Mikrotubule fisionog vretena. mitoza - aparat diobe ćelije Učestvuje u formiranju diobenog vretena
b) osmotski pritisak u ćeliji d) selektivna permeabilnost
2. Membrane celuloze i hloroplasta nemaju ćelije
a) alge b) mahovine c) paprati d) životinje
3. U ćeliji se nalazi jezgro i organele
a) citoplazma _ c) endoplazmatski retikulum
b) Golgijev kompleks d) vakuole
4. Na membranama granularnog endoplazmatskog retikuluma dolazi do sinteze
a) proteini b) ugljeni hidrati c) lipidi d) nukleinske kiseline
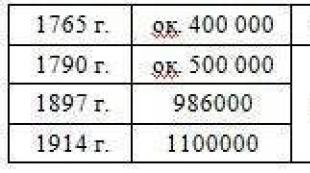
5. Skrob se akumulira u
a) hloroplasti b) jezgra c) leukoplasti d) hromoplasti
6. Proteini, masti i ugljeni hidrati se skladište u
a) jezgro b) lizozomi c) Golgijev kompleks d) mitohondrije
7. Uključeno je formiranje fisijskog vretena
a) citoplazma b) ćelijski centar c) vakuola d) Golgijev kompleks
8. Organoid, koji se sastoji od mnogih međusobno povezanih šupljina, u
koji se akumuliraju sintetizirani u ćeliji organska materija- ovo je
a) Golgijev kompleks c) mitohondrije
b) hloroplast d) endoplazmatski retikulum
9. Razmjena supstanci između ćelije i njene okoline se odvija putem
školjke zbog prisustva u njoj
a) molekule lipida c) molekule ugljikohidrata
b) brojne rupe d) molekule nukleinske kiseline
10 organskih supstanci sintetiziranih u ćeliji prelazi u organele
a) korištenjem Golgijevog kompleksa c) korištenjem vakuola
b) uz pomoć lizosoma d) kroz kanale endoplazmatskog retikuluma
11. Razgradnja organske materije u ćeliji, praćena oslobađanjem.
energija i sinteza velikog broja ATP molekula se odvija u
a) mitohondrije b) lizozome c) hloroplaste d) ribozome
12. Organizmi čije ćelije nemaju formirano jezgro, mitohondrije,
Golgijev kompleks, pripadaju grupi
a) prokarioti b) eukarioti c) autotrofi d) heterotrofi
13. Prokarioti uključuju
a) alge b) bakterije c) gljive d) virusi
14. Jedro igra važnu ulogu u ćeliji, jer učestvuje u sintezi
a) glukoza b) lipidi c) vlakna d) nukleinske kiseline i proteini
15. Organoid, odvojen od citoplazme jednom membranom, koji sadrži
mnogi enzimi koji razgrađuju složene organske tvari
za jednostavne monomere, jeste
a) mitohondrija b) ribosoma c) Golgijevog kompleksa d) lizozoma
Pomozite pliz A1. Spajanje filamenata vretena se dešava: u 1) interfazi 2) profazi 3) metefazi 4) anafazi. A2. U profazi ne dolazi do mitozedit: 1) rastvaranje nuklearnog omotača 2) formiranje fisijskog vretena 3) udvostručenje DNK 4) rastvaranje jezgrica. A3) kod životinja tokom mitoze, za razliku od mejoze, nastaju ćelije: 1) somatske 2) sa polovičnim setom hromozoma 3) polne 4) ćelije spora. A4) do divergencije hromotida na polove ćelije dolazi u: 1) profazi prve deobe mejoze 2) profazi druge deobe mejoze 3) interfazi pre prve deobe 4) interfazi pre druge deobe
1. Škrobakumulira se u
A
- hloroplasti B - jezgro C - leukoplasti D - hromoplasti
2. Citoplazma ne ispunjava
funkcija
A
- kretanje supstanci B - interakcije svih organela
V
- napajanje G - zaštitno
3. Rezervni
hranljive materije i otpadni proizvodi se akumuliraju u biljnim ćelijama u
A
- lizozomi B - hloroplasti C - vakuole D - jezgro
4. Proteini,
masti i ugljikohidrati se oksidiraju uz oslobađanje energije u
A
- mitohondrije B - leukoplasti
V
- endoplazmatski retikulum G - Golgijev kompleks
5. "Sastavljanje"
ribosom se javlja u
A
- endoplazmatski retikulum B - Golgijev kompleks
V
- citoplazma G - jezgre
6. Na površini glatkog endoplazmatskog retikuluma sintetišu se molekuli A - mineralne soli B - nukleotidi C - ugljeni hidrati, lipidi G - proteini
7. Na površini grubog endoplazmatskog retikuluma A - lizozomi B - mikrotubule C - mitohondrije D - ribozomi
8. Eukarioti su organizmi koji imaju A - plastide B - bičeve C - ćelijsku membranu D - formirano jezgro
9. Ćelija je osnovna jedinica strukture svih organizama, budući da A - reprodukcija organizama se zasniva na diobi ćelije B - metaboličke reakcije se odvijaju u ćeliji C - ćelijska dioba je u osnovi rasta organizma D - svi organizmi sastoje se od ćelija
10. A - citoplazma B - ćelijski centar C - endoplazmatski retikulum D - vakuola je uključena u formiranje fisijskog vretena
I podjela ćelija. Tipično vreteno je bipolarno - vretenasti sistem mikrotubula formira se između dva pola. Mikrotubule vretena se vežu za kinetohore hromatida u regionu centromera i obezbeđuju kretanje hromozoma prema polovima.
Vreteno se sastoji od tri glavna strukturna elementa: mikrotubula, polova podjele i hromozoma. Centrosomi koji sadrže centriole uključeni su u organiziranje polova diobe kod životinja. U biljkama, kao i u oocitima nekih životinja, centrosomi su odsutni, a formira se acentrosomalno vreteno sa širokim polovima. Motorni proteini koji pripadaju familiji dineina i kinezina igraju važnu ulogu u formiranju vretena.
Potpuno fisiono vreteno formira se u fazi prometafaze nakon uništenja nuklearne membrane, kada citoplazmatske mikrotubule i centrosomi (kod životinja) dobiju pristup hromozomima i drugim komponentama vretena. Izuzetak je vreteno diobe pupajućeg kvasca, koje se formira unutar jezgre.
Struktura
Vreteno diobe tipične ćelije sisara sastoji se od tri strukturni elementi- centrosomi, mikrotubule i hromozomi - koji formiraju simetričnu bipolarnu strukturu. Na polovima vretena nalaze se centrosomi - male organele koje funkcioniraju kao centri organizacije mikrotubula. Svaki centrosom se sastoji od para centriola okruženih s mnogo različitih proteina. Zgusnuti hromozomi se nalaze između polova vretena, koji se sastoje od para hromatida pričvršćenih na centromeri. Na centromernim regijama hromozoma nalaze se kinetohori - složene strukture odgovorne za pričvršćivanje vretena na mikrotubule.
Fisijsko vreteno se sastoji od dva poluvretena. Poluvreteno je formirano od polarizovanih mikrotubula. Negativni minus krajevi mikrotubula se skupljaju na polovima vretena oko centrosoma. Plus krajevi mikrotubula odmiču se od dva pola i ukrštaju se u srednjem ekvatorijalnom dijelu vretena. Kod većine kralježnjaka, poluvreteno se sastoji od 600-750 mikrotubula, od kojih 30-40% završava kinetohorima. Zovu se mikrotubule koje povezuju polove vretena sa kinetohorima hromozoma kinetohoričan... Štaviše, svaki kinetohor se tokom formiranja vretena vezuje za mnoštvo mikrotubula i formira kinetohorni snop. Zovu se mikrotubule koje se nalaze između polova i ne pričvršćuju se za kinetohore interpol... Neke od mikrotubula vretena formiraju radijalne strukture oko svakog pola, koje se nazivaju zvijezde ili asteri. Ove mikrotubule se nazivaju astralno .
U biljkama, kao i u oocitima nekih životinja, centrosomi su odsutni, a formira se acentrosomalno vreteno sa širokim polovima. Također, astralne mikrotubule su odsutne na polovima acentrosomalnog vretena. Inače, struktura vretena biljne ćelije odgovara strukturi vretena životinjske ćelije.
Sklop razdjelnog vretena
Početak montaže vretena u profazi
Sastavljanje fisijskog vretena počinje u profazi. Međutim, u ovoj fazi, formiranje punopravnog vretena je nemoguće zbog izolacije hromozoma, kao i važnih motornih, regulatornih i stabilizirajućih proteina unutar jezgre.
U biljkama, zbog odsustva centrosoma, nuklearni omotač igra ulogu centra organizacije mikrotubula u profazi. Mikrotubule se skupljaju blizu površine jezgra i do kraja profaze su orijentisane duž ose budućeg fisionog vretena, formirajući takozvano profazno vreteno.
U životinjskim ćelijama, centrosom je centar organizacije mikrotubula. Stoga, formiranje fisijskog vretena počinje odvajanjem i divergencijom para centrosoma tokom profaze. Divergenciju centrosoma u profazi osiguravaju motorni proteini dineini. Fiksirani su sa unutrašnje strane stanične membrane i na vanjskoj površini jezgra. Dineini fiksirani u membrani vezuju se za astralne mikrotubule i kreću se prema minus-kraju mikrotubula. Zbog toga se centrosomi pomiču na suprotne dijelove ćelijske membrane i razilaze se dalje jedan od drugog.
Montaža vretena u prometafazi
Samoorganizacija vretena:
Izuzetak je vreteno diobe pupajućeg kvasca, koje se formira unutar jezgre.
Samoorganizacija vretena
Kod svih eukariota, sklapanje bipolarnog vretena u velikoj mjeri ovisi o sposobnosti komponenti vretena da se samoorganiziraju. Samoorganizacija je jedini mehanizam za sklapanje diobenog vretena u stanicama bez centrosoma. Sklop bipolarnog vretena bez uključivanja centrosoma naziva se acentrosomalnim. Karakteristična je za više biljke, a zapaža se i u mejozi u ranim fazama razvoja nekih životinja. Štaviše, pretpostavlja se da je samoorganizacija mikrotubula preovlađujući mehanizam sklapanja vretena, čak i u životinjskim ćelijama koje sadrže centrosome.
Samoorganizacija vretena počinje nakon uništenja nuklearne membrane. Citoplazmatske mikrotubule se skupljaju (nukleiraju) oko hromozoma. Ovdje se uz učešće lokalnih stabilizirajućih faktora produžavaju akumulirajuće mikrotubule. Zatim, organizacija mikrotubula počinje uz učešće tri grupe motornih proteina:
- Motorni proteini porodice kinezin-5(Eg5) vezuju se za dve suprotno orijentisane mikrotubule i istovremeno se kreću prema plus-kraju svake od njih. Kao rezultat toga, antiparalelno polarizirane mikrotubule se sortiraju i "prošive" u području plus-kraja.
- Hromokinezini - proteinski motori porodice kinezina-4 i -10, lokalizovani na kracima hromozoma, vezuju mikrotubule koje se nalaze u blizini hromozoma i kreću se prema plus-kraju mikrotubula. Dakle, krak hromozoma je povezan sa plus-krajem mikrotubula, a minus-kraj je udaljen od hromozoma.
- Treća grupa motornih proteina putuje prema minus krajevima mikrotubula i osigurava snop minus krajeva na polovima vretena. Ova grupa motora uključuje citoplazmatske dineine, kinezin-14. Dynein sudjeluje u fokusiranju fisijskih polova zajedno s brojnim nuklearnim proteinima, na primjer NuMA1(eng. Nu jasno M ikrotubul- A povezani protein 1).
Skupština uz učešće centrosoma
U mnogim životinjskim ćelijama, uključujući ljudske ćelije, centrosomi su uključeni u sastavljanje vretena, koji su polovi vretena podjele. Kao i u sklapanju acentrosomalnog vretena, motorni i drugi proteini su uključeni u samoorganizaciju mikrotubula u bipolarnu strukturu, koja je fokusirana minus krajevima mikrotubula u području centrosoma. U ovom slučaju, centrosomi također sudjeluju u sklapanju vretena i doprinose formiranju polova diobe, ali nisu sastavni dio vretena, budući da se proces sklapanja može odvijati i kada su centrosomi inaktivirani.
Ovisno o vremenu divergencije centrosoma u odnosu na trenutak uništenja nuklearnog omotača, razlikuju se dva mehanizma formiranja vretena:
- Ako se nuklearni omotač uništi prije nego što se centrosom počne divergirati, tada se oslobođeni kromosomi raspoređuju po citoplazmi i formira se "unipolarno" vreteno s mikrotubulama koje se divergiraju od uparenog centrosoma. Dalje formiranje bipolarnog vretena nastaje zbog odbojnih sila mikrotubula koji se preklapaju i pod djelovanjem sila povlačenja astralnih mikrotubula. Odbojnu silu između mikrotubula koji se preklapaju stvaraju proteini slični kinezinu Eg5. Sile povlačenja koje se primjenjuju na astralne mikrotubule stvaraju citoplazmatski dineini vezani za unutrašnju površinu ćelijske membrane.
- Druga opcija uključuje divergenciju centrosoma i formiranje primarnog vretena prije uništenja nuklearne ovojnice. Primarno vreteno nastaje zahvaljujući vučnim silama astralnih mikrotubula, koje stvaraju citoplazmatski dineini, fiksirani na unutrašnjoj površini ćelijske membrane i na površini nuklearne ovojnice. Smjer divergencije centrosoma određuju aktinski filamenti, koji stupaju u interakciju s miozinom koji se nalazi u samim centrosomima ili duž mikrotubula. Primarno vreteno je nestabilno. Za njegovu stabilnost neophodna je interakcija sa kinetohorima hromozoma i drugim proteinima koji se nalaze unutar ćelijskog jezgra.
Vezanje hromozoma za vreteno
Najviše proučavan mehanizam vezivanja hromozoma za vreteno u životinjskim ćelijama koje sadrže centrosome. Tokom profaze, oko centrosoma se formira zvezdasta struktura mikrotubula koji se divergiraju u radijalnom pravcu. Nakon uništenja nuklearne membrane, područje jezgre se aktivno ispituje dinamički nestabilnim mikrotubulama koje su zarobljene kinetohorima hromozoma. Neki hromozomi se brzo vežu za mikrotubule sa suprotnih polova. Drugi dio hromozoma se prvo veže za mikrotubule koje izlaze iz jednog od polova. Zatim se kreće u pravcu odgovarajućeg pola. Zatim, hromozomi povezani sa jednim polom hvataju mikrotubule sa suprotnog pola. U procesu metafaze, oko 10-40 mikrotubula je vezano za svaki kinetohor, koji formira kinetohorni snop. Svi hromozomi su povezani sa suprotnim polovima podjele i sastavljeni su u metafaznu ploču u središtu vretena.
Postoji i alternativni model za vezivanje kinetohora na vreteno, pogodan i za ćelije sa centrosomima i za ćelije bez centrosoma. Prema ovom modelu, nukleacija kratkih mikrotubula se dešava u blizini hromozoma uz učešće kompleksa gama-tubulinskog prstena. Sa svojim plus krajevima, mikrotubule su ugrađene u kinetohore. Nakon toga slijedi kontrolirani rast (polimerizacija) mikrotubula. Produženi minus krajevi mikrotubula su "prošiveni" i fokusirani u području polova podjela uz sudjelovanje motornih proteina. Centrosomi (ako su prisutni) olakšavaju vezivanje kinetohornih mikrotubula na polove podjele.
Bipolarna orijentacija sestrinskih hromatida
Za jednaku distribuciju hromozoma između ćelija kćeri, važno je da kinetohori uparenih hromatida budu vezani za mikrotubule koje izlaze iz suprotnih polova. Normalno bipolarno vezivanje kinetohora za suprotne polove naziva se amphithelic... Međutim, tokom sastavljanja vretena može doći do drugih vezanja hromozoma. Vezanje jedne kinetohore za jedan pol podjele naziva se monotelna... Spajanje dva kinetohora jednog hromozoma odjednom na jedan pol podjele naziva se sintetički... Također je moguće merotelic priključak, u kojem je jedan kinetohor spojen na dva pola odjednom.
Neusklađeno vezivanje je delimično sprečeno samom geometrijom sestrinskih kinetohora, koje se nalaze na suprotnim stranama centromernog regiona hromozoma. Osim toga, nepravilni pričvršćivanje je nestabilno i reverzibilno, a normalno bipolarno vezivanje kinetohora je stabilno. Stabilna veza se postiže zahvaljujući vlačnim silama koje proizlaze iz suprotnih polova pregrade. Glavna komponenta regulatornog sistema odgovorna za pravilno vezivanje kinetohora za suprotne polove je protein kinaza ISBN 978-0-9539181-2-6.
Fazu G1 karakteriše nastavak intenzivnih procesa biosinteze, koji se naglo usporavaju tokom mitoze, a za kratko vreme citokineza potpuno prestaje. Ukupni sadržaj proteina se kontinuirano povećava tokom ove faze. Za većinu ćelija postoji kritična tačka u G1 fazi, takozvana restrikciona tačka. Tokom njenog prolaska dolazi do unutrašnjih promjena u ćeliji, nakon čega ćelija mora proći sve naredne faze ćelijskog ciklusa. Granica između S i G2 faze određena je pojavom supstance - aktivatora S-faze.
G2 faza se smatra periodom pripreme ćelije za početak mitoze. Njegovo trajanje je kraće od ostalih perioda. U njemu se sintetiziraju fisioni proteini (tubulin) i uočava se fosforilacija proteina uključenih u kondenzaciju hromatina.
Profaza
Tokom profaze odvijaju se dva paralelna procesa. To je postupna kondenzacija kromatina, pojava jasno vidljivih hromozoma i raspadanje jezgre, kao i formiranje diobenog vretena, koje osigurava ispravnu raspodjelu kromosoma između stanica kćeri. Ova dva procesa prostorno su razdvojena nuklearnim omotačem, koji opstaje tokom cijele profaze i uništava se tek na svom kraju. Centar organizacije mikrotubula kod većine životinja i nekih biljnih ćelija je ćelijski centar ili centrosom. U interfaznoj ćeliji, nalazi se na strani jezgra. U središnjem dijelu centrosoma nalaze se dvije centriole uronjene u njegov materijal pod pravim kutom jedna prema drugoj. Od perifernog dijela centrosoma odlaze brojni tubuli formirani od proteina tubulina. Oni takođe postoje u interfaznoj ćeliji, formirajući u njoj citoskelet. Mikrotubule se vrlo brzo sklapaju i rastavljaju. Oni su nestabilni i njihov niz se stalno ažurira. Na primjer, u ćelijama fibroblasta u kulturi in vitro, prosječni životni vijek mikrotubula je manji od 10 minuta. Na početku mitoze citoplazmatske mikrotubule se raspadaju, a zatim počinje njihova obnova. Prvo se pojavljuju u perinuklearnoj zoni, formirajući blistavu strukturu - zvijezdu. Središte njegovog formiranja je centrosom. Mikrotubule su polarne strukture jer su molekule tubulina od kojih su formirane orijentirane na određeni način. Jedan njegov kraj se produžava tri puta brže od ostalih. Brzo rastući krajevi se nazivaju plus krajevi, sporo rastući minus krajevi. Osim toga, krajevi su usmjereni naprijed u smjeru rasta. Centriol je mala cilindrična organela debljine oko 0,2 mikrona i dužine 0,4 mikrona. Njegov zid čini devet grupa cevastih trojki. Kod tripleta, jedan tubul je potpun, a dva susjedna su nekompletna. Svaka trojka je nagnuta prema centralnoj osi. Susedni trojci su međusobno povezani umrežavanjem. Novi centrioli nastaju samo udvostručavanjem postojećih. Ovaj proces se poklapa sa vremenom sinteze DNK u S-fazi. U periodu G1, centriole, formirajući par, pomiču se za nekoliko mikrona. Zatim se na svakom od centriola, u njegovom srednjem dijelu, pod pravim uglom ugrađuje po jedan centriol. Rast ćerki centriola je završen u G2 fazi, ali su one i dalje uronjene u jednu masu centrosomalnog materijala. Na početku profaze svaki par centriola postaje dio posebnog centrosoma iz kojeg polazi radijalni snop mikrotubula - zvijezda. Formirane zvijezde se udaljavaju jedna od druge s obje strane jezgra, postajući potom polovi fisionog vretena.
Metafaza

Prometafaza počinje brzim raspadanjem nuklearnog omotača na membranske fragmente koji se ne razlikuju od EPS fragmenata. Kreću se na periferiju ćelije pomoću hromozoma i vretena diobe. Na centromerama hromozoma formira se proteinski kompleks koji na elektronskim fotografijama izgleda kao lamelarna troslojna struktura - kinetohor. Obje hromatide nose jednu kinetohoru; na nju su vezane proteinske mikrotubule fisijskog vretena. Metode molekularne genetike otkrile su da je informacija koja određuje specifičnu konstrukciju kinetohora sadržana u nukleotidnoj sekvenci DNK u regiji centromera. Mikrotubule vretena pričvršćene za kinetohore hromozoma igraju veoma važnu ulogu; prvo, orijentišu svaki hromozom u odnosu na vreteno deobe tako da su njegova dva kinetohora okrenuta suprotnim polovima ćelije. Drugo, mikrotubule pokreću hromozome tako da su njihove centromere u ravnini ćelijskog ekvatora. Ovaj proces u ćelijama sisara traje 10 do 20 minuta i završava se do kraja prometafaze. Broj mikrotubula povezanih sa svakom kinetohorom razlikuje se od vrste do vrste. Kod ljudi ih je od 20 do 40, kod kvasca - 1. Plus krajevi mikrotubula se vezuju za hromozome. Osim kinetohornih mikrotubula, fisiono vreteno također sadrži mikrotubule polova koje se protežu od suprotnih polova i prošivene su posebnim proteinima na ekvatoru. Mikrotubule koje se protežu od centrosoma i nisu uključene u fisijsko vreteno nazivaju se astralne, one formiraju zvijezdu.
Metafaza. Zauzima značajan dio mitoze. Lako se prepoznaje po dvije karakteristike: bipolarnoj strukturi fisijskog vretena i metafaznoj hromozomskoj ploči. Ovo je relativno stabilno stanje ćelije; mnoge ćelije mogu ostati u metafazi nekoliko sati ili dana ako se tretiraju supstancama koje depolimerizuju tubule vretena. Nakon uklanjanja agensa, mitotičko vreteno je sposobno za regeneraciju i ćelija je sposobna da završi mitozu.
Anafaza

Anafaza počinje brzim sinhronim cijepanjem svih hromozoma na sestrinske hromatide, od kojih svaka ima svoj kinetohor. Cepanje hromozoma na hromatide povezano je sa replikacijom DNK u regionu centromera. Replikacija tako male površine događa se za nekoliko sekundi. Signal za početak anafaze dolazi iz citosola; povezan je s kratkotrajnim brzim povećanjem koncentracije kalcijevih jona za faktor 10. Elektronska mikroskopija je pokazala da se membranske vezikule bogate kalcijumom nakupljaju na polovima vretena. Kao odgovor na signal anafaze, sestrinske hromatide počinju da se kreću prema polovima. To je najprije zbog skraćivanja kinetohornih cijevi (anafaza A), a zatim i širenja samih polova, što je povezano sa produžavanjem polarnih mikrotubula (anafaza B). Procesi su relativno samostalni, na šta ukazuje njihova različita osjetljivost na otrove. U različitim organizmima, doprinos anafaze A i anafaze B konačnom razdvajanju hromozoma je različit. Na primjer, u stanicama sisara, anafaza B počinje nakon anafaze A i završava se kada vreteno dosegne dužinu 1,5-2 puta duže nego u metafazi. Kod protozoa prevladava anafaza B, zbog čega se vreteno izdužuje 15 puta. Skraćivanje kinetohornih tubula nastaje njihovom depolimerizacijom. Podjedinice se gube sa plus kraja, tj. sa strane kinetohora, kao rezultat, kinetohor se kreće sa hromozomom do pola. Što se tiče mikrotubula polova. Zatim, u anafazi, oni se sklapaju i produžavaju kako se polovi razilaze. Do kraja anafaze, hromozomi su potpuno podijeljeni u dvije identične grupe na polovima ćelije.
Podjela jezgra i citoplazme su povezani. U ovom slučaju, mitotičko vreteno igra važnu ulogu. U životinjskim ćelijama fisiona brazda se pojavljuje već u anafazi u ravnini ekvatora vretena. Položen je pod pravim uglom u odnosu na dugu osu mitotičkog vretena. Formiranje žlijeba je posljedica aktivnosti kontraktilnog prstena, koji se nalazi ispod stanične membrane. Sastoji se od najfinijih filamenata - aktinskih filamenata. Kontraktilni prsten ima dovoljnu snagu da savije tanku staklenu iglu ubačenu u kavez. Kako se brazda produbljuje, debljina kontraktilnog prstena se ne povećava, jer se dio filamenata gubi sa smanjenjem njegovog radijusa. Nakon završetka citokineze, kontraktilni prsten se potpuno raspada, plazma membrana u području diobene brazde se skuplja. Neko vrijeme se u kontaktnoj zoni novonastalih ćelija zadržava tijelo ostataka tijesno zbijenih mikrotubula. U biljnim ćelijama sa krutim ćelijskim zidom, citoplazma se deli formiranjem novog zida na granici između ćelija kćeri. U biljnim ćelijama nema kontraktibilnog prstena. U ravni ekvatora ćelije formira se fragmoplast, koji se postepeno širi od centra ćelije ka njenoj periferiji, sve dok rastuća ćelijska ploča ne dođe do plazma membrane matične ćelije. Membrane se spajaju, potpuno razdvajajući formirane ćelije.
/. Struktura vretena
2. Funkcije vretena. Mehanizmi kretanja niti
1. Prilikom fisije jezgra između dva suprotna pola ćelije, tzv vreteno, koji se sastoji od:
Od filamenata (vlakana), koji su snopovi velikog broja mikrotubula (ponekad i više od 100);
Dva centriola, od kojih je svaki na svom polu s različitim organizacioni centri:
Ili sa pericentriolarnim centrom organizatora mikrotubula (kod životinja);
Ili sa amorfnim ("polarna kapa" u većini biljaka);
Bilo sa lamelarnim ili slojevitim ("polarna tijela vretena" kod mnogih gljiva i nekih podrasta).
Dodatni organizacioni centar, kinetohor, leži u centromeri svake hromatide. Razlikovati sljedeće vrste navoja vretena:
Hromozomski (kinetohor, ili potezni) filamenti koji se formiraju iz kinetohora i povezuju ga s jednim od polova;
Centralni filamenti formirani od polarnih organizacionih centara i povezuju oba pola;
Polarni filamenti, koji se formiraju samo u prisustvu centriola u pericentriolarnim organizacionim centrima i završavaju u citoplazmi.
2. Funkcije fisiono vreteno su kako slijedi:
Vreteno osigurava divergenciju hromatida ili hromozoma do polova. Kromosomski filamenti se skraćuju i povlače hromozome prema polovima;
Kod životinja se središnji filamenti obično izdužuju i razmiču polove. U tom slučaju se debljina navoja vretena ne mijenja.
Mehanizmipokreti niti:
Aktivno klizanje filamenata vretena nastaje, očigledno, pri interakciji sa proteinom sličnim dineinu. Mehanizam je sličan onom kod flagela;
Aktivnu ulogu igraju mikrofilamenti, koji se pričvršćuju na niti vretena i uz njihovu pomoć izvlače hromate ili kromosome. U aparatu vretena pronađeni su aktinski filamenti i miozin. Citohanazin, koji destabilizira aktinske filamente, može blokirati djelovanje vretena.
Fisijsko vreteno EMBRIOLOGIJA ŽIVOTINJA
SPINNO OD PODJELA, MITOTIČKA BRZINA - sistem mikrotubula u ćeliji koja se dijeli, koji osigurava promjenu staničnih parametara i divergenciju hromozoma u mitozi i mejozi. Formiranje fisijskog vretena završava u metafazi, u telofazi se raspada. Fisijsko vreteno je napravljeno od filamenata mikrotubula. Postoje tri vrste filamenata: polni - formiraju citoskelet polova ćelije, kontinuirani - povezuju polove ćelije, formirajući citoskelet koji vuče ćeliju tokom deobe, i diskontinualni filamenti koji povezuju centromere hromozoma sa polovima ćelija.
Opća embriologija: Terminološki rječnik - Stavropolj. O.V. Dilekova, T.I. Lapina. 2010 .
Pogledajte šta je "podeljeno vreteno" u drugim rečnicima:
PODELA VRETENA- SPREADY DIVISION, štapićasti sistem mikrotubula u citoplazmi ćelije u procesu MITOZE ili MEJOZE. HROMOSOMI su pričvršćeni za ispupčenje fisionog vretena (ekvator). Vreteno diobe uzrokuje razdvajanje hromozoma, uzrokujući podjelu stanica. cm … Naučno-tehnički enciklopedijski rečnik
fisiono vreteno- (biol.), sistem mikrotubula u ćeliji koja se dijeli, koji osigurava divergenciju i striktno identičnu (tokom mitoze) distribuciju hromozoma između ćelija kćeri. * * * Fisija vretena Fisija vretena, u biologiji, sistem mikrotubula u fisijskom ... ... enciklopedijski rječnik
vreteno [fisija]- ahromatsko vreteno Sistem mikrotubula u ćeliji koja se deli, koji obezbeđuje divergenciju hromozoma, postoji u periodu od metafaze do telofaze; mikrotubule sa svojstvima dvostrukog prelamanja čine snopove vidljive u ... ... Vodič za tehnički prevodilac
PODELA VRETENA- ahromatinsko vreteno, sistem mikrotubula u ćeliji koja se deli, koji obezbeđuje divergenciju hromozoma u mitozi i mejozi. V. d. Nastaje u prometafazi i raspada se u telofazi. V. niti, koje su snopovi mikrotubula, posjeduju ... ... Biološki enciklopedijski rječnik
PODELA VRETENA- u biologiji, sistem mikrotubula u ćeliji koja se dijeli, koji osigurava divergenciju i striktno identičnu (tokom mitoze) raspodjelu hromozoma između ćelija kćeri... Veliki enciklopedijski rječnik
Fisijsko vreteno- U ovom članku nedostaju linkovi na izvore informacija. Informacija mora biti provjerljiva, inače se može dovesti u pitanje i izbrisati. Možete... Wikipedia
vreteno [fisija]- vreteno vretena [fisija], akromatsko vreteno. Sistem mikrotubula u ćeliji koja se dijeli, koji osigurava divergenciju hromozoma, postoji u periodu od metafaze do telofaze; mikrotubule sa svojstvima dvostrukog prelamanja čine ... ... Molekularna biologija i genetika. Eksplanatorni rječnik.
fisiono vreteno- (fusus divisionis) ćelijska struktura koja osigurava ravnomjerno razdvajanje hromozoma tokom mitoze ili mejoze; V. d. Nastaje u profazi i sastoji se od centralnih filamenata koji povezuju oba pola ćelije i hromozomskih filamenata koji povezuju polove sa ... Sveobuhvatni medicinski rječnik
PODELA VRETENA- (biol.), sistem mikrotubula u ćeliji koja se dijeli, koji osigurava divergenciju i striktno identičnu (tokom mitoze) raspodjelu hromozoma između ćelija kćeri... Prirodna nauka. enciklopedijski rječnik
PODELA VRETENA- (vreteno) struktura koja se sastoji od mikrotubula i povezanih proteina; formiran tokom mitoze (u profazi) između dva para cntriola (ur.). Mikrotubule se protežu od polova ćelije i sastaju se na ekvatoru, ... ... Eksplanatorni medicinski rječnik