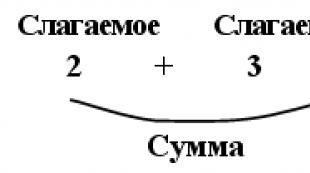
Lõhustumisspindli mikrotuubulid. mitoos - rakkude jagunemise aparaat Osaleb jagunemisspindli moodustamises
b) osmootne rõhk rakus d) selektiivne läbilaskvus
2. Tselluloosi ja kloroplastide membraanidel ei ole rakke
a) vetikad b) samblad c) sõnajalad d) loomad
3. Rakus paiknevad tuum ja organellid
a) tsütoplasma _ c) endoplasmaatiline retikulum
b) Golgi kompleks d) vakuoolid
4. Granulaarse endoplasmaatilise retikulumi membraanidel toimub süntees
a) valgud b) süsivesikud c) lipiidid d) nukleiinhapped
5. Tärklis koguneb sisse
a) kloroplastid b) tuum c) leukoplastid d) kromoplastid
6. Valgud, rasvad ja süsivesikud kogunevad sisse
a) tuum b) lüsosoomid c) Golgi kompleks d) mitokondrid
7. Kaasatud on lõhustumisspindli moodustumine
a) tsütoplasma b) rakukeskus c) vakuool d) Golgi kompleks
8. Organoid, mis koosneb paljudest omavahel ühendatud õõnsustest, sisse
mis akumuleeruvad rakus sünteesituna orgaaniline aine- see
a) Golgi kompleks c) mitokondrid
b) kloroplast d) endoplasmaatiline retikulum
9. Ainevahetus raku ja selle keskkonna vahel toimub läbi
kest selles sisaldumise tõttu
a) lipiidimolekulid c) süsivesikute molekulid
b) arvukalt auke d) nukleiinhappemolekule
10.Rakus sünteesitud orgaanilised ained liiguvad organellidesse
a) kasutades Golgi kompleksi c) kasutades vakuoole
b) lüsosoomide abil d) endoplasmaatilise retikulumi kanalite kaudu
11. Orgaanilise aine lagunemine rakus, millega kaasneb vabanemine.
aastal toimub suure hulga ATP molekulide energia ja süntees
a) mitokondrid b) lüsosoomid c) kloroplastid d) ribosoomid
12. Organismid, mille rakkudes puudub moodustunud tuum, mitokondrid,
Golgi kompleks, kuuluvad rühma
a) prokarüootid b) eukarüootid c) autotroofid d) heterotroofid
13. Prokarüootide hulka kuuluvad
a) vetikad b) bakterid c) seened d) viirused
14. Tuum mängib rakus olulist rolli, kuna osaleb sünteesis
a) glükoos b) lipiidid c) kiudained d) nukleiinhapped ja valgud
15. Organoid, tsütoplasmast piiritletud ühe membraaniga, sisaldades
palju ensüüme, mis lagundavad keerulisi orgaanilisi aineid
lihtsatele monomeeridele, see on
a) mitokondrid b) ribosoom c) Golgi kompleks d) lüsosoom
Abi pliz A1. Spindli filamentide kinnitumine toimub: 1) interfaasis 2) profaasis 3) metafaasis 4) anafaasis. A2. Profaasis mitoosi ei esinedit: 1) tuumaümbrise lahustumine 2) lõhustumisspindli moodustumine 3) DNA kahekordistumine 4) tuumade lahustumine. A3) loomadel tekivad mitoosi ajal erinevalt meioosist rakud: 1) somaatilised 2) poole kromosoomikomplektiga 3) sugu 4) spoorirakud. A4) kromatiidide lahknemine raku poolustele toimub: 1) meioosi esimese jagunemise profaasis 2) meioosi teise jagunemise profaasis 3) interfaasis enne esimest jagunemist 4) interfaasis enne teist jagunemist
1. Tärkliskoguneb sisse
A
- kloroplastid B - tuum C - leukoplastid D - kromoplastid
2. Tsütoplasma ei täida
funktsiooni
A
- ainete liikumine B - kõigi organellide vastastikmõjud
V
- toiteallikas G - kaitsev
3. Varu
toitained ja jääkained kogunevad taimerakkudesse
A
- lüsosoomid B - kloroplastid C - vakuoolid D - tuum
4. Valgud,
rasvad ja süsivesikud oksüdeeritakse koos energia vabanemisega
A
- mitokondrid B - leukoplastid
V
- endoplasmaatiline retikulum G - Golgi kompleks
5. "Kokkupanek"
ribosoom esineb
A
- endoplasmaatiline retikulum B - Golgi kompleks
V
- tsütoplasma G - tuumad
6. Sileda endoplasmaatilise retikulumi pinnal sünteesitakse molekulid A - mineraalsoolad B - nukleotiidid C - süsivesikud, lipiidid G - valgud.
7. Kareda endoplasmaatilise retikulumi pinnal A - lüsosoomid B - mikrotuubulid C - mitokondrid D - ribosoomid
8. Eukarüootid on organismid, millel on A - plastiidid B - lipukesed C - rakumembraan D - moodustunud tuum
9. Rakk on kõigi organismide ehituse põhiüksus, kuna A - organismide paljunemine põhineb rakkude jagunemisel B - rakus toimuvad metaboolsed reaktsioonid C - raku jagunemine on organismi kasvu aluseks D - kõik organismid koosnevad rakkudest
10. A - tsütoplasma B - rakukeskus C - endoplasmaatiline retikulum D - vakuool osaleb lõhustumisspindli moodustamisel
Ja rakkude jagunemine. Tüüpiline spindel on bipolaarne – kahe pooluse vahele moodustub spindlikujuline mikrotuubulite süsteem. Spindli mikrotuubulid kinnituvad kromatiidide kinetokooridele tsentromeeri piirkonnas ja tagavad kromosoomide liikumise pooluste suunas.
Spindli moodustavad kolm peamist struktuurielementi: mikrotuubulid, poolused ja kromosoomid. Loomade jagunemispooluste organiseerimine hõlmab tsentrosoome, mis sisaldavad tsentrioole. Taimedel, aga ka mõnede loomade munarakkudes tsentrosoomid puuduvad ja moodustub laiade poolustega atsentrosomaalne spindel. Spindli moodustamisel mängivad olulist rolli düneiini ja kinesiini perekonda kuuluvad motoorsed valgud.
Täielik lõhustumise spindel moodustub prometafaasi staadiumis pärast tuumamembraani hävimist, kui tsütoplasmaatilised mikrotuubulid ja tsentrosoomid (loomadel) pääsevad ligi kromosoomidele ja teistele spindli komponentidele. Erandiks on tärkava pärmi jagunemisvõll, mis moodustub tuuma sees.
Struktuur
Tüüpilise imetajaraku jagunemisspindel koosneb kolmest konstruktsioonielemendid- tsentrosoomid, mikrotuubulid ja kromosoomid - mis moodustavad sümmeetrilise bipolaarse struktuuri. Spindli poolustel on tsentrosoomid - väikesed organellid, mis toimivad mikrotuubulite organiseerimise keskustena. Iga tsentrosoom koosneb tsentrioolide paarist, mida ümbritsevad paljud erinevad valgud. Kondenseerunud kromosoomid paiknevad tsentromeeri piirkonnas omavahel seotud kromatiidide paarist koosneva spindli pooluste vahel. Kromosoomide tsentromeersetes piirkondades on kinetokoorid - keerulised struktuurid, mis vastutavad spindli kinnitamise eest mikrotuubulitele.
Lõhustumisspindel koosneb kahest poolspindlist. Poolspindel on moodustatud polariseeritud mikrotuubulitest. Mikrotuubulite negatiivsed miinusotsad kogunevad tsentrosoomi ümber asuvatele spindli poolustele. Mikrotuubulite plussotsad eemalduvad kahest poolusest ja lõikuvad spindli keskmises ekvaatoriosas. Enamikul selgroogsetel koosneb poolspindel 600–750 mikrotuubulist, millest 30–40% lõpevad kinetokooridega. Nimetatakse mikrotuubuleid, mis ühendavad spindli pooluseid kromosoomi kinetokooridega kinetokooriline... Lisaks seostub iga kinetokoor spindli moodustumise ajal paljude mikrotuubulitega ja moodustab kinetokoori kimbu. Nimetatakse mikrotuubuleid, mis paiknevad pooluste vahel ja ei kinnitu kinetokoore külge interpool... Mõned spindli mikrotuubulid moodustavad iga pooluse ümber radiaalseid struktuure, mida nimetatakse tähtedeks või astriteks. Neid mikrotuubuleid nimetatakse astraal .
Taimedel, aga ka mõnede loomade munarakkudes tsentrosoomid puuduvad ja moodustub laiade poolustega atsentrosomaalne spindel. Samuti puuduvad atsentrosomaalse spindli poolustes astraalsed mikrotuubulid. Vastasel juhul vastab taimeraku võlli ehitus loomaraku spindli ehitusele.
Jagamisspindli koost
Spindli kokkupaneku algus profaasis
Lõhustumisspindli kokkupanek algab profaasis. Kuid selles etapis on täisväärtusliku spindli moodustumine võimatu kromosoomide, aga ka tuuma sees olevate oluliste motoorsete, reguleerivate ja stabiliseerivate valkude eraldamise tõttu.
Taimedes mängib tsentrosoomide puudumise tõttu tuumaümbris profaasis mikrotuubulite organiseerimiskeskuse rolli. Mikrotuubulid kogunevad tuuma pinna lähedale ja on profaasi lõpuks orienteeritud piki tulevase lõhustumisspindli telge, moodustades nn profaasi spindli.
Loomarakkudes on tsentrosoom mikrotuubulite organiseerimise keskus. Seetõttu algab lõhustumisspindli moodustumine tsentrosoomide paari eraldumisest ja lahknemisest profaasi ajal. Tsentrosoomide lahknemise profaasis tagavad motoorsete valkude düneiinid. Need on fikseeritud seestpoolt rakumembraan ja südamiku välispinnal. Membraanis fikseeritud dyneiinid kinnituvad astraalmikrotuubulitele ja liiguvad mikrotuubuli miinusotsa poole. Tänu sellele liiguvad tsentrosoomid rakumembraani vastandlikesse osadesse ja lahknevad üksteisest kaugemale.
Spindli kokkupanek prometafaasis
Spindli isekorraldus:
Erandiks on tärkava pärmi jagunemisvõll, mis moodustub tuuma sees.
Spindli iseorganiseerumine
Kõigis eukarüootides sõltub bipolaarse spindli kokkupanek suuresti spindli komponentide iseorganiseerumisvõimest. Iseorganiseerumine on ainus mehhanism jagunemisspindli kokkupanekuks rakkudes, kus puuduvad tsentrosoomid. Bipolaarse spindli koostu ilma tsentrosoomi kaasamiseta nimetatakse atsentrosomaalseks. See on iseloomulik kõrgematele taimedele ja seda täheldatakse ka mõnede loomade meioosi puhul varases arengujärgus. Veelgi enam, arvatakse, et mikrotuubulite iseorganiseerumine on spindli koostamise domineeriv mehhanism isegi tsentrosoome sisaldavates loomarakkudes.
Spindli iseorganiseerumine algab pärast tuumamembraani hävimist. Tsütoplasmaatilised mikrotuubulid kogunevad (tuumad) kromosoomide ümber. Siin pikendatakse kohalike stabiliseerivate tegurite osalusel akumuleeruvaid mikrotuubuleid. Järgmisena algab mikrotuubulite organiseerimine kolme motoorsete valkude rühma osalusel:
- Perekonna motoorsed valgud kinesiin-5(Eg5) seostuvad kahe vastassuunas orienteeritud mikrotuubuliga ja liiguvad samaaegselt mõlema pluss-otsa poole. Selle tulemusena sorteeritakse ja "õmmeldakse" pluss-otsa piirkonnas antiparalleelsed polariseeritud mikrotuubulid.
- Kromokinesiinid - kinesiin-4 ja -10 perekonna valgumootorid, mis paiknevad kromosoomide harudel, seovad kromosoomide läheduses paiknevaid mikrotuubuleid ja liiguvad mikrotuubuli pluss-otsa suunas. Seega on kromosoomi õlg ühendatud mikrotuubuli pluss-otsaga ja miinus-ots on kromosoomist eemal.
- Kolmas motoorsete valkude rühm liigub mikrotuubulite miinusotste suunas ja annab spindli pooluste juures miinusotste kimbu. Sellesse mootorite rühma kuuluvad tsütoplasmaatilised düneiine, kinesiin-14. Dynein osaleb näiteks lõhustumise pooluste fokuseerimises koos paljude tuumavalkudega NuMA1(ing. Nu selge M ikrotuubul - A seotud valk 1).
Kokkupanek tsentrosoomi osalusel
Paljudes loomarakkudes, sealhulgas inimese rakkudes, osalevad spindli koostamises tsentrosoomid, mis on jaotusvõlli poolused. Nagu atsentrosomaalse spindli koostamisel, osalevad motoorsed ja teised valgud mikrotuubulite iseorganiseerumises bipolaarseks struktuuriks, mille fokusseerivad tsentrosoomide piirkonnas mikrotuubulite miinusotsad. Sel juhul osalevad tsentrosoomid ka spindli kokkupanemises ja aitavad kaasa jaotuspooluste moodustumisele, kuid need ei ole spindli lahutamatu osa, kuna koosteprotsess võib jätkuda isegi tsentrosoomide inaktiveerimisel.
Sõltuvalt tsentrosoomide lahknemise ajast tuumaümbrise hävimise hetke suhtes eristatakse kahte spindli moodustumise mehhanismi:
- Kui tuumaümbris hävitatakse enne, kui tsentrosoom hakkab lahknema, jaotuvad vabanenud kromosoomid tsütoplasmas ja moodustub "unipolaarne" spindel, mille mikrotuubulid lahknevad paaritud tsentrosoomist. Bipolaarse spindli edasine moodustumine toimub kattuvate mikrotuubulite tõukejõudude tõttu ja astraalsete mikrotuubulite tõmbejõudude toimel. Kattuvate mikrotuubulite vahelise tõukejõu loovad kinesiinitaolised valgud Eg5. Astraalmikrotuubulitele rakendatavad tõmbejõud tekitavad rakumembraani sisepinnale kinnitatud tsütoplasmaatilised düneiinid.
- Teine võimalus on seotud tsentrose lahknemisega ja primaarse spindli moodustumisega enne tuumaümbrise hävitamist. Primaarne spindel moodustub astraalsete mikrotuubulite tõmbejõudude toimel, mis tekivad tsütoplasmaatiliste düneiinide poolt, mis on fikseeritud rakumembraani sisepinnale ja tuumaümbrise pinnale. Tsentrosoomide lahknemise suuna määravad aktiini filamendid, mis interakteeruvad müosiiniga, mis paikneb tsentrosoomides endis või piki mikrotuubuleid. Esmane spindel on ebastabiilne. Selle stabiilsuse tagamiseks on vaja suhelda kromosoomide ja teiste raku tuumas asuvate valkude kinetokooridega.
Kromosoomide kinnitumine spindlile
Enim uuritud kromosoomide spindli külge kinnitumise mehhanism tsentrosoome sisaldavates loomarakkudes. Profaasi ajal moodustub tsentrosoomi ümber radiaalsuunas lahknevate mikrotuubulite tähtstruktuur. Pärast tuumamembraani hävitamist uurivad tuuma piirkonda aktiivselt dünaamiliselt ebastabiilsed mikrotuubulid, mille kinni püüavad kromosoomide kinetokoorid. Mõned kromosoomid seostuvad kiiresti vastaspooluste mikrotuubulitega. Teine osa kromosoomidest kinnitub kõigepealt ühest poolusest väljuvatele mikrotuubulitele. Seejärel liigub see vastava pooluse suunas. Seejärel hõivavad ühe poolusega seotud kromosoomid vastaspooluse mikrotuubuleid. Metafaasi käigus kinnitub igale kinetokoorile umbes 10-40 mikrotuubulit, mis moodustavad kinetokoori kimbu. Kõik kromosoomid on ühendatud jagunemise vastaspoolustega ja koondatakse spindli keskele metafaasiplaadiks.
Kinetokooride spindli külge kinnitamiseks on olemas ka alternatiivne mudel, mis sobib nii tsentrosoomidega kui ka ilma tsentrosoomideta rakkude jaoks. Selle mudeli kohaselt toimub kromosoomide lähedal lühikeste mikrotuubulite tuumastumine gamma-tubuliini tsükli kompleksi osalusel. Oma plussotstega on mikrotuubulid kinnistatud kinetokooridesse. Sellele järgneb mikrotuubulite kontrollitud kasv (polümerisatsioon). Mikrotuubulite pikenevad miinusotsad on "õmmeldud" ja fokusseeritud jaotuspooluste piirkonda motoorsete valkude osalusel. Tsentrosoomid (kui need on olemas) hõlbustavad kinetokoorsete mikrotuubulite kinnitumist pooluste külge.
Õdekromatiidide bipolaarne orientatsioon
Kromosoomide võrdseks jaotumiseks tütarrakkude vahel on oluline, et paariskromatiidide kinetokoorid oleksid kinnitatud vastaspoolustelt lähtuvate mikrotuubulite külge. Tavalist bipolaarset kinetokooride kinnitust vastaspoolustele nimetatakse amfiteelne... Spindli kokkupanemise ajal võib aga esineda muid kromosoomide kinnitusi. Ühe kinetokoori kinnitumist ühe pooluse poolusele nimetatakse monoteelne... Ühe kromosoomi kahe kinetokoori liitumist korraga ühe jagunemispoolusega nimetatakse sünteetilised... See on ka võimalik merotelik kinnitus, milles üks kinetokoor ühendatakse korraga kahe poolusega.
Sobimatut kinnitumist takistab osaliselt õdekinetokooride geomeetria, mis asuvad kromosoomide tsentromeerse piirkonna vastaskülgedel. Lisaks on ebaregulaarsed kinnitused ebastabiilsed ja pöörduvad ning kinetokooride normaalne bipolaarne kinnitus on stabiilne. Stabiilne ühendus saavutatakse tänu jaotuse vastaspoolustelt tulevatele tõmbejõududele. Kinetokooride õige kinnitumise eest vastaspoolustele reguleeriva süsteemi põhikomponent on proteiinkinaas ISBN 978-0-9539181-2-6.
G1 faasi iseloomustab intensiivsete biosünteesiprotsesside taasalustamine, mis mitoosi ajal järsult aeglustub ja lühikeseks ajaks tsütokinees peatub täielikult. Kogu valgusisaldus suureneb selle faasi jooksul pidevalt. Enamiku rakkude jaoks on G1 faasis kriitiline punkt, nn restriktsioonipunkt. Selle läbimise ajal toimuvad rakus sisemised muutused, misjärel peab rakk läbima kõik järgnevad rakutsükli faasid. S- ja G2-faasi vahelise piiri määrab aine ilmumine - S-faasi aktivaator.
G2 faasi peetakse rakkude ettevalmistamise perioodiks mitoosi alguseks. Selle kestus on lühem kui ülejäänud perioodid. Selles sünteesitakse lõhustuvaid valke (tubuliin) ja täheldatakse kromatiini kondenseerumisel osalevate valkude fosforüülimist.
Profaas
Profaasi ajal toimub kaks paralleelset protsessi. See on järkjärguline kromatiini kondenseerumine, selgelt nähtavate kromosoomide ilmumine ja tuumade lagunemine, samuti jagunemisspindli moodustumine, mis tagab kromosoomide õige jaotumise tütarrakkude vahel. Need kaks protsessi on ruumiliselt eraldatud tuumaümbrisega, mis püsib kogu profaasi vältel ja hävib alles selle lõpus. Enamiku loomade ja mõnede taimerakkude mikrotuubulite organiseerimise keskus on rakukeskus või tsentrosoom. Interfaasilises rakus asub see tuuma küljel. Tsentrosoomi keskosas on kaks tsentriooli, mis on selle materjalis üksteise suhtes täisnurga all. Tsentrosoomi perifeersest osast väljuvad arvukad valgu tubuliini poolt moodustatud torukesed. Nad eksisteerivad ka faasidevahelises rakus, moodustades selles tsütoskeleti. Mikrotuubulid pannakse väga kiiresti kokku ja lahti. Need on ebastabiilsed ja nende massiivi uuendatakse pidevalt. Näiteks fibroblastirakkudes in vitro kultuuris on mikrotuubulite keskmine eluiga alla 10 minuti. Mitoosi alguses lagunevad tsütoplasmaatilised mikrotuubulid ja seejärel algab nende taastumine. Esiteks ilmuvad nad perinukleaarsesse tsooni, moodustades kiirgava struktuuri - tähe. Selle moodustumise keskpunkt on tsentrosoom. Mikrotuubulid on polaarsed struktuurid, kuna tubuliini molekulid, millest need moodustuvad, on teatud viisil orienteeritud. Selle üks ots pikeneb kolm korda kiiremini kui teised. Kiiresti kasvavaid otsi nimetatakse plussotsteks, aeglaselt kasvavateks miinusotsteks. Lisaks on otsad suunatud kasvu suunas ettepoole. Tsentriool on väike silindriline organell, mille paksus on umbes 0,2 mikronit ja pikkus 0,4 mikronit. Selle seina moodustavad üheksa torukujuliste kolmikute rühma. Kolmiku puhul on üks toruke terviklik ja kaks kõrvuti asetsevat tuubulit mittetäielikud. Iga kolmik on kallutatud kesktelje suunas. Kõrvuti asetsevad kolmikud on omavahel ühendatud ristsidumise teel. Uued tsentrioolid tekivad ainult olemasolevate kahekordistamisel. See protsess langeb kokku DNA sünteesi ajaga S-faasis. G1 perioodil liiguvad tsentrioolid, moodustades paari, mitme mikroni võrra eemale. Seejärel ehitatakse igale tsentrioolile, selle keskossa, täisnurga all tütartsentriool. Tütartsentrioolide kasv lõpeb G2 faasis, kuid need on siiski sukeldatud ühte tsentrosomaalse materjali massi. Profaasi alguses muutub iga tsentrioolide paar osaks eraldiseisvast tsentrosoomist, millest väljub radiaalne mikrotuubulite kimp – täht. Moodustunud tähed eemalduvad üksteisest mõlemal pool südamikku, muutudes seejärel lõhustumisspindli poolusteks.
Metafaas

Prometafaas algab tuumaümbrise kiire lagunemisega membraani fragmentideks, mida EPS fragmentidest ei eristata. Nad liiguvad kromosoomide ja jagunemisspindli abil raku perifeeriasse. Kromosoomide tsentromeeridele moodustub valgukompleks, mis elektroonilistel fotodel näeb välja nagu lamellne kolmekihiline struktuur – kinetokoor. Mõlemal kromatiidil on üks kinetokoor, mille külge on kinnitatud lõhustumisspindli valgu mikrotuubulid. Molekulaargeneetika meetodid on näidanud, et informatsioon, mis määrab kinetokooride spetsiifilise ehituse, sisaldub DNA nukleotiidjärjestuses tsentromeeri piirkonnas. Kromosoomide kinetokooride külge kinnitatud spindli mikrotuubulid mängivad väga olulist rolli; esiteks suunavad nad iga kromosoomi jagunemisspindli suhtes nii, et selle kaks kinetokoori on suunatud raku vastaspooluste poole. Teiseks liigutavad mikrotuubulid kromosoome nii, et nende tsentromeerid on raku ekvaatori tasapinnal. See protsess imetajarakkudes kestab 10–20 minutit ja lõpeb prometafaasi lõpuks. Iga kinetokooriga seotud mikrotuubulite arv on liigiti erinev. Inimestel on neid 20 kuni 40, pärmis - 1. Mikrotuubulite plussotsad seonduvad kromosoomidega. Lisaks kinetokoor-mikrotuubulitele sisaldab lõhuspind ka pooluste mikrotuubuleid, mis ulatuvad vastaspoolustelt ja on ekvaatoril spetsiaalsete valkudega kokku õmmeldud. Mikrotuubuleid, mis ulatuvad tsentrosoomist välja ja mis ei kuulu lõhustumisspindlisse, nimetatakse astraalseks, need moodustavad tähe.
Metafaas. Hõlmab olulise osa mitoosist. Seda on lihtne ära tunda kahe tunnuse järgi: lõhustumisspindli bipolaarne struktuur ja metafaasi kromosomaalne plaat. See on raku suhteliselt stabiilne olek; paljud rakud võivad jääda metafaasi mitmeks tunniks või päevaks, kui neid töödeldakse ainetega, mis depolümeriseerivad spindli tuubuleid. Pärast toimeaine eemaldamist on mitootiline spindel võimeline taastuma ja rakk suudab mitoosi lõpule viia.
Anafaas

Anafaas algab kõigi kromosoomide kiire sünkroonse lõhustumisega õdekromatiidideks, millest igaühel on oma kinetokoor. Kromosoomide lõhenemine kromatiidideks on seotud DNA replikatsiooniga tsentromeeri piirkonnas. Sellise väikese ala replikatsioon toimub mõne sekundiga. Signaal anafaasi alguse kohta pärineb tsütosoolist, see on seotud kaltsiumiioonide kontsentratsiooni lühiajalise kiire tõusuga 10 korda. Elektronmikroskoopia näitas, et spindli poolustele kogunevad kaltsiumirikkad membraani vesiikulid. Vastuseks anafaasi signaalile hakkavad sõsarkromatiidid liikuma pooluste suunas. Seda seostatakse esmalt kinetokoortorude lühenemisega (anafaas A) ja seejärel pooluste endi levikuga, mis on seotud polaarsete mikrotuubulite pikenemisega (anafaas B). Protsessid on suhteliselt iseseisvad, mida näitab nende erinev tundlikkus mürkide suhtes. Erinevates organismides on anafaasi A ja anafaasi B panus kromosoomide lõplikku eraldumisse erinev. Näiteks imetajarakkudes algab anafaas B pärast anafaasi A ja lõpeb siis, kui spindli pikkus on 1,5–2 korda pikem kui metafaasis. Algloomadel domineerib anafaas B, mille tõttu spindel pikeneb 15 korda. Kinetokoori tuubulite lühenemine toimub nende depolümerisatsiooni kaudu. Kaotatakse allüksused plussotsast, st. kinetokoori küljelt, selle tulemusena liigub kinetokoor koos kromosoomiga poolusele. Mis puutub pooluste mikrotuubulitesse. Seejärel pannakse need anafaasis kokku ja pikendatakse pooluste lahknemisel. Anafaasi lõpuks jagunevad kromosoomid raku poolustes täielikult kaheks identseks rühmaks.
Tuuma ja tsütoplasma jagunemine on omavahel seotud. Sel juhul mängib olulist rolli mitootiline spindel. Loomarakkudes tekib lõhustumisvagu juba anafaasis spindli ekvaatori tasapinnal. See asetatakse mitootilise spindli pikitelje suhtes täisnurga all. Soone moodustumine on tingitud kontraktiilse rõnga tegevusest, mis asub rakumembraani all. See koosneb kõige peenematest filamentidest – aktiini filamentidest. Kokkutõmbumisrõngal on piisavalt jõudu, et painutada puuri sisestatud õhuke klaasnõel. Soone süvenedes kokkutõmbumisrõnga paksus ei suurene, kuna osa filamentidest kaob selle raadiuse vähenemisega. Pärast tsütokineesi lõppu laguneb kontraktiilne ring täielikult, jaotusvao piirkonnas olev plasmamembraan tõmbub kokku. Mõnda aega säilib äsja moodustunud rakkude kontakttsoonis tihedalt pakitud mikrotuubulite jäänuste keha. Jäiga rakumembraaniga taimerakkudes jaguneb tsütoplasma uue seina moodustumisega tütarrakkude piiril. Taimerakkudes ei ole kokkutõmbuvat ringi. Raku ekvaatori tasapinnal moodustub fragmoplast, mis laieneb järk-järgult raku keskmest selle perifeeriasse, kuni kasvav rakuplaat jõuab emaraku plasmamembraanini. Membraanid sulanduvad, eraldades moodustunud rakud täielikult.
/. Spindli struktuur
2. Spindli funktsioonid. Keerme liikumise mehhanismid
1. Tuuma lõhustamisel raku kahe vastandpooluse vahel nn spindel, koosnevad:
Filamentidest (kiududest), mis on suure hulga mikrotuubulite (mõnikord üle 100) kimbud;
Kaks tsentriooli, millest igaüks on oma poolusel erinevate korralduskeskused:
Või peritsentriolaarse mikrotuubulite organiseerimiskeskusega (loomadel);
Või amorfse ("polaarkübar" enamikus taimedes);
Kas lamell- või kihiliste ("spindli polaarkehad" paljudes seentes ja osades alusmetsades).
Täiendav organiseerimiskeskus, kinetokoor, asub iga kromatiidi tsentromeeris. Eristama järgmist tüüpi spindli keermed:
Kromosomaalsed (kinetokoor või tõmbavad) filamendid, mis moodustuvad kinetokoorist ja ühendavad selle ühe poolusega;
Tsentraalsed filamendid, mis on moodustatud polaarsetest organiseerimiskeskustest ja ühendavad mõlemat poolust;
Polaarsed filamendid, mis tekivad ainult tsentrioolide juuresolekul peritsentriolaarsetes organiseerimiskeskustes ja lõpevad tsütoplasmas.
2. Funktsioonid lõhustumise spindel on järgmised:
Spindel tagab kromatiidide ehk kromosoomide lahknemise poolustele. Kromosoomi filamendid lühenevad ja tõmbavad kromosoome pooluste poole;
Loomadel tsentraalsed niidid tavaliselt pikenevad ja liiguvad poolused lahku. Sel juhul spindli keermete paksus ei muutu.
Mehhanismidniidi liigutused:
Spindli filamentide aktiivne libisemine toimub ilmselt düneiinitaolise valguga suhtlemisel. Mehhanism on sarnane flagella omaga;
Aktiivset rolli mängivad mikrokiud, mis kinnituvad spindli niitide külge ja tõmbavad nende abil kromaate ehk kromosoome üles. Spindli aparaadist leiti aktiini filamente ja müosiini. Tsütohanasiin, mis destabiliseerib aktiini filamente, on võimeline blokeerima spindli toimet.
Lõhustumisspindel LOOMADE EMBRÜOLOOGIA
KINNITUSLAINE, MITOOTILINE SPINNO - mikrotuubulite süsteem jagunevas rakus, mis tagab raku parameetrite muutused ja kromosoomide lahknemise mitoosis ja meioosis. Lõhustumisspindli teke lõpeb metafaasis, telofaasis laguneb. Lõhustumisspindel on valmistatud mikrotuubulite filamentidest. Filamente on kolme tüüpi: poolus - moodustab raku pooluste tsütoskeleti, pidev - ühendab raku poolused, moodustades tsütoskeleti, mis tõmbab rakku jagunemise ajal, ja katkendlikud filamendid, mis ühendavad kromosoomide tsentromeere raku poolustega. rakk.
Üldembrüoloogia: terminoloogiline sõnaraamat - Stavropol. O.V. Dilekova, T.I. Lapina. 2010 .
Vaadake, mis on "jaotusvõll" teistes sõnaraamatutes:
SPINDLI JAOTUS- SPREADY DIVISION, pulgakujuline mikrotuubulite süsteem raku tsütoplasmas MITOOSI või MEIOOSI protsessis. KROMOSOOMID kinnituvad lõhustumisspindli kühmule (ekvaatorile). Jagunemisvõll põhjustab kromosoomide eraldumist, põhjustades rakkude jagunemise. cm… Teaduslik ja tehniline entsüklopeediline sõnastik
lõhustumise spindel- (biol.), mikrotuubulite süsteem jagunevas rakus, mis tagab kromosoomide lahknemise ja rangelt identse (mitoosi ajal) jaotuse tütarrakkude vahel. * * * Spindli lõhustumine Spindli lõhustumine, bioloogias mikrotuubulite süsteem lõhustuvas ... ... entsüklopeediline sõnaraamat
spindel [lõhustumine]- akromaatiline spindel Mikrotuubulite süsteem jagunevas rakus, mis tagab kromosoomide lahknemise, eksisteerib perioodil metafaasist telofaasini; kaksikmurdvate omadustega mikrotuubulid moodustavad ... ... Tehniline tõlkija juhend
SPINDLI JAOTUS- akromatiini spindel, mikrotuubulite süsteem jagunevas rakus, mis tagab kromosoomide lahknemise mitoosis ja meioosis. V. d. Moodustub prometafaasis ja laguneb telofaasis. V. niidid, mis on mikrotuubulite kimbud, omavad ... ... Bioloogia entsüklopeediline sõnastik
SPINDLI JAOTUS- bioloogias mikrotuubulite süsteem jagunevas rakus, mis tagab kromosoomide lahknemise ja rangelt identse (mitoosi ajal) jaotuse tütarrakkude vahel ... Suur entsüklopeediline sõnaraamat
Lõhustumisspindel- Selles artiklis puuduvad lingid teabeallikatele. Teave peab olema kontrollitav, vastasel juhul saab selle kahtluse alla seada ja eemaldada. Saate ... Wikipedia
spindel [lõhustumine]- spindli spindel [jaotus], akromaatiline spindel. Mikrotuubulite süsteem jagunevas rakus, mis tagab kromosoomide lahknemise, eksisteerib perioodil metafaasist telofaasini; kaksikmurdvate omadustega mikrotuubulid moodustavad ... ... Molekulaarbioloogia ja geneetika. Sõnastik.
lõhustumise spindel- (fusus divisionis) rakustruktuur, mis tagab kromosoomide ühtlase eraldumise mitoosi või meioosi ajal; V. d. Tekib profaasis ja koosneb tsentraalsetest filamentidest, mis ühendavad raku mõlemat poolust, ja kromosomaalsetest filamentidest, mis ühendavad poolused ... Põhjalik meditsiiniline sõnastik
SPINDLI JAOTUS- (biol.), mikrotuubulite süsteem jagunevas rakus, mis tagab kromosoomide lahknemise ja rangelt identse (mitoosi ajal) jaotuse tütarrakkude vahel ... Loodusteadus. entsüklopeediline sõnaraamat
SPINDLI JAOTUS- (spindel) mikrotuubulitest ja nendega seotud valkudest koosnev struktuur; tekkinud mitoosi käigus (In prophase) kahe cntrioolipaari vahel (toim.). Mikrotuubulid ulatuvad välja raku poolustelt ja kohtuvad ekvaatoril, ... ... Arstiteaduse seletav sõnaraamat