Plasmamembranets funktioner. Plasmamembran Plasmas struktur och funktion
Plasmamembranet intar en speciell position, eftersom det begränsar cellen utanför och är direkt kopplat till den extracellulära miljön. Det är cirka 10 nm tjockt och är det tjockaste cellmembranet. Huvudkomponenterna är proteiner (över 60%), lipider (cirka 40%) och kolhydrater (ca 1%). Liksom alla andra cellmembran syntetiseras det i EPS-kanalerna.
Plasmalemmafunktioner.
Transport.
Plasmamembranet är semipermeabelt, dvs. selektivt passerar olika molekyler genom den med olika hastigheter. Det finns två sätt att transportera ämnen över membranet: passiv och aktiv transport.
Passiv transport. Passiv transport eller diffusion kräver ingen energiinsats. Oladdade molekyler diffunderar längs koncentrationsgradienten, transporten av laddade molekyler beror på koncentrationsgradienten för väteprotoner och den transmembrana potentialskillnaden, som kombineras till en elektrokemisk protongradient. Som regel bär den inre cytoplasmatiska ytan av membranet en negativ laddning, vilket underlättar penetrationen av positivt laddade joner in i cellen. Det finns två typer av diffusion: enkel och lätt.
Enkel diffusion är typisk för små neutrala molekyler (H 2 O, CO 2, O 2), såväl som för hydrofoba organiska ämnen med låg molekylvikt. Dessa molekyler kan passera genom membranets porer eller kanaler utan någon interaktion med membranproteiner så länge som koncentrationsgradienten bibehålls.
Underlättad diffusion är karakteristisk för hydrofila molekyler, som transporteras över membranet även längs koncentrationsgradienten, men med hjälp av speciella membranbärarproteiner enligt principen uniformer.
Underlättad diffusion är mycket selektiv, eftersom bärarproteinet har ett bindningscentrum som är komplementärt till den transporterade substansen, och överföringen åtföljs av konformationsförändringar i proteinet. En av de möjliga mekanismerna för underlättad diffusion är följande: transportproteinet (translokaset) binder ämnet, närmar sig sedan den motsatta sidan av membranet, släpper detta ämne, antar den initiala konformationen och är återigen redo att utföra transportfunktionen. Lite är känt om hur själva proteinet rör sig. En annan möjlig överföringsmekanism involverar involvering av flera bärarproteiner. I detta fall passerar den initialt bundna föreningen själv från ett protein till ett annat, sekventiellt bindande med ett eller annat protein tills det är på motsatt sida av membranet.
Aktiv transport. Sådan transport sker när överföringen utförs mot koncentrationsgradienten. Det kräver att cellen förbrukar energi. Aktiv transport tjänar till ackumulering av ämnen inuti cellen. Energikällan är ofta ATP. För aktiv transport, förutom energikällan, är deltagandet av membranproteiner nödvändigt. Ett av de aktiva transportsystemen i djurceller är ansvarigt för transporten av Na- och K+-joner över cellmembranet. Detta system kallas Na + - K * -pump. Det är ansvarigt för att upprätthålla sammansättningen av den intracellulära miljön, där koncentrationen av K+-joner är högre än för Na*-joner.
Koncentrationsgradienten för båda jonerna upprätthålls genom att överföra K + till insidan av cellen och Na + till utsidan. Båda transporterna sker mot en koncentrationsgradient. Denna fördelning av joner bestämmer vattenhalten i cellerna, nervcellers och muskelcellers excitabilitet och andra egenskaper hos normala celler. Na + -K + -pump är ett protein - transportera ATPas. Molekylen av detta enzym är en oligomer och genomsyrar membranet. Under pumpdriftens hela cykel överförs 3 Na+-joner från cellen till den intercellulära substansen, och i motsatt riktning - 2 K+-joner, medan ATP-molekylens energi används. Det finns transportsystem för överföring av kalciumjoner (Ca 2+ -ATPas), protonpumpar (H + -ATPas), etc.
Den aktiva överföringen av ett ämne genom membranet, som utförs på grund av energin från koncentrationsgradienten av ett annat ämne, kallas symptom... I detta fall har transport-ATPas bindningsställen för båda substanserna. Antiportär ett ämnes rörelse mot dess koncentrationsgradient. I det här fallet rör sig ett annat ämne i motsatt riktning längs sin koncentrationsgradient. Symtom och antiport (samtransport) kan uppstå under absorption av aminosyror från tarmen och reabsorption av glukos från primär urin, med hjälp av energin från koncentrationsgradienten av Na + joner som skapas av Na +, K + -ATPas.
Ytterligare två typer av transporter är endocytos och exocytos.
Endocytos- infångning av stora partiklar av cellen. Det finns flera sätt för endocytos: pinocytos och fagocytos. Vanligtvis under pinocytos förstå infångningen av flytande kolloidala partiklar av en cell, under fagocytos- infångning av blodkroppar (tätare och större partiklar upp till andra celler). Mekanismen för pino- och fagocytos är olika.
I allmänhet kallas inträde i cellen av fasta partiklar eller vätskedroppar från utsidan heterofagi. Denna process är mest utbredd i protozoer, men den är mycket viktig hos människor (liksom hos andra däggdjur). Heterofagi spelar en viktig roll i försvaret av kroppen (segmenterade neutrofiler - granulocyter; makrofagocyter), ombyggnad av benvävnad (osteoklaster), tyroxinbildning av sköldkörtelfolliklar, reabsorption av protein och andra makromolekyler i det proximala nefronet och andra processer.
Pinocytos.
För att externa molekyler ska komma in i cellen måste de först bindas av glykokalyxreceptorer (en uppsättning molekyler bundna till membranets ytproteiner) (Fig.).
På platsen för sådan bindning finns klatrinproteinmolekyler under plasmalemma. Plasmalemmat, tillsammans med de molekyler som är fästa utifrån och klatrinet som ligger bakom cytoplasman, börjar invadera. Utsprånget blir djupare, dess kanter närmar sig och stängs sedan. Som ett resultat delas en bubbla av från plasmalemma, som bär de fångade molekylerna. Clathrin på ytan ser ut som en ojämn kant på elektroniska mikrofotografier, därför kallas sådana bubblor kantade.
Clathrin förhindrar vesiklar från att fästa vid intracellulära membran. Därför kan de kantade vesiklarna fritt transporteras i cellen till just de delar av cytoplasman där deras innehåll ska användas. Det är så i synnerhet steroidhormoner levereras till kärnan. Vanligtvis släpper dock de kantade vesiklarna sin gräns strax efter klyvning från plasmalemma. Clathrin överförs till plasmalemma och kan återigen delta i endocytosreaktioner.
Vid cellytan i cytoplasman finns mer permanenta vesiklar - endosomer. Kantade vesiklar släpper ut klatrin och smälter samman med endosomer, vilket ökar volymen och ytan på endosomerna. Sedan spjälkas den överflödiga delen av endosomerna av i form av en ny vesikel, där det inte finns några ämnen som har kommit in i cellen, de stannar kvar i endosomen. Den nya vesikeln riktas mot cellytan och smälter samman med membranet. Som ett resultat återställs minskningen av plasmalemma, som inträffar under delning av den kantade vesikeln, medan dess receptorer också återgår till plasmalemma.
Endosomer är nedsänkta i cytoplasman och smälts samman med lysosomens membran. De inkommande ämnena inuti en sådan sekundär lysosom genomgår olika biokemiska omvandlingar. När processen har fullbordats kan lysosomens membran sönderdelas till fragment, och sönderfallsprodukterna och innehållet i lysosomen blir tillgängliga för intracellulära metaboliska reaktioner. Till exempel binder aminosyror till tRNA och levereras till ribosomerna, medan glukos kan komma in i Golgi-komplexet eller tubuli i den agranulära EPS.
Även om endosomer inte har en klatrinkant, smälter inte alla samman med lysosomer. Vissa av dem är riktade från en cellyta till en annan (om cellerna bildar ett epitelskikt). Där smälter endosomets membran samman med plasmolemma och innehållet utsöndras. Som ett resultat överförs ämnen genom cellen från en miljö till en annan utan förändringar. Denna process kallas transcytos... Proteinmolekyler, i synnerhet immunglobuliner, kan också transporteras genom transcytos.
Fagocytos.
Om en stor partikel har molekylära grupperingar på sin yta som kan kännas igen av cellreceptorer, binder den. Inte alltid främmande partiklar själva har sådana grupper. Men när de kommer in i kroppen omges de av molekyler av immunglobuliner (opsoniner), som alltid finns i blodet och i den extracellulära miljön. Immunglobuliner känns alltid igen av fagocytiska celler.
Efter att opsoninerna som täcker den främmande partikeln binder till fagocytens receptorer, aktiveras dess ytkomplex. Aktinmikrofilament börjar interagera med myosin, och cellytans konfiguration förändras. Utväxter av fagocytcytoplasman sträcks runt partikeln. De täcker partikelns yta och kombineras över den. De yttre bladen på utväxterna smälter samman och stänger cellytan.
Djupa ark av utväxter bildar ett membran runt den absorberade partikeln - den bildas fagosomen. Fagosomen smälter samman med lysosomer, vilket resulterar i deras komplexa - heterolysosom (heterosom, eller fagolysosom). I den sker lys av de fångade komponenterna i partikeln. Vissa av lysprodukterna avlägsnas från heterosomen och används av cellen, medan vissa kanske inte är mottagliga för verkan av lysosomala enzymer. Dessa rester bildar restkroppar.
Potentiellt alla celler har förmågan till fagocytos, men i kroppen är det bara ett fåtal som specialiserar sig i denna riktning. Dessa är neutrofila leukocyter och makrofager.
Exocytos.
Detta är avlägsnandet av ämnen från cellen. Först segregeras stormolekylära föreningar i Golgi-komplexet i form av transportbubblor. De senare, med deltagande av mikrotubuli, riktas mot cellytan. Vesikelns membran är inbäddat i plasmalemma, och innehållet i vesikeln är utanför cellen (Fig.) Fusionen av vesikeln med plasmalemma kan ske utan några ytterligare signaler. Denna exocytos kallas konstitutiv. Det är så de flesta av produkterna från den egna ämnesomsättningen tas bort från cellen. Ett antal celler är dock utformade för att syntetisera speciella föreningar - hemligheter som används i andra delar av kroppen. För att transportvesikeln med hemligheten ska smälta samman med plasmamembranet krävs externa signaler. Först då kommer fusionen att äga rum och hemligheten kommer att släppas. Denna exocytos kallas reglerad... Signalmolekyler som främjar sekretfrisättning kallas liberiner (frisättande faktorer), och förhindra eliminering - statiner.
Receptorfunktioner.
De tillhandahålls huvudsakligen av glykoproteiner som finns på ytan av plasmalemma och som kan binda till deras ligander. Liganden motsvarar sin receptor som en nyckel till ett lås. Bindning av liganden till receptorn orsakar en förändring i polypeptidens konformation. Med en sådan förändring i transmembranproteinet etableras kommunikation mellan den extra- och intracellulära miljön.
Typer av receptorer.
Receptorer associerade med proteinjonkanaler. De interagerar med en signalmolekyl som tillfälligt öppnar eller stänger kanalen för passage av joner. (T.ex. är receptorn för signalsubstansen acetylkolin ett protein som består av 5 subenheter som bildar en jonkanal. I frånvaro av acetylkolin stängs kanalen, och efter infästning öppnas den och låter natriumjoner passera).
Katalytiska receptorer. De består av en extracellulär del (receptorn i sig) och en intracellulär cytoplasmatisk del, som fungerar som ett enzym prolinkinas (till exempel tillväxthormonreceptorer).
Receptorer associerade med G-proteiner. Dessa är transmembranproteiner som består av en receptor som interagerar med en ligand och ett G-protein (guanosintrifosfatkopplat regulatoriskt protein), som överför en signal till ett membranbundet enzym (adenylatcyklas) eller till en jonkanal. Som ett resultat aktiveras cykliska AMP- eller kalciumjoner. (Så här fungerar adenylatcyklassystemet. I levercellerna finns till exempel en receptor för hormonet insulin. Den supercellulära delen av receptorn binder till insulin. Detta orsakar aktivering av den intracellulära delen - enzymet adenylatcyklas. Det syntetiserar cykliskt AMP från ATP, som reglerar hastigheten för olika intracellulära processer, vilket orsakar aktivering eller hämning av dessa eller andra metaboliska enzymer).
Receptorer som uppfattar fysiska faktorer. Till exempel fotoreceptorproteinet rhodopsin. Vid absorption av ljus ändrar det sin konformation och exciterar en nervimpuls.
Den består av ett bilipidlager, vars lipider är strikt orienterade - den hydrofoba delen av lipiderna (svansen) är vänd inåt lagret, medan den hydrofila delen (huvudet) är utåtriktad. Förutom lipider är tre typer av membranproteiner involverade i konstruktionen av plasmamembranet: perifer, integral och semi-integral.
Ett av studieområdena för membran är för närvarande en detaljerad studie av egenskaperna hos både olika strukturella och regulatoriska lipider och individuella integrala och semi-integrala proteiner som utgör membran.
Integrala membranproteiner
Huvudrollen i själva membranets organisation spelas av integrerade och semi-integrala proteiner, som har en globulär struktur och är associerade med lipidfasen genom hydrofila - hydrofoba interaktioner. Kulorna av integrala proteiner penetrerar hela membranets tjocklek, och deras hydrofoba del är belägen i mitten av kulan och är nedsänkt i den hydrofoba zonen av lipidfasen.
Semi-integrala membranproteiner
I semi-integrala proteiner är hydrofoba aminosyror koncentrerade vid en av kulornas poler, och följaktligen är kulorna endast halvt nedsänkta i membranet och sticker ut utåt från en (extern eller inre) membranyta.
Funktioner hos membranproteiner
Integrala och semi-integrala proteiner i plasmamembranet tilldelades tidigare två funktioner: generella strukturella och specifika. Följaktligen särskiljdes strukturella och funktionella proteiner bland dem. Förbättringen av metoder för att isolera proteinfraktioner av membran och en mer detaljerad analys av enskilda proteiner indikerar nu frånvaron av strukturella proteiner som är universella för alla membran som inte har några specifika funktioner. Däremot är membranproteiner med specifika funktioner mycket olika. Dessa är proteiner som utför receptorfunktioner, proteiner som är aktiva och passiva bärare av olika föreningar, och slutligen proteiner som ingår i många enzymsystem. Material från sajten
Egenskaper hos membranproteiner
Den gemensamma egenskapen för alla dessa integrerade och semi-integrala membranproteiner, som skiljer sig inte bara i funktion, utan också kemiskt, är deras grundläggande förmåga att röra sig, "flyta" i membranplanet i den flytande lipidfasen. Som noterats ovan har förekomsten av sådana rörelser i plasmamembranen hos vissa celler bevisats experimentellt. Men detta är långt ifrån den enda typen av rörelse som identifieras i membranproteiner. Förutom lateral förskjutning kan individuella integrala och semi-integrala proteiner rotera i membranplanet i horisontella och till och med vertikala riktningar, och kan även ändra graden av nedsänkning av molekylen i lipidfasen.
Opsin. Alla dessa olika och komplexa rörelser av proteinkulor illustreras särskilt väl av exemplet med opsinproteinet, som är specifikt för membranen hos fotoreceptorceller (Fig. 3). Som ni vet är opsin i mörker associerad med karotenoiden retinal, som innehåller en dubbel cissbindning; komplexet av retinal och opsin bildar rhodopsin, eller visuellt lila. Rhodopsinmolekylen är kapabel till lateral rörelse och rotation i membranets horisontella plan (Fig. 3, A). När den utsätts för ljus genomgår retinal fotoisomerisering och omvandlas till en transform. I detta fall ändras näthinnans konformation och den separeras från opsinen, vilket i sin tur ändrar rotationsplanet från horisontellt till vertikalt (fig. 3, B). Konsekvensen av sådana transformationer är en förändring i membranpermeabiliteten för joner, vilket leder till uppkomsten av en nervimpuls.
Intressant nog kan förändringar i konformationen av opsinkulor inducerade av ljusenergi inte bara tjäna till att generera en nervimpuls, som sker i cellerna i ögats näthinna, utan är också det enklaste fotosyntetiska systemet som finns i speciella lila bakterier.
Biologiska membran utgör grunden för cellens strukturella organisation. Plasmamembranet (plasmalemma) är membranet som omger cytoplasman i en levande cell. Membran består av lipider och proteiner. Lipider (främst fosfolipider) bildar ett dubbelt lager, där de hydrofoba "svansarna" av molekylerna är riktade mot insidan av membranet, och de hydrofila - mot dess ytor. Proteinmolekyler kan vara lokaliserade på membranets yttre och inre yta, de kan delvis sjunka ner i lipidskiktet eller penetrera det genom och igenom. De flesta av de nedsänkta membranproteinerna är enzymer. Detta är en vätske-ben-mosaikmodell av strukturen hos ett plasmamembran. Protein- och lipidmolekyler är rörliga, vilket säkerställer dynamiken i membranet. Membranen innehåller även kolhydrater i form av glykolipider och glykoproteiner (glykokalyx) placerade på membranets yttre yta. Uppsättningen av proteiner och kolhydrater på membranytan av varje cell är specifik och är en slags indikator på celltypen.
Membranfunktioner:
- Separerande. Det består i bildandet av en barriär mellan cellens inre innehåll och den yttre miljön.
- Säkerställa utbyte av ämnen mellan cytoplasman och den yttre miljön. Vatten, joner, oorganiska och organiska molekyler kommer in i cellen (transportfunktion). Produkterna som bildas i cellen (sekretorisk funktion) utsöndras i den yttre miljön.
- Transport. Transport över membranet kan ske på olika sätt. Passiv transport utförs utan energiförbrukning, genom enkel diffusion, osmos eller underlättad diffusion med hjälp av bärarproteiner. Aktiv transport sker med hjälp av bärarproteiner, och det kräver energi (till exempel en natrium-kaliumpump). Material från sajten
Stora molekyler av biopolymerer kommer in i cellen som ett resultat av endocytos. Det är uppdelat i fagocytos och pinocytos. Fagocytos är infångning och absorption av stora partiklar av cellen. Fenomenet beskrevs först av I.I. Mechnikov. Först fäster ämnena till plasmamembranet, till specifika receptorproteiner, sedan sjunker membranet och bildar en depression.
En matsmältningsvakuol bildas. Det smälter de ämnen som kommer in i cellen. Hos människor och djur är leukocyter kapabla till fagocytos. Vita blodkroppar absorberar bakterier och andra partiklar.
Pinocytos är processen att fånga och absorbera vätskedroppar med ämnen lösta i den. Ämnen fäster vid membranproteiner (receptorer), och en droppe lösning omges av ett membran och bildar en vakuol. Pinocytos och fagocytos inträffar med utgifterna för ATP-energi.
- Sekretorisk. Sekretion - cellens frisättning av ämnen som syntetiseras i cellen till den yttre miljön. Hormoner, polysackarider, proteiner, fettdroppar är inneslutna i vesiklar avgränsade av ett membran, och går till plasmalemma. Membranen smälter samman och innehållet i vesikeln släpps ut i miljön som omger cellen.
- Anslutning av celler i vävnad (på grund av vikta utväxter).
- Receptor. Membran innehåller ett stort antal receptorer - speciella proteiner, vars roll är att överföra signaler från utsidan till insidan av cellen.
1. Barriär- ger en reglerad, selektiv, passiv och aktiv metabolism med omgivningen.
Cellmembran har selektiv permeabilitet: glukos, aminosyror, fettsyror, glycerol och joner diffunderar långsamt genom dem, själva membranen reglerar aktivt denna process - vissa ämnen släpps igenom, medan andra inte gör det.
2. Transport- genom membranet transporteras ämnen in i och ut ur cellen. Transport genom membran ger: leverans av näringsämnen, avlägsnande av slutmetaboliska produkter, utsöndring av olika ämnen, skapande av joniska gradienter, upprätthållande av lämpligt pH och jonkoncentration i cellen, som är nödvändiga för arbetet med cellulära enzymer.
Det finns fyra huvudmekanismer för att ämnen kommer in i cellen eller avlägsnas från cellen till utsidan:
a) Passiv (diffusion, osmos) (kräver ingen energiförbrukning)
Diffusion
Spridningen av molekyler eller atomer av en substans mellan molekyler eller atomer av en annan, vilket leder till en spontan utjämning av deras koncentrationer över hela den upptagna volymen. I vissa situationer har ett av ämnena redan en utjämnad koncentration och de talar om diffusion av ett ämne i ett annat. I detta fall sker överföringen av materia från ett område med hög koncentration till ett område med låg koncentration (längs vektorn för koncentrationsgradienten (fig. 2.4).
Ris. 2.4. Diffusionsprocessdiagram
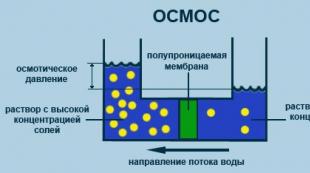
Osmos
Processen med envägsdiffusion genom ett semipermeabelt membran av lösningsmedelsmolekyler mot en högre koncentration av ett löst ämne från en volym med en lägre koncentration av ett löst ämne (fig. 2.5).

Ris. 2.5. Osmos processdiagram
b) Aktiv transport (kräver energiförbrukning)
Natrium-kalium pump- mekanismen för aktiv konjugerad transmembrantransport av natriumjoner (från cellen) och kaliumjoner (in i cellen), vilket ger en koncentrationsgradient och en transmembranpotentialskillnad. Den senare fungerar som grund för många funktioner hos celler och organ: utsöndring av körtelceller, muskelkontraktion, ledning av nervimpulser, etc. (fig. 2.6).

Ris. 2.6. Schemat för kalium-natriumpumpen
I det första steget fäster enzymet Na + / K + -ATPas tre Na +-joner från membranets insida. Dessa joner ändrar konformationen av det aktiva centret av ATPas. Därefter kan enzymet hydrolysera en ATP-molekyl. Energin som frigörs efter hydrolys går åt till att förändra bärarens konformation, vilket gör att tre joner Na + och jon PO 4 3– (fosfat) uppträder på membranets utsida. Här spjälkas Na+-joner och PO 4 3– ersätts med två K+-joner. Därefter återgår enzymet till sin ursprungliga konformation och K+-joner visas på insidan av membranet. Här delas K+-jonerna av, och bäraren är återigen redo för arbete.
Som ett resultat skapas en hög koncentration av Na+-joner i den extracellulära miljön, och en hög koncentration av K+ skapas inuti cellen. Denna koncentrationsskillnad används i celler under ledning av en nervimpuls.
c) Endocytos (fagocytos, pinocytos)
Fagocytos(äter av cellen) - processen för absorption av fasta föremål av cellen, såsom eukaryota celler, bakterier, virus, rester av döda celler, etc. En stor intracellulär vakuol (fagosom) bildas runt det absorberade föremålet. Storleken på fagosomer är från 250 nm och mer. Genom att sammansmälta fagosomen med den primära lysosomen bildas en sekundär lysosom. I en sur miljö bryter hydrolytiska enzymer ner makromolekyler i den sekundära lysosomen. Klyvningsprodukter (aminosyror, monosackarider och andra användbara ämnen) transporteras sedan genom det lysosomala membranet in i cellens cytoplasma. Fagocytos är mycket utbredd. Hos högorganiserade djur och människor spelar fagocytosprocessen en skyddande roll. Den fagocytiska aktiviteten hos leukocyter och makrofager är av stor betydelse för att skydda kroppen från patogena mikrober och andra oönskade partiklar som kommer in i den. Fagocytos beskrevs först av den ryska vetenskapsmannen I.I.Mechnikov (bild 2.7)
Pinocytos(att dricka av cellen) - cellens absorption av vätskefasen från miljön som innehåller lösliga ämnen, inklusive stora molekyler (proteiner, polysackarider, etc.). Vid pinocytos lösgörs små vesiklar - endosomer - från membranet inuti cellen. De är mindre än fagosomer (deras storlek är upp till 150 nm) och innehåller vanligtvis inga stora partiklar. Efter bildandet av endosomen närmar sig den primära lysosomen den, och dessa två membranvesiklar smälter samman. Den resulterande organellen kallas en sekundär lysosom. Processen med pinocytos utförs ständigt av alla eukaryota celler. (fig. 7)
Receptormedierad endocytos - en aktiv specifik process där cellmembranet buktar in i cellen och bildar kantade gropar. Den intracellulära sidan av den fodrade fossan innehåller en uppsättning adaptiva proteiner. Makromolekyler som binder till specifika receptorer på cellytan passerar in i en mycket högre hastighet än ämnen som kommer in i cellerna genom pinocytos.

Ris. 2.7. Endocytos
d) Exocytos (negativ fagocytos och pinocytos)
En cellulär process där intracellulära vesiklar (membranvesiklar) smälter samman med det yttre cellmembranet. Under exocytos frigörs innehållet i de sekretoriska vesiklarna (exocytiska vesiklerna) utanför, och deras membran smälter samman med cellmembranet. Nästan alla makromolekylära föreningar (proteiner, peptidhormoner etc.) frigörs från cellen med denna metod. (bild 2.8)

Ris. 2.8. Exocytosschema
3. Generering och ledning av biopotentialer- med hjälp av membranet upprätthålls en konstant koncentration av joner i cellen: koncentrationen av K+-jonen inuti cellen är mycket högre än utanför, och koncentrationen av Na+ är mycket lägre, vilket är mycket viktigt, eftersom detta säkerställer upprätthållandet av potentialskillnaden på membranet och genereringen av en nervimpuls.
4. Mekanisk- ger cellens autonomi, dess intracellulära strukturer, såväl som förbindelse med andra celler (i vävnader).
5. Energi- under fotosyntes i kloroplaster och cellandning i mitokondrier, fungerar energiöverföringssystem i deras membran, i vilka proteiner också är involverade;
6. Receptor- vissa proteiner i membranet är receptorer (molekyler genom vilka cellen uppfattar vissa signaler).
7. Enzymatisk- membranproteiner är ofta enzymer. Till exempel innehåller plasmamembranen i tarmepitelceller matsmältningsenzymer.
8. Matris- ger ett visst ömsesidigt arrangemang och orientering av membranproteiner, deras optimala interaktion;
9. Burmärkning- det finns antigener på membranet som fungerar som markörer - "etiketter" som gör att du kan identifiera cellen. Dessa är glykoproteiner (det vill säga proteiner med grenade oligosackaridsidokedjor fästa vid dem) som spelar rollen som "antenner". Med hjälp av markörer kan celler känna igen andra celler och agera i samverkan med dem, till exempel under bildandet av organ och vävnader. Det tillåter också immunsystemet att känna igen främmande antigener.
Cellulära inneslutningar
Cellulära inneslutningar inkluderar kolhydrater, fetter och proteiner. Alla dessa ämnen ackumuleras i cellens cytoplasma i form av droppar och korn av olika storlekar och former. De syntetiseras periodiskt i cellen och används i den metaboliska processen.
Cytoplasma
Det är en del av en levande cell (protoplast) utan plasmamembran och kärna. Cytoplasmans sammansättning inkluderar: cytoplasmatisk matris, cytoskelett, organeller och inneslutningar (ibland kallas inneslutningar och innehåll i vakuoler inte som levande materia i cytoplasman). Avgränsad från den yttre miljön av plasmamembranet är cytoplasman cellernas inre halvflytande medium. Kärnan och olika organeller finns i cytoplasman hos eukaryota celler. Den innehåller också en mängd olika inneslutningar - produkter av cellulär aktivitet, vakuoler, såväl som de minsta tubuli och filament som bildar cellens skelett. Proteiner dominerar i sammansättningen av huvudämnet i cytoplasman.
Cytoplasmatiska funktioner
1) de viktigaste metaboliska processerna äger rum i den.
2) förenar kärnan och alla organeller till en helhet, säkerställer deras interaktion.
3) rörlighet, irritabilitet, ämnesomsättning och reproduktion.
Mobilitet finns i olika former:
Intracellulär rörelse av cellcytoplasman.
Amöba-formad rörelse. Denna form av rörelse uttrycks i bildandet av pseudopodi av cytoplasman mot den eller den stimulansen eller bort från den. Denna form av rörelse är inneboende i amöba, blodleukocyter, såväl som vissa vävnadsceller.
Flimrande rörelse. Det manifesterar sig i form av slag av små protoplasmatiska utväxter - flimmerhår och flageller (ciliater, epitelceller från flercelliga djur, spermier, etc.).
Sammandragande rörelse. Det tillhandahålls på grund av närvaron i cytoplasman av en speciell organoid myofibriller, vars förkortning eller förlängning bidrar till sammandragningen och avslappningen av cellen. Förmågan att dra ihop sig är mest utvecklad i muskelceller.
Irritabilitet uttrycks i cellers förmåga att svara på irritation genom att ändra ämnesomsättning och energi.
Cytoskelett
En av de utmärkande egenskaperna hos en eukaryot cell är närvaron i dess cytoplasma av skelettformationer i form av mikrotubuli och buntar av proteinfibrer. Elementen i cytoskelettet, nära förknippade med det yttre cytoplasmatiska membranet och kärnhöljet, bildar komplexa vävar i cytoplasman.
Cytoskelettet bildas av mikrotubuli, mikrofilament och det mikrotrabekulära systemet. Cytoskelettet bestämmer cellens form, deltar i cellrörelser, i själva cellens delning och rörelse, i den intracellulära transporten av organeller.
Mikrotubuli finns i alla eukaryota celler och är ihåliga ogrenade cylindrar, vars diameter inte överstiger 30 nm och väggtjockleken är 5 nm. De kan vara upp till flera mikrometer långa. Lätt sönderfalla och återmontera. Mikrotubuliväggen består huvudsakligen av spiralvikta tubulinproteinsubenheter (bild 2.09)
Mikrotubulus funktioner:
1) utföra en stödfunktion;
2) bilda en klyvningsspindel; säkerställa divergensen av kromosomer till cellens poler; är ansvariga för rörelsen av cellorganeller;
3) delta i intracellulär transport, sekretion, cellväggsbildning;
4) är en strukturell komponent av flimmerhår, flageller, basala kroppar och centrioler.
Mikrofilament representeras av filament med en diameter på 6 nm, bestående av aktinprotein, som ligger nära muskelaktin. Aktin utgör 10-15% av det totala proteinet i cellen. I de flesta djurceller bildas ett tätt nätverk av aktinfilament och associerade proteiner under själva plasmamembranet.
Förutom aktin finns även myosinfilament i cellen. Men deras antal är mycket mindre. På grund av interaktionen mellan aktin och myosin uppstår muskelsammandragning. Mikrofilament är förknippade med rörelsen av hela cellen eller dess individuella strukturer inom den. I vissa fall tillhandahålls rörelse endast av aktinfilament, i andra - av aktin tillsammans med myosin.
Funktioner av mikrofilament
1) mekanisk styrka
2) låter cellen ändra sin form och röra sig.

Ris. 2.09. Cytoskelett
Organeller (eller organeller)
Delas in i icke-membran, enkelmembran och dubbelmembran.
TILL icke-membranorganeller eukaryota celler inkluderar organeller som inte har sitt eget slutna membran, nämligen: ribosomer och organeller byggda på basis av tubulin mikrotubuli - cellcentrum (centrioler) och rörelseorganeller (flageller och flimmerhår). I cellerna hos de flesta encelliga organismer och den överväldigande majoriteten av högre (marklevande) växter saknas centrioler.
TILL enmembranorganeller relatera: endoplasmatiskt retikulum, Golgi-apparater, lysosomer, peroxisomer, sfärosomer, vakuoler och några andra. Alla enmembranorganeller är sammankopplade till ett enda cellsystem. Växtceller har speciella lysosomer, djurceller har speciella vakuoler: matsmältning, utsöndring, kontraktil, fagocytisk, autofagocytisk, etc.
TILL tvåmembransorganeller relatera mitokondrier och plastider.
Icke-membranorganeller
A) Ribosomer- organeller som finns i cellerna i alla organismer. Dessa är små organeller, representerade av klotformade partiklar med en diameter på cirka 20 nm. Ribosomer är sammansatta av två underenheter av olika storlek - stora och små. Ribosomer inkluderar proteiner och ribosomalt RNA (rRNA). Det finns två huvudtyper av ribosomer: eukaryota (80S) och prokaryota (70S).
Beroende på lokaliseringen i cellen finns det fria ribosomer belägna i cytoplasman som syntetiserar proteiner och fästa ribosomer - ribosomer förbundna med stora subenheter med den yttre ytan av EPR-membranen, syntetiserar proteiner som kommer in i Golgi-komplexet, och sedan utsöndras av Cellen. Under proteinbiosyntesen kan ribosomer kombineras till komplex - polyribosomer (polysomer).
Eukaryota ribosomer bildas i kärnan. Först syntetiseras rRNA på det nukleolära DNA:t, som sedan täcks med ribosomala proteiner som kommer från cytoplasman, klyvs till önskad storlek och bildar ribosomsubenheter. Det finns inga färdigbildade ribosomer i kärnan. Föreningen av subenheter till en hel ribosom sker i cytoplasman, vanligtvis under proteinbiosyntes.
Ribosomer finns i cellerna i alla organismer. Var och en består av två partiklar, små och stora. Ribosomerna inkluderar proteiner och RNA.
Funktioner
proteinsyntes.
De syntetiserade proteinerna ackumuleras först i kanalerna och håligheterna i det endoplasmatiska retikulumet och transporteras sedan till organellerna och delar av cellen. EPS och ribosomer placerade på dess membran representerar en enda apparat för biosyntes och transport av proteiner (fig. 2.10-2.11).

Ris. 2.10. Ribosomstruktur

Ris. 2.11. Ribosomstruktur
B) Cellcentrum (centrioler)
Centriolen är en cylinder (0,3 µm lång och 0,1 µm i diameter), vars vägg är bildad av nio grupper av tre sammanslagna mikrotubuli (9 tripletter), sammankopplade med vissa intervall genom tvärbindning. Centrioler är ofta parade, där de är i rät vinkel mot varandra. Om centriolen ligger vid basen av cilium eller flagellum, kallas det basalkroppen.
Nästan alla djurceller har ett par centrioler, som är mittelementet i cellcentret.
Före delning divergerar centriolerna till motsatta poler och en dottercentriol dyker upp nära var och en av dem. Från centrioler som ligger vid olika poler i cellen bildas mikrotubuli som växer mot varandra.
Funktioner
1) bildar en mitotisk spindel, som bidrar till en enhetlig fördelning av genetiskt material mellan dotterceller,
2) är centrum för organiseringen av cytoskelettet. Några av spindelfilamenten är fästa vid kromosomerna.
Centrioler är självreplikerande organeller i cytoplasman. De uppstår som ett resultat av dubblering av befintliga. Detta inträffar när centriolen divergerar. Omogen centriol innehåller 9 enkla mikrotubuli; uppenbarligen är varje mikrotubuli en matris för sammansättning av tripletter som är karakteristiska för en mogen centriol (fig. 2.12).
Cetrioler finns i cellerna hos lägre växter (alger).

Ris. 2.12. Cellcentrumcentrioler
Enkelmembranorganeller
D) Endoplasmatiskt retikulum (EPS)
Hela cytoplasmans inre zon är fylld med många små kanaler och håligheter, vars väggar är membran som liknar plasmamembranets struktur. Dessa kanaler förgrenar sig, ansluter till varandra och bildar ett nätverk som kallas endoplasmatiska retikulum. Det endoplasmatiska retikelet är heterogent till sin struktur. Två typer av det är kända - grynig och slät.
På membranen i kanalerna och kaviteterna i det granulära nätverket finns det många små rundade kroppar - ribosom som ger hinnorna ett strävt utseende. Membranen i det släta endoplasmatiska retikulum bär inte ribosomer på sin yta. EPS utför många olika funktioner.
Funktioner
Huvudfunktionen hos det granulära endoplasmatiska retikulumet är deltagande i proteinsyntes, som utförs i ribosomerna. På membranen i det släta endoplasmatiska retikulum syntetiseras lipider och kolhydrater. Alla dessa syntesprodukter ackumuleras i kanaler och håligheter och transporteras sedan till olika organeller i cellen, där de konsumeras eller ackumuleras i cytoplasman som cellulära inneslutningar. EPS förbinder cellens huvudorganeller (fig. 2.13).

Ris. 2.13. Strukturen av det endoplasmatiska reticulum (EPS) eller reticulum
E) Golgi-apparat
Strukturen hos denna organoid är liknande i cellerna hos växt- och djurorganismer, trots mångfalden av dess form. Den har många viktiga funktioner.
Enkel membran organoid. Det är en stapel av tillplattade "cisterner" med förlängda kanter, med vilka ett system av små enmembranbubblor (Golgi-bubblor) är associerat. Golgi-bubblor är huvudsakligen koncentrerade på sidan som gränsar till EPS och på periferin av staplarna. Man tror att de överför proteiner och lipider till Golgi-apparaten, vars molekyler, som rör sig från cistern till cistern, genomgår kemisk modifiering.
Alla dessa ämnen ackumuleras först, komplicerar kemiskt och kommer sedan i form av stora och små bubblor in i cytoplasman och används antingen i själva cellen under dess liv eller tas bort från den och används i kroppen (fig. 2.14-2.15).

Ris. 2.14. Strukturen av Golgi-apparaten
Funktioner:
Modifiering och ackumulering av proteiner, lipider, kolhydrater;
Packning i membranvesiklar (vesiklar) av inkommande organiska ämnen;
Plats för bildandet av lysosomer;
Sekretorisk funktion, därför är Golgi-apparaten väl utvecklad i sekretoriska celler.

Ris. 2.15. Golgi komplex
E) Lysosomer
De är små rundade kroppar. Inuti lysosomen finns enzymer som bryter ner proteiner, fetter, kolhydrater, nukleinsyror. Lysosomer närmar sig matpartikeln som kom in i cytoplasman, smälter samman med den och en matsmältningsvakuol bildas, inuti vilken det finns en matpartikel omgiven av lysosomala enzymer.
Lysosomala enzymer syntetiseras på en grov EPS, överförs till Golgi-apparaten, där de modifieras och förpackas i membranvesiklar av lysosomer. Lysosomen kan innehålla från 20 till 60 olika typer av hydrolytiska enzymer. Nedbrytning av ämnen med hjälp av enzymer kallas lys.
Skilj mellan primära och sekundära lysosomer. Primära lysosomer kallas som har knoppat från Golgi-apparaten.
Sekundära lysosomer kallas, bildade som ett resultat av fusionen av primära lysosomer med endocytiska vakuoler. I det här fallet smälter de ämnen som kom in i cellen genom fagocytos eller pinocytos, så de kan kallas matsmältningsvakuoler.
Lysosomers funktioner:
1) nedbrytning av ämnen eller partiklar (bakterier, andra celler) som fångas upp av cellen under endocytos,
2) autofagi - förstörelsen av strukturer som är onödiga för cellen, till exempel under ersättning av gamla organeller med nya, eller nedbrytning av proteiner och andra ämnen som produceras inuti själva cellen,
3) autolys - självsmältning av en cell, vilket leder till dess död (ibland är denna process inte patologisk, men följer med kroppens utveckling eller differentieringen av vissa specialiserade celler) (fig. 2.16-2.17).
Exempel: När en grodyngel förvandlas till en groda smälter lysosomerna i svanscellerna den: svansen försvinner, och de ämnen som bildas under denna process absorberas och används av andra celler i kroppen.

Ris. 2.16. Lysosombildning

Ris. 2.17. Lysosomers funktion
G) Peroxisomer
Organoider, liknande i strukturen som lysosomer, vesiklar med en diameter på upp till 1,5 mikron med en homogen matris som innehåller cirka 50 enzymer.
Katalas orsakar nedbrytning av väteperoxid 2H 2 O 2 → 2H 2 O + O 2 och förhindrar lipidperoxidation
Peroxisomer bildas genom knoppning från tidigare existerande, d.v.s. tillhör självreproducerande organeller, trots att de inte innehåller DNA. De växer på grund av att enzymer tränger in i dem, peroxisomenzymer bildas på den grova EPS och i hyaloplasman (bild 2.18).

Ris. 2.18. Peroxisom (i mitten av den kristallina nukleoiden)
H) Vakuoler
Enkelmembranorganeller. Vakuoler är "behållare" fyllda med vattenlösningar av organiska och oorganiska ämnen. EPS och Golgi-apparaten är involverade i bildandet av vakuoler.
Unga växtceller innehåller många små vakuoler, som sedan, när cellerna växer och differentierar, smälter samman med varandra och bildar en stor central vakuol.
Den centrala vakuolen kan uppta upp till 95 % av volymen av en mogen cell, medan kärnan och organellerna trycks tillbaka till cellmembranet. Membranet som begränsar växtvakuolen kallas tonoplast.
Vätskan som fyller växtvakuolen kallas cellsav. Sammansättningen av cellsav inkluderar vattenlösliga organiska och oorganiska salter, monosackarider, disackarider, aminosyror, slutprodukter eller giftiga metaboliska produkter (glykosider, alkaloider), vissa pigment (antocyaniner).
Av organiska ämnen lagras oftare sockerarter och proteiner. Socker - oftare i form av lösningar kommer proteiner i form av EPR-bubblor och Golgi-apparaten, varefter vakuolerna dehydreras och förvandlas till aleuronkorn.
I djurceller finns små matsmältnings- och autofagiska vakuoler som tillhör gruppen sekundära lysosomer och som innehåller hydrolytiska enzymer. Hos encelliga djur finns också kontraktila vakuoler som utför funktionen osmoreglering och utsöndring.
Funktioner
I växter
1) ansamling av vätska och underhåll av turgor,
2) ackumulering av reservnäringsämnen och mineralsalter,
3) färgning av blommor och frukter och därmed attrahera pollinatörer och distributörer av frukter och frön.
Hos djur:
4) matsmältningsvakuoler - förstör organiska makromolekyler;
5) kontraktila vakuoler reglerar cellens osmotiska tryck och tar bort onödiga ämnen från cellen
6) fagocytiska vakuoler bildas under fagocytos av immunceller av antigener
7) autofagocytiska vakuoler bildas under fagocytos av immunceller i sina egna vävnader
Tvåmembransorganeller (mitokondrier och plastider)
Dessa organeller är semi-autonoma, eftersom de har sitt eget DNA och sin egen proteinsyntetiserande apparat. Mitokondrier finns i nästan alla eukaryota celler. Plastider finns bara i växtceller.
I) Mitokondrier
Dessa är organeller för energiförsörjning för metaboliska processer i cellen. I hyaloplasman är mitokondrierna vanligtvis diffust fördelade, men i specialiserade celler är de koncentrerade till de områden där det finns störst energibehov. Till exempel, i muskelceller, är ett stort antal mitokondrier koncentrerade längs de kontraktila fibrillerna, längs spermiernas flagellum, i njurtubuliets epitel, i området för synapser, etc. Detta arrangemang av mitokondrier ger mindre förlust av ATP under dess spridning.
Det yttre membranet separerar mitokondrierna från cytoplasman, är stängt om sig självt och bildar inte invaginationer. Det inre membranet begränsar det inre innehållet i mitokondrierna - matrisen. En karakteristisk egenskap är bildandet av många invaginationer - cristae, på grund av vilka arean av de inre membranen ökar. Antalet och graden av utveckling av cristae beror på vävnadens funktionella aktivitet. Mitokondrier har sitt eget genetiska material (fig. 2.19).
Mitokondriellt DNA är en sluten cirkulär dubbelsträngad molekyl, i mänskliga celler har den en storlek på 16569 nukleotidpar, vilket är ungefär 105 gånger mindre än det DNA som är lokaliserat i kärnan. Mitokondrier har sitt eget proteinsyntetiseringssystem, medan antalet proteiner som översätts från mitokondriellt mRNA är begränsat. Mitokondriellt DNA kan inte koda för alla mitokondrieproteiner. De flesta av proteinerna i mitokondrier är under genetisk kontroll av kärnan.

Ris. 2.19. Mitokondriell struktur
Mitokondriella funktioner
1) bildandet av ATP
2) proteinsyntes
3) deltagande i specifika synteser, till exempel syntesen av steroidhormoner (binjurar)
4) förbrukade mitokondrier kan också ackumulera utsöndringsprodukter, skadliga ämnen, d.v.s. kan ta på sig funktionerna hos andra cellorganeller
K) Plastider
Plastider– Organeller, karakteristiska endast för växter.
Det finns tre typer av plastider:
1) kloroplaster(gröna plastider);
2) kromoplaster(gula, orange eller röda plastider)
3) leukoplaster(färglösa plastider).
Vanligtvis finns bara en typ av plastider i en cell.
Kloroplaster
Dessa organeller finns i cellerna i löv och andra gröna växtorgan, såväl som i en mängd olika alger. Högre växter innehåller vanligtvis flera tiotals kloroplaster i en cell. Den gröna färgen på kloroplaster beror på innehållet av klorofyllpigment i dem.
Kloroplast är den huvudsakliga organoiden av växtceller, i vilken fotosyntes äger rum, det vill säga bildandet av organiska ämnen (kolhydrater) från oorganiskt (CO 2 och H 2 O) med hjälp av solljusets energi. Kloroplaster liknar mitokondrier strukturellt.
Kloroplaster har en komplex struktur. De är avgränsade från hyaloplasman av två membran - externa och interna. Det inre innehållet kallas stroma... Det inre membranet bildar ett komplext, strikt ordnat system av membran i form av platta bubblor, kallade tylakoider.
Tylakoider samlas i högar - korn liknar kolumner av mynt . Grana är förbundna med varandra av stromala tylakoider, som passerar genom dem längs plastiden (fig. 2.20-2.22). Klorofyll och kloroplaster produceras endast genom exponering för ljus.

Ris. 2.20. Kloroplaster under ett ljusmikroskop

Ris. 2.21. Kloroplaststruktur under ett elektronmikroskop

Ris. 2.22. Schematisk struktur av kloroplaster
Funktioner
1) fotosyntes(bildningen av organiska ämnen från oorganiska ämnen på grund av ljusets energi). Klorofyll spelar en central roll i denna process. Den absorberar ljusenergi och styr den att utföra fotosyntetiska reaktioner. I kloroplaster, som i mitokondrier, syntetiseras ATP.
2) delta i syntesen av aminosyror och fettsyror,
3) tjäna som förvar för tillfälliga stärkelsereserver.
Leukoplaster- små färglösa plastider som finns i cellerna i organ dolda från solljus (rötter, rhizomer, knölar, frön). Deras struktur liknar strukturen hos kloroplaster. (fig. 2.23).
Men till skillnad från kloroplaster har leukoplaster ett dåligt utvecklat inre membransystem, eftersom de är involverade i syntesen och ackumuleringen av reservnäringsämnen - stärkelse, proteiner och lipider. I ljuset kan leukoplaster förvandlas till kloroplaster.

Ris. 2.23. Leukoplaststruktur
Kromoplaster- plastider av orange, röd och gul färg, som orsakas av pigment som tillhör gruppen karotenoider. Kromoplaster finns i cellerna i kronbladen hos många växter, mogna frukter, sällan rotgrödor, såväl som i höstlöv. Det inre membransystemet i kromoplaster är som regel frånvarande (fig. 24).

Ris. 2.24. Kromoplaststruktur
Betydelsen av kromoplaster har ännu inte helt klarlagts. De flesta av dem är åldrande plastider. De utvecklas som regel från kloroplaster, medan klorofyll och den inre membranstrukturen förstörs i plastiderna och karotenoider ackumuleras. Detta händer när frukterna mognar och löven gulnar på hösten. Den biologiska betydelsen av kromoplaster ligger i det faktum att de bestämmer den ljusa färgen på blommor och frukter, vilket lockar insekter för korspollinering och andra djur för att sprida frukt. Leukoplaster kan också förvandlas till kromoplaster.
Funktioner av plastider
Syntes av organiska ämnen i klorofyll från enkla oorganiska föreningar: koldioxid och vatten i närvaro av mängder solljus - fotosyntes, ATP-syntes i ljusfasen av fotosyntesen
Syntes av proteiner på ribosomer (mellan kloroplastens inre membran finns DNA, RNA och ribosomer, därför syntetiseras protein i kloroplaster, såväl som i mitokondrier, vilket är nödvändigt för dessa organellers aktivitet).
Närvaron av kromoplaster förklaras av den gula, orange och röda färgen på kronan av blommor, frukter, höstlöv.
Leukoplaster innehåller lagringsämnen (i stjälkar, rötter, knölar).
Kloroplaster, kromoplaster och leukoplaster kan övergå mellan celler. Så när frukter mognar eller ändrar färg på löv på hösten, förvandlas kloroplaster till kromoplaster, och leukoplaster kan förvandlas till kloroplaster, till exempel när potatisknölar blir gröna.
I en evolutionär mening är den primära, initiala typen av plastider kloroplaster, från vilka plastiderna av de andra två typerna härstammar. Plastider delar många likheter med mitokondrier, vilket skiljer dem från andra komponenter i cytoplasman. Detta är först och främst ett skal av två membran och relativ genetisk autonomi, på grund av närvaron av dess egna ribosomer och DNA. Denna egenhet hos organeller utgjorde grunden för idén att föregångarna till plastider och mitokondrier var bakterier, som i evolutionsprocessen visade sig vara inbyggda i en eukaryot cell och gradvis förvandlades till kloroplaster och mitokondrier. (fig. 2.25).

Ris. 2,25. Bildandet av mitokondrier och kloroplaster enligt teorin om symbiogenes
Cell- en elementär enhet i ett levande system. Olika strukturer i en levande cell, som är ansvariga för utförandet av en viss funktion, kallas organeller, liksom organen i hela organismen. Specifika funktioner i cellen är fördelade mellan organeller, intracellulära strukturer som har en viss form, såsom cellkärnan, mitokondrier etc.
Cellulära strukturer:
Cytoplasma... Den obligatoriska delen av cellen, innesluten mellan plasmamembranet och kärnan. CytosolÄr en trögflytande vattenlösning av olika salter och organiska ämnen, genomträngd av ett system av proteinfilament - cytoskelett. De flesta av cellens kemiska och fysiologiska processer äger rum i cytoplasman. Struktur: Cytosol, cytoskelett. Funktioner: inkluderar olika organeller, cellens inre miljö
Plasmamembran... Varje cell av djur, växter, begränsas från miljön eller andra celler av plasmamembranet. Tjockleken på detta membran är så liten (ca 10 nm) att det bara kan ses med ett elektronmikroskop.

Lipider de bildar ett dubbelt lager i membranet, och proteiner genomsyrar hela dess tjocklek, är nedsänkta på olika djup i lipidskiktet eller är belägna på membranets yttre och inre ytor. Strukturen av membranen i alla andra organeller liknar plasmamembranet. Struktur: dubbla lager av lipider, proteiner, kolhydrater. Funktioner: begränsa, bibehålla cellens form, skydda mot skador, reglera intag och avlägsnande av ämnen.
Lysosomer... Lysosomer är membranorganeller. De har en oval form och en diameter på 0,5 mikron. De innehåller en uppsättning enzymer som förstör organiskt material. Lysosommembranet är mycket starkt och förhindrar inträngning av sina egna enzymer i cellens cytoplasma, men om lysosomen skadas av någon yttre påverkan, förstörs hela cellen eller en del av den.
Lysosomer finns i alla celler hos växter, djur och svampar.
Genom att smälta olika organiska partiklar ger lysosomer ytterligare "råmaterial" för kemiska och energimässiga processer i cellen. Under svält smälter lysosomceller några organeller utan att döda cellen. Denna partiella matsmältning ger cellen det nödvändiga minimumet av näringsämnen under en tid. Ibland smälter lysosomer hela celler och grupper av celler, vilket spelar en viktig roll i utvecklingen av djur. Ett exempel är förlusten av en svans när en grodyngel förvandlas till en groda. Struktur: ovala vesiklar, membran utanför, enzymer inuti. Funktioner: dela organiskt material, förstöra döda organeller, förstöra förbrukade celler.
Golgi komplex... Produkterna av biosyntes som kommer in i lumen i håligheterna och tubuli i det endoplasmatiska retikulumet koncentreras och transporteras i Golgi-apparaten. Denna organoid är 5–10 µm stor.

Strukturera: membranomgivna håligheter (vesiklar). Funktioner: ackumulering, packning, utsöndring av organiskt material, bildning av lysosomer
Endoplasmatiska retiklet... Det endoplasmatiska retikulumet är ett system för syntes och transport av organiska ämnen i cellens cytoplasma, som är en genombruten struktur av sammankopplade hålrum.
Ett stort antal ribosomer är fästa vid membranen i det endoplasmatiska retikulum - de minsta organellerna i cellen, som ser ut som en sfär med en diameter på 20 nm. och består av RNA och protein. Proteinsyntes sker på ribosomer. Sedan kommer de nysyntetiserade proteinerna in i systemet av håligheter och tubuli, längs vilka de rör sig inuti cellen. Hålrum, tubuli, tubuli från membran, på ytan av ribosommembranen. Funktioner: syntes av organiska ämnen med hjälp av ribosomer, transport av ämnen.
Ribosomer... Ribosomer är fästa vid membranen i det endoplasmatiska retikulumet eller är fritt belägna i cytoplasman, de är belägna i grupper, proteiner syntetiseras på dem. Proteinsammansättning, ribosomalt RNA Funktioner: ger proteinbiosyntes (sammansättning av en proteinmolekyl från).
Mitokondrier... Mitokondrier är energiorganeller. Formen på mitokondrier är annorlunda, de kan vara resten, stavformade, filamentösa med en genomsnittlig diameter på 1 mikron. och en längd av 7 mikron. Antalet mitokondrier beror på cellens funktionella aktivitet och kan nå tiotusentals i insekters flygande muskler. Mitokondrierna avgränsas på utsidan av det yttre membranet, under finns det inre membranet, som bildar många utväxter - cristae.

Inuti mitokondrierna finns RNA, DNA och ribosomer. Specifika enzymer är inbyggda i dess membran, med hjälp av vilka energin från livsmedelsämnen omvandlas till energin av ATP, som är nödvändig för cellens liv och kroppen som helhet, i mitokondrierna.
Membran, matris, utväxter - cristae. Funktioner: syntes av ATP-molekylen, syntes av sina egna proteiner, nukleinsyror, kolhydrater, lipider, bildandet av sina egna ribosomer.
Plastider... Endast i en växtcell: Lekoplaster, kloroplaster, kromoplaster. Funktioner: ansamling av reservorganiska ämnen, attraktion av pollinerande insekter, syntes av ATP och kolhydrater. Kloroplaster är formade som en skiva eller sfär med en diameter på 4–6 µm. Med dubbelt membran - yttre och inre. Inuti kloroplasten finns ribosom-DNA och speciella membranstrukturer - korn, kopplade till varandra och till kloroplastens inre membran. Varje kloroplast innehåller cirka 50 korn, förskjutna för bättre ljusfångning. Klorofyll finns i granmembranen, tack vare det omvandlas solljusenergin till den kemiska energin av ATP. ATP-energi används i kloroplaster för syntes av organiska föreningar, främst kolhydrater.
Kromoplaster... De röda och gula pigmenten som finns i kromoplaster ger olika delar av växten en röd och gul färg. morötter, tomatfrukter.
Leukoplaster är en plats för ackumulering av ett reservnäringsämne - stärkelse. Det finns särskilt många leukoplaster i cellerna hos potatisknölar. I ljuset kan leukoplaster förvandlas till kloroplaster (som ett resultat av vilket potatiscellerna blir gröna). På hösten förvandlas kloroplaster till kromoplaster och gröna blad och frukter blir gula och röda.
Cellcentrum... Består av två cylindrar, centrioler, placerade vinkelrätt mot varandra. Funktioner: gängstöd för delande spindlar

Cellulära inneslutningar uppträder i cytoplasman och försvinner sedan i celllivsprocessen.

Täta, i form av granulat, innehåller inneslutningar reservnäringsämnen (stärkelse, proteiner, sockerarter, fetter) eller avfallsprodukter från cellen, som ännu inte kan avlägsnas. Alla plastider av växtceller har förmågan att syntetisera och ackumulera reservnäringsämnen. I växtceller sker ansamling av reservnäringsämnen i vakuolerna.
Korn, granulat, droppar Funktioner: icke-permanenta formationer som lagrar organiskt material och energi
Kärna... Kärnmembran av två membran, kärnjuice, nukleolus. Funktioner: lagring av ärftlig information i cellen och dess reproduktion, syntes av RNA - informativ, transport, ribosomal. Det finns sporer i kärnmembranet, genom vilka ett aktivt utbyte av ämnen mellan kärnan och cytoplasman äger rum. Kärnan lagrar ärftlig information inte bara om alla tecken och egenskaper hos en given cell, om de processer som måste inträffa för den (till exempel proteinsyntes), utan också om egenskaperna hos organismen som helhet. Information registreras i DNA-molekyler, som är huvuddelen av kromosomerna. Det finns en kärna i kärnan. Kärnan, på grund av närvaron i den av kromosomer som innehåller ärftlig information, utför funktionerna hos ett centrum som kontrollerar all vital aktivitet och utveckling av cellen.