբինացիայի փոքր ՌՆԹ-ների հատուկ կլաստերի հետ: Այնուհետև այս ամբողջ վերադասավորվող շրջանն ընդլայնվում է. այլ կերպ ասած, դրա բազմաթիվ պատճեններ են ստեղծվում գենոմում: Հետևաբար, փոքր ՌՆԹ-ները, որոնք գտնվում են «ներքևում», քան տեղահանված պրոմոուտերը, արտահայտվում են շատ ավելի ուժեղ, քան պետք է: Ակտիվ փոքր ՌՆԹ-ի պարունակության մակարդակը նորմայից մոտավորապես 150-1000 անգամ բարձր է։
Բրինձ. 18.3.Ալկոհոլի կողմից ակտիվացված փոքր ՌՆԹ-ները կարող են կապվել սուրհանդակային ՌՆԹ-ների հետ, որոնք չեն ազդում ալկոհոլի նկատմամբ մարմնի դիմադրության վրա: Բայց այս փոքր ՌՆԹ-ները չեն կապվում ՌՆԹ-ի սուրհանդակային մոլեկուլների հետ, որոնք նպաստում են այս դիմադրությանը: Սա հանգեցնում է սուրհանդակային ՌՆԹ-ի մոլեկուլների հարաբերական գերակշռությանը, որոնք կոդավորում են սպիտակուցային տատանումները՝ կապված ալկոհոլի դիմադրության հետ:
Այս կլաստերը կոդավորում է ավելի քան 40 տարբեր փոքր ՌՆԹ: Իրականում, սա ընդհանուր առմամբ ամենամեծն է պրիմատների մոտ առկա նման կլաստերներից: Այն սովորաբար արտահայտվում է միայն մարդու զարգացման վաղ փուլում՝ սաղմի կյանքի առաջին 8 շաբաթվա ընթացքում։ Նրա ուժեղ ակտիվացումը նորածնի ուղեղում հանգեցնում է աղետալի ազդեցության գենետիկական արտահայտման վրա: Հետևանքներից մեկը էպիգենետիկ սպիտակուցի արտահայտումն է, որը փոփոխություններ է ավելացնում ԴՆԹ-ին: Սա հանգեցնում է ԴՆԹ-ի մեթիլացման ողջ օրինաչափության լայնածավալ փոփոխությունների, հետևաբար՝ բոլոր տեսակի գեների աննորմալ արտահայտման, որոնցից շատերը պետք է արտահայտվեն միայն այն ժամանակ, երբ ուղեղի անհաս բջիջները բաժանվում են մարմնի զարգացման վաղ փուլերում: Ահա թե ինչպես է սկսվում քաղցկեղի 25 ծրագիրը մանկական բջիջներում։
Փոքր ՌՆԹ-ների և բջջի էպիգենետիկ ապարատի նման հաղորդակցությունը կարող է զգալի ազդեցություն ունենալ այլ իրավիճակների վրա, երբ բջիջներում առաջանում է քաղցկեղի հակում: Այս մեխանիզմը, հավանաբար, հանգեցնում է նրան, որ փոքր ՌՆԹ-ների արտահայտման խանգարման ազդեցությունն ուժեղանում է էպիգենետիկ մոդիֆիկացիաների փոփոխությամբ, որոնք փոխանցվում են մորից դուստր բջիջներին: Այսպես կարող է ձևավորվել գեների արտահայտման բնույթի պոտենցիալ վտանգավոր փոփոխությունների սխեման։
Առայժմ գիտնականները չեն պարզել փոքր ՌՆԹ-ների էպիգենետիկ գործընթացների հետ փոխազդեցության բոլոր փուլերը, բայց նրանց դեռ հաջողվում է որոշ ակնարկներ ստանալ տեղի ունեցողի առանձնահատկությունների մասին: Օրինակ, պարզվեց, որ փոքր ՌՆԹ-ների որոշակի դաս, որոնք մեծացնում են կրծքագեղձի քաղցկեղի ագրեսիվությունը, թիրախավորում են սուրհանդակային ՌՆԹ-ի որոշ ֆերմենտներ, որոնք հեռացնում են հիմնական էպիգենետիկ փոփոխությունները: Սա փոխում է քաղցկեղի բջջի էպիգենետիկ փոփոխությունների օրինաչափությունը և հետագայում խաթարում է գենետիկական արտահայտությունը 26:
Քաղցկեղի շատ տեսակներ դժվար է հետևել հիվանդի մոտ: Ուռուցքաբանական պրոցեսները կարող են տեղի ունենալ դժվարամատչելի վայրերում, ինչը բարդացնում է նմուշառման ընթացակարգը։ Նման դեպքերում բժշկի համար դժվար է վերահսկել քաղցկեղի գործընթացի զարգացումը և բուժման արձագանքը: Հաճախ բժիշկները ստիպված են լինում ապավինել անուղղակի չափումների՝ ասենք, ուռուցքի տոմոգրաֆիկ սկանավորմանը: Որոշ հետազոտողներ կարծում են, որ փոքր ՌՆԹ մոլեկուլները կարող են օգնել ստեղծել ուռուցքի զարգացման մոնիտորինգի նոր մեթոդ, որը նաև թույլ է տալիս ուսումնասիրել դրա ծագումը: Երբ քաղցկեղի բջիջները մահանում են, փոքր ՌՆԹ-ները հեռանում են բջիջներից, երբ դրանք պատռվում են: Այս փոքրիկ անպետք մոլեկուլները հաճախ բարդույթներ են կազմում բջջային սպիտակուցներկամ փաթաթված բեկորներով բջջային մեմբրաններ... Սա նրանց շատ կայուն է դարձնում հեղուկ կրիչներօրգանիզմ, ինչը նշանակում է, որ նման ՌՆԹ-ն կարելի է մեկուսացնել և վերլուծել։ Քանի որ դրանց թիվը փոքր է, հետազոտողները ստիպված կլինեն օգտագործել խիստ զգայուն վերլուծական մեթոդներ: Այնուամենայնիվ, այստեղ անհնարին ոչինչ չկա. հաջորդականության զգայունությունը նուկլեինաթթուներանընդհատ աճող 27. Հրապարակված տվյալներ, որոնք հաստատում են այս մոտեցման խոստումը կրծքագեղձի քաղցկեղի 28, ձվարանների քաղցկեղի 29 և մի շարք այլ քաղցկեղների հետ կապված։ Թոքերի քաղցկեղով հիվանդների մոտ փոքր շրջանառվող ՌՆԹ-ների վերլուծությունը ցույց է տվել, որ այդ ՌՆԹ-ներն օգնում են տարբերակել մեկ թոքային հանգույցով (թերապիա չպահանջող) և չարորակ ուռուցքային հանգույցներ (բուժում պահանջող) հիվանդների միջև 30:
Ա.Մ. Դեյխման, Ս.Վ.Զինովև, Ա.Յու.Բարիշնիկով
ԳԵՆԻ ԱՐՏԱՀԱՅՏՈՒՄԸ ԵՎ ՓՈՔՐ ՌՆԹ-ն Ուռուցքաբանության մեջ
GU RONTs նրանց. N.N.Blokhina, Ռուսաստանի բժշկական գիտությունների ակադեմիա, Մոսկվա
ԱՄՓՈՓՈՒՄ
Հոդվածում ներկայացված է փոքր ՌՆԹ-ների դերը, որոնք վերահսկում են բջջի և մարմնի կենսական գործառույթների մեծ մասը, և դրանց հնարավոր կապը, մասնավորապես, օնկոգենեզի և գենոմային արտահայտման այլ (ներառյալ հիպոթետիկ) ներբջջային մեխանիզմների հետ:
Հիմնաբառերփոքր ՌՆԹ-ներ, ՌՆԹ-ի միջամտություն (RNAi), երկշղթա ՌՆԹ (lncRNA), ՌՆԹ-ի խմբագրում, օնկոգենեզ:
Ա.Մ. Դեյխման, S. V. Zinoviev, A. Yu. Baryshnikov.
ԳԵՆԱՅԻՆ ԱՐՏԱԴՐՈՒԹՅՈՒՆԸ ԵՎ ՓՈՔՐ ՌՆԹ-ները Ուռուցքաբանության մեջ
Ն.Ն. Բլոխինի անվան քաղցկեղի հետազոտական ռուսական կենտրոն RAMS, մշօվ
Վերացական
Բջիջների և օրգանիզմի կենսագործունեության մեծամասնությունը վերահսկող փոքր ՌՆԹ-ների թղթային դերը և դրանց հնարավոր կապը, մասնավորապես, օնկոգենեզի և գենոմի արտահայտման այլ (ներառյալ հիպոթետիկ) ներբջջային մեխանիզմների հետ:
Բանալի բառերՓոքր ՌՆԹ, միջամտության ՌՆԹ (RNAi), երկշղթա ՌՆԹ (dsRNAs), ՌՆԹ-ի խմբագրում, ուռուցքածին:
Ներածություն
Առանձին գեների և էուկարիոտների ամբողջ գենոմների արտահայտությունը, ներառյալ մշակումը, տարբեր տեսակի տրանսկրիպցիան, զուգավորումը, վերադասավորումը, ՌՆԹ-ի խմբագրումը, վերամիավորումները, թարգմանությունը, ՌՆԹ-ի միջամտությունը, կարգավորվում են որոշ սպիտակուցներով (կարգավորող, կառուցվածքային, հոմեոտիկ գեների արտադրանք, տրանսկրիպցիոն գործոններ: ), շարժական տարրեր, ՌՆԹ և ցածր մոլեկուլային քաշի էֆեկտորներ։ Մշակող ՌՆԹ-ները ներառում են rRNA, tRNA, mRNA, որոշ կարգավորող ՌՆԹ և փոքր ՌՆԹ:
Մինչ օրս հայտնի է, որ փոքր ՌՆԹ-ները չեն կոդավորում սպիտակուցը, հաճախ դրանց թիվը հասնում է հարյուրների մեկ գենոմում և մասնակցում է տարբեր էուկարիոտիկ գեների (սոմատիկ, իմունային, սերմինալ, ցողունային բջիջների) արտահայտման կարգավորմանը: Դիֆերենցման գործընթացները վերահսկվում են (հեմատոպոեզ, անգիոգենեզ, ադիպոգենեզ, միոգենեզ, նեյրոգենեզ), մորֆոգենեզ (ներառյալ սաղմնային փուլերը, զարգացումը/աճը, ֆիզիոլոգիական կարգավորումը), տարածումը, ապոպտոզը, քաղցկեղածինությունը, մուտագենեզը, իմունոգենեզը, ծերացումը (կյանքի երկարացում): ; Եղել են նյութափոխանակության կարգավորման դեպքեր (օրինակ՝ գլիկոսֆինգոլիպիդներ)։ 20-300 / 500 նուկլեոտիդների և դրանց RNP-ների ոչ կոդավորող ՌՆԹ-ների ավելի լայն դասը հայտնաբերվել է ոչ միայն միջուկում / միջուկում / ցիտոպլազմայում, այլև ԴՆԹ պարունակող բջջային օրգանելներում (կենդանիների միտոքոնդրիաներ, միկրո-ՌՆԹ-ներ և քլորոպլաստների տրանսկրիպտների կոնսենսուսային հաջորդականություններ: հայտնաբերված բույսերում): ՌՆԹ):
Վ.Ն.-ի կառավարման և կարգավորման համար գործընթացները, կարևոր է. 1. որ փոքր չափի բնական/արհեստական ՌՆԹ-ն (փոքր ՌՆԹ, tRNA և այլն) և դրանց կոմպլեքսները սպիտակուցների հետ (RNP) կարող են տրանսմեմբրանային բջջային և միտոքոնդրիալ փոխադրում իրականացնել. 2. որ միտոքոնդրիաների քայքայվելուց հետո դրանց պարունակության մի մասը՝ ՌՆԹ-ն և ՌՆՊ-ն, կարող են հայտնվել ցիտոպլազմում և միջուկում։ Փոքր ՌՆԹ-ների (RNPs) թվարկված հատկությունները, որոնց ֆունկցիոնալ նշանակալի դերը միայն աճում է ուսումնասիրության գործընթացում, ակնհայտորեն կապված են քաղցկեղի և այլ գենետիկական հիվանդությունների նկատմամբ զգոնության գործոնի հետ: Միաժամանակ պարզ դարձավ էպիգենոմիկ քրոմատինի փոփոխությունների մեծ նշանակությունը ուռուցքների առաջացման գործում: Նմանատիպ բազմաթիվ դեպքերից մենք կքննարկենք միայն շատ սահմանափակ թվով դեպքեր:
Փոքր ՌՆԹ-ներ
Փոքր ՌՆԹ-ների գործողության մեխանիզմը կայանում է նրանում, որ նրանք կարող են գրեթե լրացնել mRNA թիրախների 3 «չթարգմանված շրջանները (3» UTR) (որոնք երբեմն պարունակում են ԴՆԹ/ՌՆԹ տրանսպոզիտոր MIR/LINE-2 տարրեր, ինչպես նաև պահպանողական Alu կրկնողություններ): ) և առաջացնել ՌՆԹ-ի միջամտություն (RNAi = RNAi; մասնավորապես, հակավիրուսային պատասխանում): Այնուամենայնիվ, բարդությունն այն է, որ բջջայինից բացի, կան վիրուսով կոդավորված փոքր ՌՆԹ-ներ (հերպես, SV40 և այլն; EBV, օրինակ, պարունակում է 23, իսկ KSHV-ն՝ 12 miRNA), որոնք փոխազդում են ինչպես վիրուսի, այնպես էլ տառադարձումների հետ: հյուրընկալող. Միայն ավելի քան 5 հազար բջջային/վիրուսային miRNAs հայտնի են 58 տեսակների մեջ: RNAi-ն սկսում է կամ դեգրադացիան (RISC-ի մասնակցությամբ, ՌՆԹ-ով առաջացած լռեցման համալիր) շարունակական lncRNA շղթաների բեկորների երկայնքով, որոնք խոցելի են նուկլեազների համար (կրկնաշղթա RNA mRNA և այլն), կամ մասամբ շրջելի արգելակում է ընդհատվող ոլորված lncRNA-ն mRNA-ի թարգմանության ընթացքում: թիրախներ. Հասուն փոքր ՌՆԹ-ները (~ 15-28 նուկլեոտիդներ) ձևավորվում են ցիտոպլազմայում տարբեր երկարությունների (տասնյակ և հարյուրավոր նուկլեոտիդներ) դրանց պրեկուրսորներից, որոնք մշակվում են միջուկում։ Բացի այդ, փոքր ՌՆԹ-ները մասնակցում են քրոմատինի խլացնող կառուցվածքի ձևավորմանը, առանձին գեների տրանսկրիպցիայի կարգավորմանը, տրանսպոզոնների արտահայտման ճնշմանը և հետերոքրոմատինի ընդլայնված շրջանների ֆունկցիոնալ կառուցվածքի պահպանմանը:
Փոքր ՌՆԹ-ների մի քանի հիմնական տեսակներ կան. Լավագույն ուսումնասիրվածներն են միկրո-ՌՆԹ-ները (miRNAs) և փոքր միջամտող ՌՆԹ-ները (siRNAs): Բացի այդ, փոքր ՌՆԹ-ների շարքում ուսումնասիրվում են հետևյալը. փոքր միջամտող ՌՆԹ-ներ, որոնք կապված են էնդոգեն ռետրոտրանսպոզոնների և կրկնվող տարրերի հետ (տեղական/գլոբալ հետերոքրոմատիզացիայով - սկսած սաղմի առաջացման վաղ փուլերից; պահպանել տելոմերների մակարդակը), Drosophia rasiRNAs; հաճախ կոդավորված է սպիտակուցային գեների ինտրոններով և ֆունկցիոնալորեն կարևոր թարգմանության, տրանսկրիպցիայի, զուգավորման (դե-/մեթիլացում, նուկլեինաթթուների կեղծ-ուրիդիլացում) փոքր միջուկային (snRNAs) և nucleolar (snoRNAs) ՌՆԹ-ների համար; փոքր մոդուլացնող ՌՆԹ-ներ, smRNA-ներ, քիչ հայտնի գործառույթներով, որոնք լրացնում են ԴՆԹ կապող NRSE (Նեյրոնի սահմանափակող խլացուցիչ տարր) շարժառիթները. տրանսակտիվացնող փոքր խանգարող բույսերի ՌՆԹ-ներ, tasiRNAs; կարճ վարսահարդարիչ ՌՆԹ-ներ, shRNA-ներ, որոնք ապահովում են կենդանիների հակավիրուսային արձագանքման ժամանակ երկարատև RNAi (գեների կայուն լռեցում) երկարատև lncRNA կառուցվածքների:
Փոքր ՌՆԹ-ները (miRNAs, siRNAs և այլն) փոխազդում են նոր սինթեզված միջուկային/ցիտոպլազմային տառադարձումների հետ (կարգավորում է զուգավորումը, mRNA-ի թարգմանությունը; rRNA-ի մեթիլացում/կեղծուրիդիլացում և այլն) և քրոմատինի հետ (ժառանգական բջիջների ժամանակավոր տեղային և էպիգենետիկորեն ժառանգված հետերոխերմատիզացիա) . Հետերոքրոմատինացումը, մասնավորապես, ուղեկցվում է ԴՆԹ-ի դե-/մեթիլացմամբ, ինչպես նաև հիստոնների մեթիլացումով, ացետիլացումով, ֆոսֆորիլացմամբ և ուբիկվիտինացիայով («հիստոնային կոդի» փոփոխություն):
Փոքր ՌՆԹ-ներից առաջինը հայտնաբերվել և ուսումնասիրվել են Caenorhabditis elegans (lin-4) նեմատոդի միՌՆԹ-ները, դրանց հատկություններն ու գեները, իսկ որոշ ժամանակ անց՝ Arabidopsis thaliana բույսի miRNA-ները: Ներկայումս դրանք կապված են բազմաբջիջ օրգանիզմների հետ, թեև դրանք ցուցադրվում են Chlamydomonas reinhardtii-ի միաբջիջ ջրիմուռներում և լռեցնելու RNAiման ուղիներում՝ կապված հակավիրուսային/նմանատիպ պաշտպանության հետ՝ այսպես կոչված մասնակցությամբ: psiRNA-ները քննարկվում են պրոկարիոտների համար: Բազմաթիվ էուկարիոտների (այդ թվում՝ Drosophila-ի, մարդկանց) գենոմները պարունակում են miRNA-ների մի քանի հարյուր գեն: Այս փուլային/հյուսվածքային հատուկ գեները (ինչպես նաև mRNA թիրախների համապատասխան շրջանները) հաճախ խիստ հոմոլոգ են ֆիլոգենետիկորեն հեռավոր տեսակների մեջ, սակայն դրանցից մի քանիսը տոհմային են: ՄիՌՆԹ-ները պարունակվում են էկզոններում (սպիտակուցը կոդավորող, ՌՆԹ գեներ), ինտրոններում (առավել հաճախ՝ նախամՌՆԹ), միջգենային միջակայքերում (ներառյալ կրկնությունները), ունեն մինչև 70-120 նուկլեոտիդների երկարություն (և ավելի) և կազմում են հանգույցի մազակալ կառուցվածքներ / ցողունի տեսակը. Նրանց գեները որոշելու համար օգտագործվում են ոչ միայն կենսաքիմիական և գենետիկական, այլև համակարգչային մոտեցումներ։
Հասուն միՌՆԹ-ների «աշխատանքային շրջանի» ամենաբնորոշ երկարությունը 21-22 նուկլեոտիդ է։ Սրանք, հնարավոր է, ամենաառատ գեներն են ոչ սպիտակուցային կոդավորող գեների մեջ: Դրանք կարող են տեղակայվել առանձին պատճենների (ավելի հաճախ) կամ կլաստերների տեսքով, որոնք պարունակում են բազմաթիվ նմանատիպ կամ տարբեր miRNA գեներ, տառադարձվել (հաճախ ինքնավար խթանողներից) որպես ավելի երկար պրեկուրսոր, որը մի քանի փուլով մշակվել է առանձին miRNA-ներին: Ենթադրվում է, որ գոյություն ունի miRNA կարգավորող ցանց, որը վերահսկում է բազմաթիվ հիմնարար կենսաբանական գործընթացներ (ներառյալ ուռուցքի առաջացումը / մետաստազը); հավանաբար արտահայտված մարդու գեների առնվազն 30%-ը կարգավորվում են miRNA-ներով:
Այս գործընթացը ներառում է lncRNA-ին հատուկ RNase-III-ի նման ֆերմենտներ Drosha (միջուկային ռիբոնուկլեազ; սկսում է ինտրոնիկ նախամիՌՆԹ-ների մշակումը հիմնական տառադարձման միացումից հետո) և Dicer-ը, որը գործում է ցիտոպլազմայում և համապատասխանաբար ճեղքում/դեգրադացնում է մազակալը: ՄիՌՆԹ-ներ (միՌՆԹ-ների հասունացման համար) և հետագայում ձևավորեցին հիբրիդային միՌՆԹ/մՌՆԹ կառուցվածքներ: Փոքր ՌՆԹ-ները՝ մի քանի սպիտակուցների հետ միասին (այդ թվում՝ vn. RNases, AGO ընտանիքի սպիտակուցներ, տրանսմեթիլազներ/ացետիլազներ և այլն) և այսպես կոչված մասնակցությամբ. RISC- և RITS-ի նման բարդույթները (երկրորդը հրահրում է տրանսկրիպցիոն լռեցում) համապատասխանաբար ի վիճակի են առաջացնել RNAi / դեգրադացիա և հետագա գենի լռեցում ՌՆԹ-ում (նախքան / թարգմանության ընթացքում) և ԴՆԹ (հետերոքրոմատինի տրանսկրիպցիայի ընթացքում):
Յուրաքանչյուր miRNA-ն պոտենցիալ կերպով զուգակցվում է բազմաթիվ թիրախների հետ, և յուրաքանչյուր թիրախ վերահսկվում է մի շարք miRNA-ների կողմից (որը հիշեցնում է gRNAs-ի միջնորդավորված նախնական mRNA խմբագրումը տրիպանոսոմային կինետոպլաստներում): In vitro վերլուծությունը ցույց տվեց, որ miRNA-ների կարգավորումը (ինչպես ՌՆԹ-ի խմբագրումը) գեների արտահայտման հիմնական հետտրանսկրիպցիոն մոդուլատորն է: Նմանատիպ miRNA-ները, որոնք մրցում են մեկ թիրախի համար, ՌՆԹ-ՌՆԹ-ի և ՌՆԹ-սպիտակուցի փոխազդեցությունների պոտենցիալ տրանսկարգավորիչներ են:
Կենդանիների մոտ miRNA-ները լավագույնս ուսումնասիրվում են Caenorhabditis Elegans նեմատոդի համար; նկարագրված է ավելի քան 112 գեն։ Այստեղ հայտնաբերվել են նաև հազարավոր էնդոգեն siRNAs (գեներ չկան, կապված են, մասնավորապես, սպերմատոգենեզի միջնորդավորված տրանսկրիպտների և տրանսպոզոնների հետ): Բազմաբջջային օրգանիզմների երկու փոքր ՌՆԹ-ները կարող են առաջանալ ՌՆԹ պոլիմերազների կողմից, որոնք ցուցադրում են RdRP-II-ի (ինչպես մյուս ՌՆԹ-ների մեծ մասի) և RdRP-III տեսակների ակտիվությունը (ոչ հոմոլոգիա): Հասուն փոքր ՌՆԹ-ները նման են կազմով (ներառյալ 5-րդ տերմինալ «-ֆոսֆատները և 3» -OH), երկարությունը (սովորաբար 21-22 նուկլեոտիդ) և գործառույթը, և կարող են մրցակցել մեկ թիրախի համար: Այնուամենայնիվ, ՌՆԹ-ի քայքայումը և ամբողջական թիրախային կոմպլեմենտարությամբ ավելի հաճախ կապված է siRNA-ների հետ; թարգմանական ռեպրեսիա, մասնակի, սովորաբար 5-6 նուկլեոտիդներով, փոխլրացում միՌՆԹ-ների հետ; և պրեկուրսորները, համապատասխանաբար, էքսո/էնդոգեն են (հարյուր/հազարավոր նուկլեոտիդներ) siRNA-ների համար, և սովորաբար էնդոգեն (տասնյակ/հարյուրավոր նուկլեոտիդներ) miRNA-ների համար և դրանց կենսագենեզը տարբեր է. սակայն, որոշ համակարգերում այդ տարբերությունները շրջելի են:
RNAi-ն, որը միջնորդվում է siRNA-ների և miRNA-ների միջոցով, ունի մի շարք բնական դերեր. բայց siRNA-ները և որոշ miRNA-ները պահպանված չեն տեսակների միջև: Բույսերում (Arabidopsis thaliana) հայտնաբերվել են հետևյալը. siRNAs, որոնք համապատասխանում են ինչպես գեներին, այնպես էլ միջգենային (ներառյալ spacers, կրկնվող) շրջաններին. մեծ թվով պոտենցիալ գենոմային տեղամասեր տարբեր տեսակի փոքր ՌՆԹ-ների համար: Նեմատոդների մեջ այսպես կոչված. փոփոխական ինքնավար արտահայտված 21Y-RNAs (dasRNAs); նրանք ունեն 5 «-Y մոնոֆոսֆատ, կազմում են 21 նուկլեոտիդ (դրանցից 20-ը փոփոխական են) և գտնվում են սպիտակուցը կոդավորող գեների ինտրոնների միջև կամ ներսում՝ IV քրոմոսոմի երկու շրջանների ավելի քան 5700 տեղամասերում։
MiRNA-ները կարևոր դեր են խաղում առողջության և հիվանդությունների գեների արտահայտման մեջ. մարդկանց մոտ՝ այդ գեներից առնվազն 450-500-ը: Սովորաբար, կապվելով mRNA-ի 3 «-UTO-տարածքների հետ (այլ թիրախներ), նրանք կարող են ընտրովի և քանակապես (մասնավորապես, ցածր արտահայտված գեների արտադրանքը շրջանառությունից հեռացնելիս) արգելափակել որոշ գեների աշխատանքը և այլ գեների գործունեությունը: ՌՆԹ-ները (և դրանց թիրախները) դինամիկ կերպով փոխվում են օնտոգենեզի, բջիջների և հյուսվածքների տարբերակման ընթացքում: Այս փոփոխությունները հատուկ են, մասնավորապես, կարդիոգենեզի ընթացքում, դենդրիտների երկարության և նյարդային բջիջների սինապսների քանակի օպտիմալացման գործընթացում (մասնակցությամբ՝ miRNA-134, այլ փոքր ՌՆԹ-ներ) բազմաթիվ պաթոլոգիաների զարգացում (օնկոգենեզ, իմունային անբավարարություն, գենետիկ հիվանդություններ, պարկինսոնիզմ, Ալցհեյմերի հիվանդություն, ակնաբուժական խանգարումներ (ռետինոբլաստոմա և այլն), որոնք կապված են տարբեր բնույթի վարակների հետ) Հայտնաբերված miRNA-ների ընդհանուր թիվը շատ է աճում: ավելի արագ, քան նրանց կարգավորիչ դերի նկարագրությունը և կապը կոնկրետ թիրախների հետ ...
Համակարգչային վերլուծությունը կանխատեսում է հարյուրավոր mRNA թիրախներ առանձին miRNA-ների համար և առանձին mRNA-ների կարգավորումը բազմաթիվ miRNA-ների կողմից: Այսպիսով, miRNA-ները կարող են ծառայել նպատակային գեների տառադարձումները վերացնելու կամ դրանց արտահայտությունը տրանսկրիպցիոն/թարգմանական մակարդակներում ճշգրտելու նպատակին: Տեսական նկատառումները և փորձարարական արդյունքները հաստատում են miRNA-ների տարբեր դերերի առկայությունը:
Վերանայումը արտացոլված է էուկարիոտների աճի/զարգացման գործընթացներում և որոշ պաթոլոգիաների (ներառյալ քաղցկեղի էպիգենոմիկայի) հետ կապված փոքր ՌՆԹ-ների հիմնարար դերի հետ կապված ասպեկտների ավելի ամբողջական ցանկը:
Փոքր ՌՆԹ-ները ուռուցքաբանության մեջ
Ուռուցքների աճի, զարգացման, առաջընթացի և մետաստազիայի գործընթացները ուղեկցվում են շատերով էպիգենետիկ փոփոխություններվերածվելով ավելի հազվադեպ, մշտական ժառանգական գենետիկական փոփոխությունների: Հազվագյուտ մուտացիաները, սակայն, կարող են մեծ քաշ ունենալ (կոնկրետ անհատի համար՝ նոզոլոգիա), քանի որ առանձին գեների (օրինակ՝ APC, K-ras, p53) առնչությամբ, այսպես կոչված. «ձագարային» էֆեկտը՝ կապված քաղցկեղի գրեթե անդառնալի զարգացման/հետևանքների հետ։ Ուռուցքին հատուկ տարբեր գեների (սպիտակուցներ, ՌՆԹ, փոքր ՌՆԹ) արտահայտման պրոֆիլի նկատմամբ, նախածննդյան բջիջների տարասեռությունը պայմանավորված է էպիգենոմային վերադասավորվող կառուցվածքների կոնյուգացված տատանումներով: Էպիգենոմը մոդուլացվում է մեթիլացմամբ, հիստոնների հետթարգմանական փոփոխություններով / փոխարինմամբ (ոչ կանոնականների համար), գեների նուկլեոսոմային կառուցվածքի / քրոմատինի վերափոխմամբ (ներառյալ գենոմային տպագրությունը, այսինքն՝ ծնողական գեների ալելների արտահայտման դիսֆունկցիան և X ): Այս ամենը և փոքր ՌՆԹ-ներով կարգավորվող ՌՆԹ-ների մասնակցությամբ հանգեցնում է հետերոքրոմատինի (այդ թվում՝ հիպոմեթիլացված ցենտրոմերային) թերի կառուցվածքների առաջացմանը։
Գենային հատուկ մուտացիաների ձևավորմանը կարող է նախորդել հարյուր հազարավոր սոմատիկ կլոնային մուտացիաների հայտնի կուտակումը ոչ կոդավորող (հազվադեպ կոդավորող) շրջանի պարզ կրկնողություններում կամ միկրոարբանյակներում, գոնե միկրոարբանյակային մուտատոր ֆենոտիպով (MMP) ուռուցքներում: ; դրանք կազմում են կոլոռեկտալ, ինչպես նաև թոքերի, ստամոքսի, էնդոմետրիումի և այլն քաղցկեղի զգալի մասը: Անկայուն մոնո-/հետերոնուկլեոտիդային միկրոարբանյակը կրկնվում է (poly-A6-10, նմանատիպ), քան գենոմի կոդավորման (էկզոն) շրջաններում: միկրոարբանյակային-անկայուն, MSI +, ուռուցքներ: Թեև MS-կայուն / անկայուն շրջանների արտաքին տեսքի բնույթը և տեղայնացման մեխանիզմները լիովին պարզ չեն, MS-անկայունության ձևավորումը կապված է բազմաթիվ գեների մուտացիաների հաճախականության հետ, որոնք նախկինում չեն մուտացիայի ենթարկվել MSI + ուռուցքներում և, հավանաբար, , ուղղեց նրանց առաջընթացի ուղիները. Այս ուռուցքներում MSI կրկնվող մուտացիաների հաճախականությունը մեծացել է ավելի քան երկու կարգով: Ոչ բոլոր գեներն են վերլուծվել կրկնությունների առկայության համար, սակայն դրանց փոփոխականության աստիճանը կոդավորող/ոչ կոդավորող շրջաններում տարբեր է, և մուտացիաների հաճախականությունը որոշելու մեթոդների ճշգրտությունը հարաբերական է: Կարևոր է, որ MSI փոփոխական կրկնությունների համար ոչ կոդավորող շրջանները հաճախ երկալելային են, մինչդեռ կոդավորման շրջանները մոնալելային են:
Ուռուցքների մեջ մեթիլացիայի գլոբալ նվազումը բնորոշ է կրկնություններին, շարժական տարրերին (ME; դրանց տրանսկրիպցիան մեծանում է), խթանողներին, ուռուցքը ճնշող miRNA գեների CpG կայքերին և փոխկապակցված է պրոգրեսիվ քաղցկեղի բջիջներում ռետրոտրանսպոզոնների հիպերտրանգրման հետ: Սովորաբար, մեթիլոմի տատանումները կապված են ծնողական / փուլային / հյուսվածքային հատուկ մեթիլացման ալիքների և հետերոքրոմատինի կենտրոնամետ արբանյակային շրջանների ուժեղ մեթիլացման հետ, որը կարգավորվում է փոքր ՌՆԹ-ներով: Երբ արբանյակները թերմեթիլացված են, արդյունքում առաջացող քրոմոսոմային անկայունությունը ուղեկցվում է ռեկոմբինացիայի ավելացմամբ, և ME մեթիլացման խանգարումը կարող է առաջացնել դրանց արտահայտումը: Այս գործոնները նպաստում են ուռուցքի ֆենոտիպի զարգացմանը: Փոքր ՌՆԹ թերապիան կարող է լինել խիստ սպեցիֆիկ, բայց պետք է վերահսկվի, քանի որ Թիրախները կարող են լինել ոչ միայն անհատական, այլև բազմաթիվ mRNA / RNA մոլեկուլներ և քրոմոսոմների տարբեր (ներառյալ ոչ կոդավորող միջգենային կրկնությունները) շրջանների նոր սինթեզված ՌՆԹ-ներ:
Մարդու գենոմի մեծ մասը կազմված է կրկնություններից և ME-ից: Retrotransposon L1 (LINE տարրը) պարունակում է, ինչպես էնդոգեն ռետրովիրուսները, ռեվերտազ (RTase), էնդոնուկլեազ և պոտենցիալ ունակ է փոխանցել ոչ ինքնավար (Alu, SVA և այլն) ռետրոէլեմենտներ. L1 / նման տարրերը խլացվում են մեթիլացմամբ CpG տեղամասերում: Նկատի ունեցեք, որ գենոմի CpG տեղամասերի շարքում գեների խթանիչների CpG կղզիները թույլ մեթիլացված են, և 5-մեթիլցիտոզինը ինքնին պոտենցիալ մուտագեն հիմք է, որը դեամինացված է թիմին (քիմիապես կամ ՌՆԹ/(ԴՆԹ) խմբագրման, ԴՆԹ-ի վերանորոգման մասնակցությամբ): ; Այնուամենայնիվ, CpG որոշ կղզիներ ենթակա են չափազանց շեղված մեթիլացման, որն ուղեկցվում է ճնշող գեների ճնշումով և քաղցկեղի զարգացմամբ: Ավելին. L1-ով կոդավորված ՌՆԹ կապող սպիտակուցը, որը փոխազդում է AGO2 (Argo-naute ընտանիքի) և FMRP (փխրուն մտավոր հետամնացություն, էֆեկտոր RISC համալիրի սպիտակուց) սպիտակուցների հետ, նպաստում է L1-տարրի շարժմանը, ինչը ցույց է տալիս. RNAi համակարգերի հնարավոր փոխադարձ կարգավորում և մարդկային LINE տարրերի հետադարձ տեղադրում: Մասնավորապես, կարևոր է, որ Ալուի կրկնությունները կարողանան տեղափոխվել գեների ինտրոն/էկզոն տարածություն:
Այս և նմանատիպ մեխանիզմները կարող են բարձրացնել ուռուցքային բջիջների գենոմի պաթոլոգիական պլաստիկությունը: RTase-ի ճնշումը (կոդավորված, ինչպես էնդոնուկլեազը, L1 տարրերով; RTase-ն կոդավորված է նաև էնդոգեն ռետրովիրուսներով) RNAi մեխանիզմով ուղեկցվել է մի շարք քաղցկեղային բջիջների գծերի տարածման նվազմամբ և դիֆերենցիացիայի ավելացմամբ: L1 տարրի ներմուծումից հետո պրոտոնկոգեն կամ ճնշող գեն, նկատվել են ԴՆԹ-ի երկշղթա ընդմիջումներ: Սաղմնային տրակտի հյուսվածքներում (մկներ/մարդ) բարձրանում է L1 արտահայտման մակարդակը, և դրա մեթիլացումը կախված է piRNA-ից (26-30-bp)՝ կապված խլացման համակարգից, որտեղ PIWI սպիտակուցները մեծ ընտանիքի տարբերակներ են։ Argo-naute սպիտակուցներ, մուտացիաներ, որոնցում հանգեցնում են L1-ի դեմեթիլացման/դերեպրեսիայի/ նման տարրերի՝ երկար տերմինալ կրկնություններով: RasiRNA-ի խլացման ուղիները կապված են PIWI սպիտակուցների հետ ավելի մեծ չափով, քան Dicer-1/2 և Ago սպիտակուցների հետ: PiRNAs / siRNAs միջնորդավորված խլացման ուղիներն իրականացվում են ներմիջուկային մարմինների միջոցով, որոնք պարունակում են էվոլյուցիոն եղանակով պահպանված բազմապրոտեինային PcG համալիրներ, որոնց գործառույթները հաճախ խախտվում են ուռուցքային բջիջներում: Այս համալիրները պատասխանատու են երկարաժամկետ գործողության համար (ավելի քան 10 կբ/վ հետո, քրոմոսոմների միջև) և կարգավորում են HOX գեների կլաստերը, որոնք պատասխանատու են մարմնի պլանի համար։
Հակասենսաբանական թերապիայի նոր սկզբունքներ կարող են մշակվել՝ հաշվի առնելով գիտելիքները ավելի բարձր սպեցիֆիկ (քան ԴՆԹ-ի/սպիտակուցի մեթիլացման հիստոն-մոդիֆիկացնող ինհիբիտորները) հակաուռուցքային էպիգենոմիկ նյութերի, էպիգենոմիկ ՌՆԹ-ի լռեցման հիմնարար հիմքերը և փոքր ՌՆԹ-ների դերը քաղցկեղածինության մեջ:
Միկրո-ՌՆԹ-ն ուռուցքաբանության մեջ
Հայտնի է, որ ուռուցքի աճի և մետաստազիայի աճը կարող է ուղեկցվել որոշ անհատների / միՌՆԹ-ների արտահայտման նվազմամբ (Աղյուսակ 1): Նրանցից ոմանք կարող են ունենալ պատճառահետևանքային դեր ուռուցքի առաջացման գործում. և նույնիսկ նույն miRNA-ները (ինչպես miR-21/-24) տարբեր ուռուցքային բջիջներում կարող են դրսևորել ինչպես օնկոգեն, այնպես էլ ճնշող հատկություններ: Մարդու չարորակ ուռուցքի յուրաքանչյուր տեսակ առանձնանում է իր «miRNA-դրոշմ»-ով, և որոշ miRNA-ներ կարող են գործել որպես օնկոգեններ, ուռուցքը ճնշողներ, բջիջների միգրացիայի, ներխուժման և մետաստազիայի նախաձեռնողներ: Հիվանդ հյուսվածքներում հաճախ հայտնաբերվում են հիմնական miRNA-ների կրճատված քանակ, որոնք հավանաբար ներգրավված են հակաքաղցկեղային պաշտպանական համակարգերում: Օնկոգենեզին մասնակցող MiRNA-ները (miRs) ձեւավորել են այսպես կոչված հասկացությունը. Oncomyrah. ավելի քան 200 miRNA-ների արտահայտվածության վերլուծությունը լիմֆոմաների և պինդ քաղցկեղների ավելի քան 1000 նմուշներից թույլ տվեց հաջողությամբ դասակարգել ուռուցքները ենթատիպերի՝ ըստ դրանց ծագման և տարբերակման փուլի: ՄիՌՆԹ-ների գործառույթներն ու դերը հաջողությամբ ուսումնասիրվել են՝ օգտագործելով. հակա-miR-օլիգոնուկլեոտիդներ՝ փոփոխված (կյանքի տևողությունը մեծացնելու համար) 2 «-O-մեթիլ և 2» -O-մեթօքսիէթիլ խմբերում; ինչպես նաև LNA օլիգոնուկլեոտիդները, որոնցում 2 «և 4» դիրքերում գտնվող ռիբոզի թթվածնի ատոմները միացված են մեթիլենային կամրջով։
(էջանիշ 1) ………………….
Ուռուցք | miRNAs |
Թոքերի քաղցկեղ | 17-92 , թող-7 ↓ , 124ա ↓ , 126 ↓ , 143 ↓ , 145 ↓ , 155 , 191 , 205 , 210 |
Կրծքագեղձի քաղցկեղ | 21 , 125b ↓ , 145 ↓ , 155 |
Շագանակագեղձի քաղցկեղ | 15 ա ↓ , 16-1 ↓ , 21 , 143 ↓ ,145 ↓ |
Աղիների քաղցկեղ | 19 ա , 21 , 143 ↓ , 145 ↓ |
Ենթաստամոքսային գեղձի քաղցկեղ | 21 , 103 , 107 , 155 v |
Ձվարանների քաղցկեղ | 210 |
Քրոնիկ լիմֆոցիտային լեյկոզ | 15 ա ↓ , 16-1 ↓ , 16-2 , 23 բ , 24-1 , 29 ↓ , 146 , 155 , 195 , 221 , 223 ↓ |
Աղյուսակ 1 .
miRNAs, որոնց արտահայտությունը մեծանում է () կամ նվազում ( ↓ ) որոշ ավելի տարածված ուռուցքներում՝ համեմատած նորմալ հյուսվածքների հետ (տես նաև):
Ենթադրվում է, որ միՌՆԹ գեների արտահայտման, անհետացման և ուժեղացման կարգավորիչ դերը ուռուցքների մեծամասնության սկզբնավորման, աճի և առաջընթացի նախատրամադրվածության մեջ էական է, և miRNA/mRNA թիրախային զույգերի մուտացիաները սինխրոնիզացված են: ՄիՌՆԹ-ների արտահայտման պրոֆիլը կարող է օգտագործվել ուռուցքաբանության մեջ դասակարգման, ախտորոշման և կլինիկական կանխատեսման համար: ՄիՌՆԹ-ի արտահայտման փոփոխությունները կարող են ազդել բջջային ցիկլի, բջջի գոյատևման ծրագրի վրա: ՄիՌՆԹ-ների մուտացիաները ցողունային և սոմատիկ բջիջներում (ինչպես նաև mRNA թիրախների պոլիմորֆ տարբերակների ընտրությունը) կարող են նպաստել կամ նույնիսկ կարևոր դեր խաղալ շատ (եթե ոչ բոլոր) չարորակ նորագոյացությունների աճի, առաջընթացի և պաթոֆիզիոլոգիայի մեջ: ՄիՌՆԹ-ներով հնարավոր է ապոպտոզի ուղղում:
Բացի առանձին miRNA-ներից, հայտնաբերվել են դրանց կլաստերներ, որոնք գործում են որպես օնկոգեն, որը հրահրում է փորձարարական մկների մոտ, մասնավորապես, արյունաստեղծ հյուսվածքի քաղցկեղի զարգացումը. ՄիՌՆԹ-ների գեները օնկոգեն և ճնշող հատկություններով կարող են տեղակայվել մեկ կլաստերում: Ուռուցքներում miRNA-ների արտահայտման պրոֆիլների կլաստերային վերլուծությունը հնարավորություն է տալիս որոշել դրա ծագումը (էպիթել, արյունաստեղծ հյուսվածք և այլն) և դասակարգել նույն հյուսվածքի տարբեր ուռուցքները՝ փոխակերպման ոչ նույնական մեխանիզմներով: ՄիՌՆԹ-ների արտահայտման պրոֆիլը կարելի է գնահատել նանո / միկրոզանգվածների միջոցով. Նման դասակարգման ճշգրտությունը, տեխնոլոգիան մշակելիս (ինչը հեշտ չէ), պարզվում է, որ ավելի բարձր է, քան mRNA պրոֆիլներ օգտագործելիս: ՄիՌՆԹ-ներից մի քանիսը ներգրավված են արյունաստեղծ բջիջների (մուկ, մարդ) տարբերակման մեջ՝ նախաձեռնելով քաղցկեղի բջիջների առաջընթացը։ Մարդու miRNA գեները հաճախ տեղակայված են այսպես կոչված. «Փխրուն» տեղամասեր, տարածքներ, որտեղ գերակշռում են ջնջումները/ներդիրները, կետային ընդմիջումները, տրանսլոկացիաները, փոխադրումները, նվազագույն ջնջված և ուժեղացված հետերոքրոմատինային շրջանները, որոնք ներգրավված են օնկոգենեզում:
Անգիոգենեզ ... ՄիՌՆԹ-ների դերը անգիոգենեզում, հավանաբար, նշանակալի է: Myc-ակտիվացված մարդու որոշ ադենոկարցինոմայի անգիոգենեզի աճը ուղեկցվել է որոշ miRNA-ների արտահայտման ձևի փոփոխությամբ, իսկ այլ miRNA-ների գեների անկումը հանգեցրել է ուռուցքի աճի թուլացման և ճնշման: Ուռուցքի աճն ուղեկցվել է K-ras, Myc և TP53 գեների մուտացիաներով, անգիոգեն VEGF-գործոնի արտադրության ավելացմամբ և Myc-ի հետ կապված անոթայինացման աստիճանով; հակաանգիոգեն Tsp1 և CTGF գործոնները արգելակվել են miR-17-92-ով և այլ կլաստերային կապակցված miRNA-ներով: Ուռուցքների անգիոգենեզը և անոթայինացումը ուժեղացել են (մասնավորապես, կոլոնոցիտներում) երկու օնկոգենների համակցված արտահայտման արդյունքում մեկից ավելի մեծ չափով:
LATS2 հակաանգիոգեն գործոնի չեզոքացումը՝ կենդանական ցիկլինից կախված կինազայի (CDK2, մարդ/մուկ) արգելակիչ, օգտագործելով miRNAs-372/373 («պոտենցիալ օնկոգեններ») խթանել ամորձիների ուռուցքների աճը՝ չվնասելով p53 գենը:
Անգիոգեն հատկությունների պոտենցիալ մոդուլատորներ (in-vitro/in-vivo) miR-221/222-ն են, որոնց թիրախները՝ c-Kit ընկալիչները (մյուսները), հանդիսանում են պորտալարի էնդոթելային երակային HUVEC բջիջների անգիոգենեզի գործոններ և այլն: Այս miRNAs և c-Kit-ը փոխազդում են բարդ ցիկլի շրջանակներում, որը վերահսկում է էնդոթելային բջիջների՝ նոր մազանոթներ ձևավորելու ունակությունը:
Քրոնիկ լիմֆոցիտային լեյկոզ (СLL): B-բջջային քրոնիկ լիմֆոցիտային լեյկոզով (CLL) մարդու քրոմոսոմի 13q14 հատվածում նկատվում է miR-15a / miR-16-1 (և այլոց) գեների արտահայտման մակարդակի նվազում՝ կառուցվածքային ամենատարածված աննորմալությունների վայրում (ներառյալ 30 կբ տարածքի ջնջումները), թեև գենոմը արտահայտում էր հարյուրավոր հասուն և մարդկային նախամիՌՆԹ-ներ: Երկու miRNA-ն էլ, որոնք պոտենցիալ արդյունավետ են ուռուցքային թերապիայի մեջ, պարունակում էին հակաապոպտոտիկ Bcl2 սպիտակուցի հակազգայուն հատվածներ, ճնշում էին դրա գերարտահայտումը/արտահայտումը, խթանում էին ապոպտոզի, բայց գրեթե/ամբողջովին բացակայում էին CLL բջիջների երկու երրորդում, որոնք «անհավասարակշռված էին»: Ցողունային/սոմատիկ բջիջներում հաջորդականացված miRNA-ների հաճախակի մուտացիաները հայտնաբերվել են 75 հիվանդներից 11-ի մոտ (14,7%), որոնք ընտանեկան նախատրամադրվածություն ունեն CLL-ի նկատմամբ (ժառանգման եղանակը անհայտ է), բայց ոչ 160 առողջ հիվանդների մոտ: Այս դիտարկումները վկայում են լեյկեմոգենեզում miRNA-ների անմիջական գործառույթի մասին: Ներկայումս ոչ բոլորն են հայտնի miRNAs գեների (և դրանց գործառույթների) արտահայտման մակարդակների և նորմալ/ուռուցքային բջիջների այլ գեների միջև փոխհարաբերությունների մասին:
ՓաստաթուղթՀամապատասխանություն. Դեմքի նյարդի դիսֆունկցիան պարոտիդ թքագեղձի վիրահատության ժամանակ հրատապ խնդիրներից է և պայմանավորված է ինչպես հիվանդության տարածվածությամբ, այնպես էլ զգալի հաճախականությամբ:
Dawson եկեղեցի - հանճար ձեր գեներում էպիգենետիկ բժշկություն և մտադրության նոր կենսաբանություն գիրք գրադարանից www e - puzzle ru գիրք գրադարանից www e - puzzle ru բովանդակության աղյուսակ
ԳիրքԷթիկա հոգևորություն ուռուցքաբանություն հիվ պ գարյաև * էնֆի ռեզյումե
ՓաստաթուղթԱյս հոդվածն արտացոլում է ուռուցքաբանության և ՄԻԱՎ վարակի հիմնախնդրի նոր հայացք՝ լեզվական ալիքային գենետիկայի (LWG) և Էական կոդավորման տեսության (ISC) լույսի ներքո՝ հիմնված ռուսաստանյան և այլ սոցիալ-մշակութային իրողությունների նյութի վրա:
Քաղցկեղի հետազոտական կենտրոնը և Բլոխին Օդինցովան Անաստասիա Սերգեևնան նոր քիմիաթերապիայի սխեմաներ արգանդի պարանոցի առաջադեմ և կրկնվող քաղցկեղի համար 14 01 12 - ուռուցքաբանություն
Թեզիս4.4. Ուրիդին գլյուկորոնիլտրանսֆերազայի (UGT1A1) իզոֆերմենտի գենի որոշում արգանդի վզիկի քաղցկեղով հիվանդների արյան շիճուկում, ովքեր ստացել են առաջին գծի իրինոտեկանի քիմիաթերապիա պլատինի ածանցյալներով 105
Մազակալներ ձևավորող փոքր ՌՆԹ-ները կամ մազակալներ ձևավորող կարճ ՌՆԹ-ները (shRNA կարճ մազակալ ՌՆԹ, փոքր մազակալ ՌՆԹ) ՌՆԹ-ի կարճ մոլեկուլներ են, որոնք երկրորդական կառուցվածքում ստեղծում են խիտ մազակալներ։ ShRNA-ն կարող է օգտագործվել արտահայտությունն անջատելու համար ... ... Վիքիպեդիա
ՌՆԹ պոլիմերազ- վերարտադրության ժամանակ T. aquaticus բջիջից: Ֆերմենտի որոշ տարրեր դառնում են թափանցիկ, իսկ ՌՆԹ-ի և ԴՆԹ-ի շղթաներն ավելի հստակ տեսանելի են: Մագնեզիումի իոնը (դեղին) գտնվում է ֆերմենտի ակտիվ տեղում։ ՌՆԹ պոլիմերազը ֆերմենտ է, որն իրականացնում է ... ... Վիքիպեդիա
ՌՆԹ միջամտություն- Մազակալներ պարունակող փոքր ՌՆԹ-ների առաքում, օգտագործելով վեկտորը, որը հիմնված է lentivirus-ի և ՌՆԹ-ի միջամտության մեխանիզմի վրա կաթնասունների բջիջներում ՌՆԹ-ի ինտերֆեյսի վրա (... Վիքիպեդիա
ՌՆԹ գեն- Ոչ կոդավորող ՌՆԹ-ն (ncRNA) ՌՆԹ-ի մոլեկուլներ են, որոնք չեն թարգմանվում սպիտակուցների: Նախկինում օգտագործված հոմանիշը՝ փոքր ՌՆԹ (smRNA, փոքր ՌՆԹ), ներկայումս չի օգտագործվում, քանի որ որոշ չկոդավորող ՌՆԹ-ներ կարող են լինել շատ ... ... Վիքիպեդիա
Փոքր միջուկային ՌՆԹ-ներ- (snRNA, snRNA) ՌՆԹ-ների դաս, որոնք հայտնաբերված են էուկարիոտային բջիջների միջուկում: Դրանք տառադարձվում են ՌՆԹ պոլիմերազ II կամ ՌՆԹ պոլիմերազ III և ներգրավված են այնպիսի կարևոր գործընթացներում, ինչպիսիք են զուգավորումը (ինտրոնների հեռացումը ոչ հասուն mRNA-ից), կարգավորումը ... Վիքիպեդիա
Փոքր միջուկային ՌՆԹ-ներ- (snoRNA, անգլերեն snoRNA) փոքր ՌՆԹ-ների դաս, որոնք ներգրավված են ռիբոսոմային ՌՆԹ-ների, ինչպես նաև tRNA և փոքր միջուկային ՌՆԹ-ների քիմիական մոդիֆիկացիաներում (մեթիլացում և կեղծուրիդիլացում): Համաձայն MeSH դասակարգման՝ փոքր միջուկային ՌՆԹ-ները համարվում են ենթախումբ ... ... Վիքիպեդիա
փոքր միջուկային (ցածր մոլեկուլային քաշի միջուկային) ՌՆԹ- Փոքր միջուկային ՌՆԹ-ների (100 300 նուկլեոտիդների) ընդարձակ խումբ (105 106)՝ կապված տարասեռ միջուկային ՌՆԹ-ի հետ, միջուկի փոքր ռիբոնուկլեոպրոտեինային հատիկներն են. M.Ya.RNA-ն զուգավորման համակարգի անհրաժեշտ բաղադրիչն է...
փոքր ցիտոպլազմային ՌՆԹ- Փոքր (100-300 նուկլեոտիդ) ՌՆԹ-ի մոլեկուլները տեղայնացված են ցիտոպլազմայում, նման փոքր միջուկային ՌՆԹ-ին: [Arefiev V.A., Lisovenko L.A. Անգլերեն ռուսերեն Բառարանգենետիկական տերմիններ 1995 407s.] Գենետիկայի թեմաներ EN scyrpssmall cytoplasmic ... ... Տեխնիկական թարգմանչի ուղեցույց
փոքր միջուկային ՌՆԹ դասի U- սպիտակուցի հետ կապված փոքր (60-ից 400 նուկլեոտիդ) ՌՆԹ մոլեկուլների խումբ, որոնք կազմում են սպլայսի պարունակության զգալի մասը և մասնակցում են ինտրոնների հեռացման գործընթացին. Usn RNA U1, U2, U4 և U5 լավ ուսումնասիրված 5 տեսակներից 4-ն ունեն 5 ... ... Տեխնիկական թարգմանչի ուղեցույց
ՌՆԹ բիոմարկերներ- * RNA biomarkers * RNA biomarkers մեծ գումարմարդկային տառադարձումներ, որոնք չեն կոդավորում սպիտակուցի սինթեզը (nsbRNA կամ npcRNA): Շատ դեպքերում փոքր (miRNA, snoRNA) և երկար (հակազգայական ՌՆԹ, dsRNA և այլ տեսակներ) ՌՆԹ մոլեկուլները ... ... Գենետիկա. Հանրագիտարանային բառարան
Գրքեր
- Գնել 1877 UAH (միայն Ուկրաինա)
- Կլինիկական գենետիկա. Դասագիրք (+ CD), Բոչկով Նիկոլայ Պավլովիչ, Պուզիրև Վալերի Պավլովիչ, Սմիրնիխինա Սվետլանա Անատոլևնա. Բոլոր գլուխները վերանայվել և լրացվել են՝ կապված բժշկական գիտության և պրակտիկայի զարգացման հետ: Բազմագործոնային հիվանդությունների, ժառանգական հիվանդությունների կանխարգելման, բուժման, ...